Voluntary Exercise Positively Affects the Recovery of Long-Nerve Gap Injury Following Tube-Bridging with Human Skeletal Muscle-Derived Stem Cell Transplantation

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Collection of Human Skeletal Muscle Samples
2.2. Isolation of Skeletal Muscle-Derived Stem Cells (Sk-SCs), Cell Sorting, and Expansion
2.3. Recipient Animals, Voluntary Wheel Exercise and Cell Transplantation
2.4. Functional Assessment of Downstream Muscles
2.5. Immunohistochemical Analysis
2.6. Characterization of Downstream Plantar Flexor Muscles and Muscle Spindles, and Associated Tibial Nerves
2.7. Quantitative Analysis
3. Results
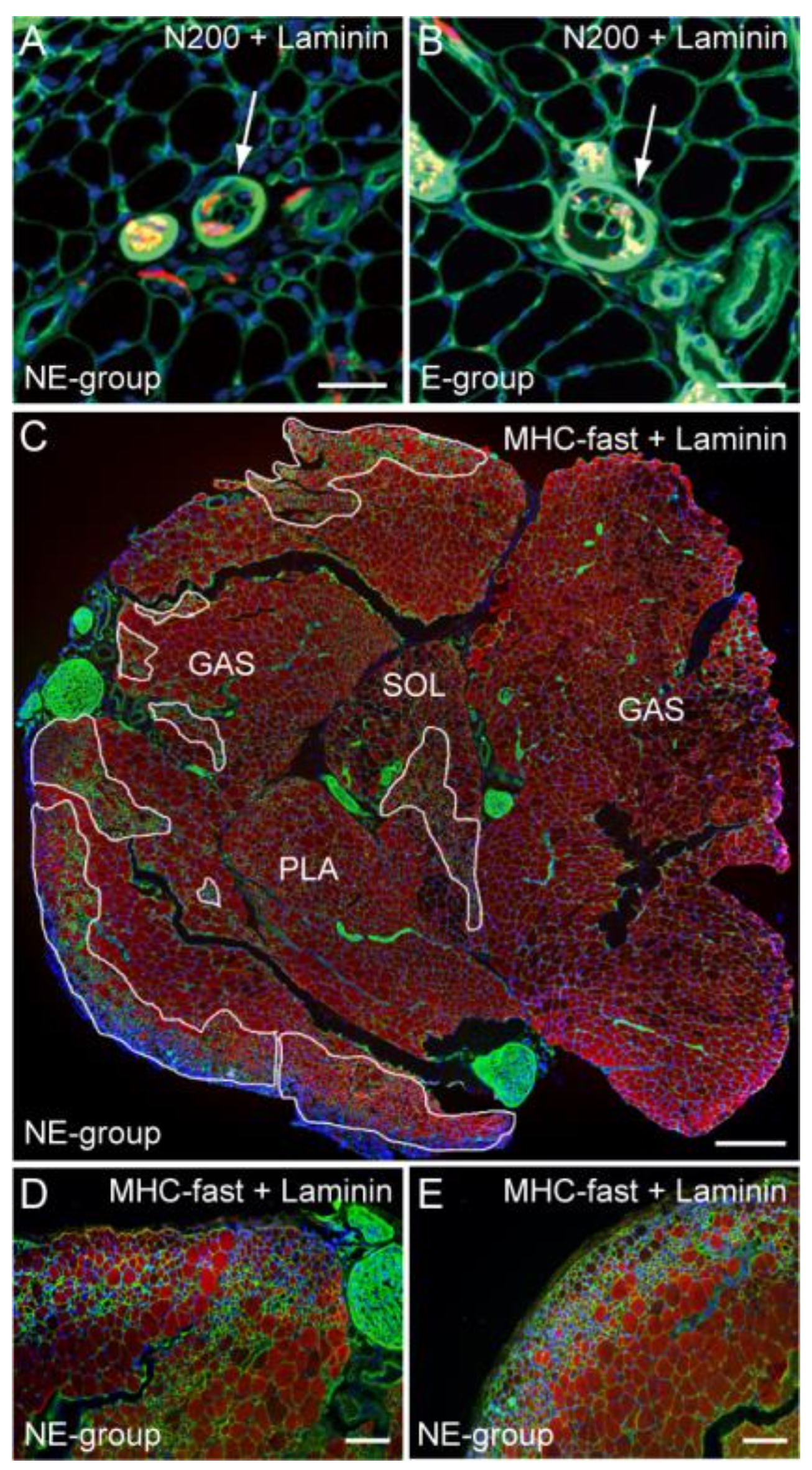
3.1. Recovery of the Number of Axon and Myelin in the Nerve Bridged Conduit
3.2. Recovery of Downstream Plantar Flexor Muscle Mass and Tension Output, and Individual Exercise Score during the Recovery Period
3.3. Axon Counts in the Downstream Tibial Nerve Branches
3.4. Characteristics of Downstream Muscle Spindle and Fibers
3.5. Differentiation of Transplanted Human Sk-34 Cells and Blood Vessel Formation in the Conduit
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Robinson, L.R. Traumatic injury to peripheral nerves. Muscle Nerve 2000, 23, 863–873. [Google Scholar] [CrossRef]
- Sunderland, S. The anatomy and physiology of nerve injury. Muscle Nerve 1990, 13, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Scholz, T.; Krichevsky, A.; Sumarto, A.; Jaffurs, D.; Wirth, G.A.; Paydar, K.; Evans, G.R. Peripheral nerve injuries: An international survey of current treatments and future perspectives. J. Reconstr. Microsurg. 2009, 25, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Braza, D.; Rice, J.B.; Dillingham, T. The incidence of peripheral nerve injury in extremity trauma. Am. J. Phys. Med. Rehabil. 2008, 87, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Hirata, M.; Soeda, S.; Nakajima, N.; Saito, K.; Nakazato, K.; Okada, Y.; Hashimoto, H.; Uchiyama, Y.; Mochida, J. Preferential and comprehensive reconstitution of severely damaged sciatic nerve using murine skeletal muscle-derived multipotent stem cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Hirata, M.; Nakajima, N.; Saito, K.; Hashimoto, H.; Soeda, S.; Uchiyama, Y.; Watanabe, M. A long-gap peripheral nerve injury therapy using human skeletal muscle-derived stem cells (Sk-SCS): An achievement of significant morphological, numerical and functional recovery. PLoS ONE 2016, 11, e0166639. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, M.J.; Redmon, N.; Schwartz, G.; English, A.W. Treadmill training promotes axon regeneration in injured peripheral nerves. Exp. Neurol. 2008, 211, 489–493. [Google Scholar] [CrossRef] [PubMed]
- English, A.W.; Cucoranu, D.; Mulligan, A.; Sabatier, M. Treadmill training enhances axon regeneration in injured mouse peripheral nerves without increased loss of topographic specificity. J. Comp. Neurol. 2009, 517, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Hoke, A. Treadmill exercise induced functional recovery after peripheral nerve repair is associated with increased levels of neurotrophic factors. PLoS ONE 2014, 9, e90245. [Google Scholar] [CrossRef] [PubMed]
- Cannoy, J.; Crowley, S.; Jarratt, A.; Werts, K.L.; Osborne, K.; Park, S.; English, A.W. Upslope treadmill exercise enhances motor axon regeneration but not functional recovery following peripheral nerve injury. J. Neurophysiol. 2016, 116, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Uchiyama, Y.; Hirata, M.; Hashimoto, H.; Nakajima, N.; Saito, K.; Terachi, T.; Mochida, J. Therapeutic isolation and expansion of human skeletal muscle-derived stem cells for the use of muscle-nerve-blood vessel reconstitution. Front. Physiol. 2015, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Akatsuka, A.; Ando, K.; Nakamura, Y.; Matsuzawa, H.; Hotta, T.; Roy, R.R.; Edgerton, V.R. Identification of myogenic-endothelial progenitor cells in the interstitial spaces of skeletal muscle. J. Cell Biol. 2002, 157, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Akatsuka, A.; Okada, Y.; Matsuzaki, Y.; Okano, H.; Kimura, M. Growth and differentiation potential of main- and side-population cells derived from murine skeletal muscle. Exp. Cell Res. 2003, 291, 83–90. [Google Scholar] [CrossRef]
- Chua, K.S.; Kuah, C.W. Innovating with rehabilitation technology in the real world: Promises, potentials, and perspectives. Am. J. Phys. Med. Rehabil. 2017, 96, S150–S156. [Google Scholar] [CrossRef] [PubMed]
- English, A.W.; Wilhelm, J.C.; Ward, P.J. Exercise, neurotrophins, and axon regeneration in the PNS. Physiology 2014, 29, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, J.C.; Xu, M.; Cucoranu, D.; Chmielewski, S.; Holmes, T.; Lau, K.S.; Bassell, G.J.; English, A.W. Cooperative roles of BDNF expression in neurons and Schwann cells are modulated by exercise to facilitate nerve regeneration. J. Neurosci. 2012, 32, 5002–5009. [Google Scholar] [CrossRef] [PubMed]
- Salerno, K.M.; Jing, X.; Diges, C.M.; Davis, B.M.; Albers, K.M. TRAF family member-associated NF-κB activator (TANK) expression increases in injured sensory neurons and is transcriptionally regulated by Sox11. Neuroscience 2013, 231, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.L.; Goldberg, J.L. Multiple transcription factor families regulate axon growth and regeneration. Dev. Neurobiol. 2011, 71, 1186–1211. [Google Scholar] [CrossRef] [PubMed]
- Struebing, F.L.; Wang, J.; Li, Y.; King, R.; Mistretta, O.C.; English, A.W.; Geisert, E.E. Differential expression of Sox11 and BDNF mRNA isoforms in the injured and regenerating nervous systems. Front. Mol. Neurosci. 2017, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; You, Y.; Gupta, V.B.; Klistorner, A.; Graham, S.L. TRKB receptor signalling: Implications in neurodegenerative, psychiatric and proliferative disorders. Int. J. Mol. Sci. 2013, 14, 10122–10142. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T. Therapeutic capacities of human and mouse skeletal muscle-derived stem cells for a long gap peripheral nerve injury. Neural Regen. Res. 2017, 12, 1811–1813. [Google Scholar] [CrossRef] [PubMed]
- Al-Majed, A.A.; Neumann, C.M.; Brushart, T.M.; Gordon, T. Brief electrical stimulation promotes the speed and accuracy of motor axonal regeneration. J. Neurosci. 2000, 20, 2602–2608. [Google Scholar] [PubMed]
- Geremia, N.M.; Gordon, T.; Brushart, T.M.; Al-Majed, A.A.; Verge, V.M. Electrical stimulation promotes sensory neuron regeneration and growth-associated gene expression. Exp. Neurol. 2007, 205, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T. Electrical stimulation to enhance axon regeneration after peripheral nerve injuries in animal models and humans. Neurotherapeutics 2016, 13, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Asensio-Pinilla, E.; Udina, E.; Jaramillo, J.; Navarro, X. Electrical stimulation combined with exercise increase axonal regeneration after peripheral nerve injury. Exp. Neurol. 2009, 219, 258–265. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Mass (g) | Muscle Mass (mg) * | Tension Output (N 1 × 102) * | |||
|---|---|---|---|---|---|
| Op-Side | Con-Side | Op-Side | Con-Side | ||
| E group | 24.3 ± 0.6 | 84.5 ± 7.5 | 133.0 ± 4.9 | 75.8 ± 11.2 | 81.2 ± 4.7 |
| NE group | 23.2 ± 0.4 | 74.5 ± 9.5 | 132.5 ± 3.9 | 57.4 ± 8.3 | 94.2 ± 9.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seta, H.; Maki, D.; Kazuno, A.; Yamato, I.; Nakajima, N.; Soeda, S.; Uchiyama, Y.; Tamaki, T. Voluntary Exercise Positively Affects the Recovery of Long-Nerve Gap Injury Following Tube-Bridging with Human Skeletal Muscle-Derived Stem Cell Transplantation. J. Clin. Med. 2018, 7, 67. https://doi.org/10.3390/jcm7040067
Seta H, Maki D, Kazuno A, Yamato I, Nakajima N, Soeda S, Uchiyama Y, Tamaki T. Voluntary Exercise Positively Affects the Recovery of Long-Nerve Gap Injury Following Tube-Bridging with Human Skeletal Muscle-Derived Stem Cell Transplantation. Journal of Clinical Medicine. 2018; 7(4):67. https://doi.org/10.3390/jcm7040067
Chicago/Turabian StyleSeta, Hiroya, Daisuke Maki, Akihito Kazuno, Ippei Yamato, Nobuyuki Nakajima, Shuichi Soeda, Yoshiyasu Uchiyama, and Tetsuro Tamaki. 2018. "Voluntary Exercise Positively Affects the Recovery of Long-Nerve Gap Injury Following Tube-Bridging with Human Skeletal Muscle-Derived Stem Cell Transplantation" Journal of Clinical Medicine 7, no. 4: 67. https://doi.org/10.3390/jcm7040067
APA StyleSeta, H., Maki, D., Kazuno, A., Yamato, I., Nakajima, N., Soeda, S., Uchiyama, Y., & Tamaki, T. (2018). Voluntary Exercise Positively Affects the Recovery of Long-Nerve Gap Injury Following Tube-Bridging with Human Skeletal Muscle-Derived Stem Cell Transplantation. Journal of Clinical Medicine, 7(4), 67. https://doi.org/10.3390/jcm7040067