Randomized Controlled Trial Considering Varied Exercises for Reducing Proactive Memory Interference


Abstract
:1. Introduction
1.1. What Is Proactive Interference?
1.2. Theoretical Overview
1.3. Biological Overview
1.4. Exercise and PI
2. Method
2.1. Participants
2.2. Study Design
2.3. Study 1: Walking
2.4. Study 2: Jogging
2.5. Materials
2.5.1. Retrospective Memory
2.5.2. Exercise
“Please select a pace similar to one you would choose if you were late to class. Thus, it will not be a leisurely walk. Nor will it be a run.”
2.6. Statistical Analysis
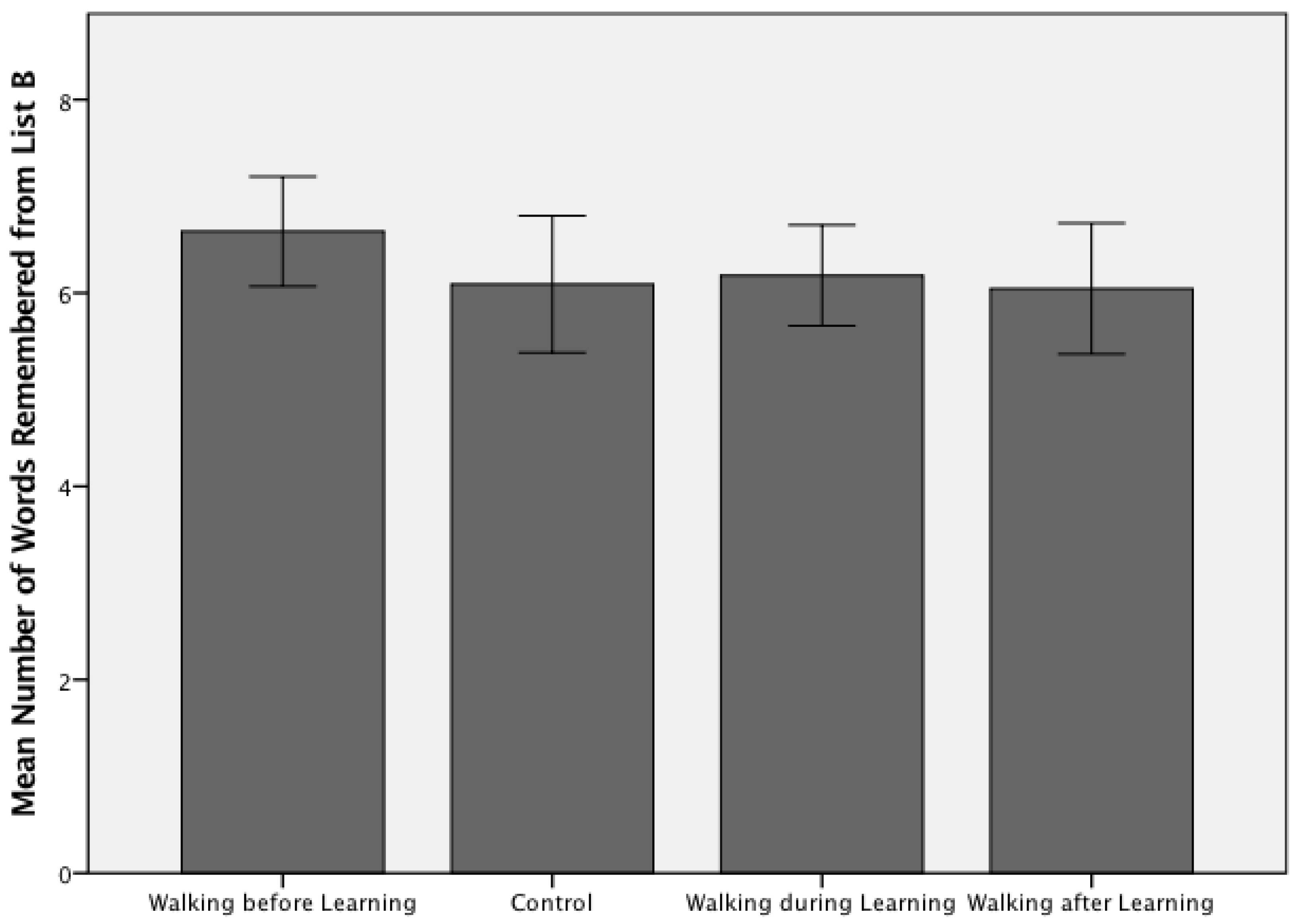
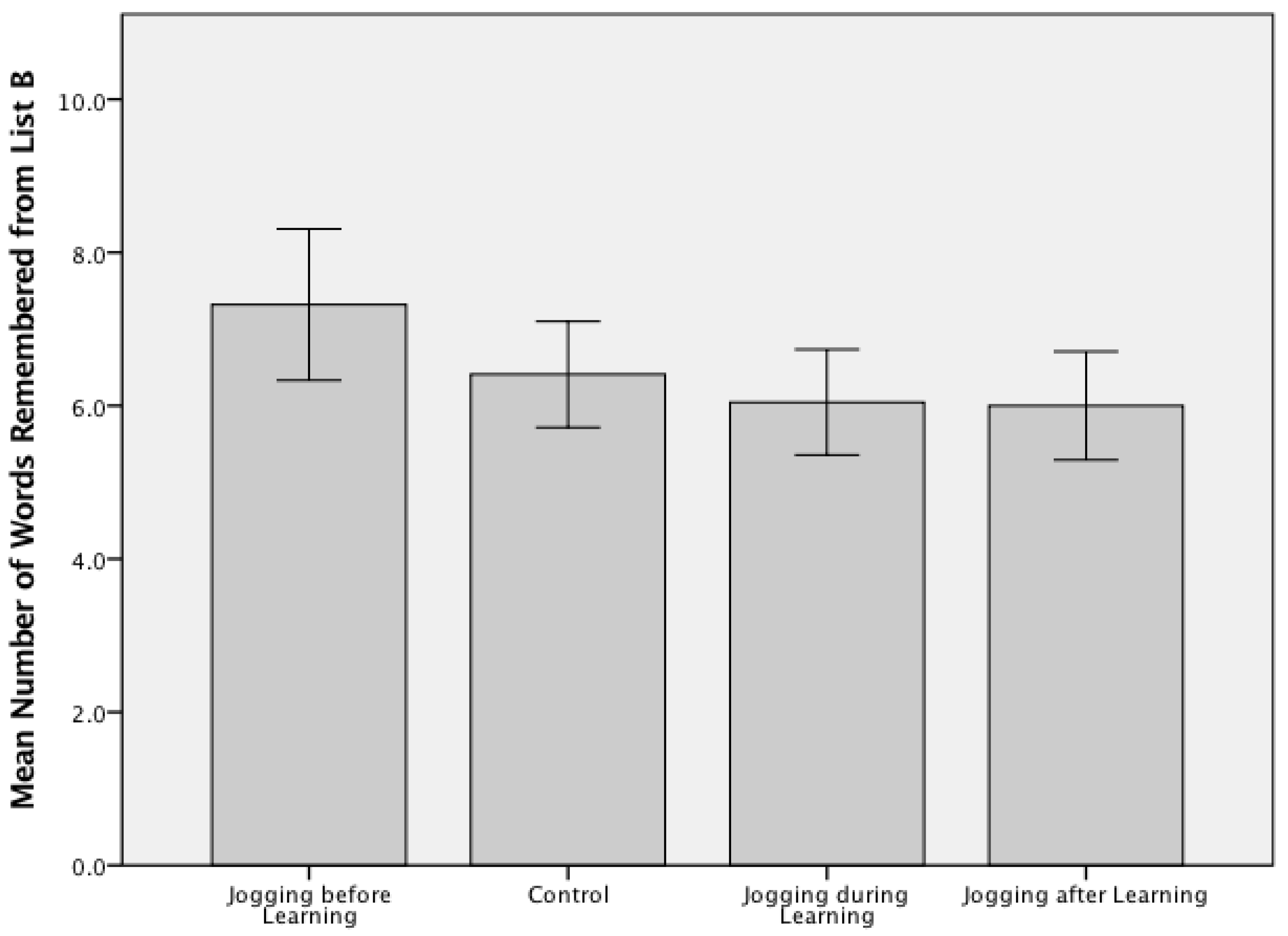
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Bennett, R.W. Proactive interference in short-term memory: Fundamental forgetting processes. J. Verbal Learn. Verbal Behav. 1975, 14, 123–144. [Google Scholar] [CrossRef]
- Bower, G.H. A Multicomponent Theory of the Memory Trace; Academic Press: New York, NY, USA, 1967. [Google Scholar]
- Turvey, M.T.; Egan, J.F. Release from proactlve interference in short-term memory as a function of change in visual and phonemic structure and retention interval. Percept. Psychophys. 1970, 7, 169–172. [Google Scholar] [CrossRef]
- Brown, J. Some tests of the decay theory of immediate memory. Q. J. Exp. Psychol. 1958, 10, 12–21. [Google Scholar] [CrossRef]
- Guise, K.G.; Shapiro, M.L. Medial prefrontal cortex reduces memory interference by modifying hippocampal encoding. Neuron 2017, 94, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Dolan, R.J.; Fletcher, P.C. Dissociating prefrontal and hippocampal function in episodic memory encoding. Nature 1997, 388, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Moscovitch, M. Multiple dissociations of function in amnesia. Hum. Memory Amnesia 1982, 337–370. [Google Scholar]
- Raichle, M.E.; Fiez, J.A.; Videen, T.O.; MacLeod, A.M.K.; Pardo, J.V.; Fox, P.T.; Petersen, S.E. Practice-related changes in human brain functional anatomy during nonmotor learning. Cereb. Cortex 1994, 4, 8–26. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, A.P.; Jurica, P.J.; Mangels, J.A.; Gershberg, F.B.; Knight, R.T. Susceptibility to memory interference effects following frontal lobe damage: Findings from tests of paired-associate learning. J. Cognit. Neurosci. 1995, 7, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Schill, S.L.J.J.; Marshuetz, C.; Smith, E.E.; D’Esposito, M.; Kan, I.P.; Knight, R.T.; Swick, D. Effects of frontal lobe damage on interference effects in working memory. Cognit. Affect Behav. Neurosci. 2002, 2, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Miller, E.K.; Desimone, R. The representation of stimulus familiarity in anterior inferior temporal cortex. J. Neurophysiol. 1993, 69, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Badre, D.; Wagner, A.D. Selection, integration, and conflict monitoring; assessing the nature and generality of prefrontal cognitive control mechanisms. Neuron 2004, 41, 473–487. [Google Scholar] [CrossRef]
- Badre, D.; Wagner, A.D. Frontal lobe mechanisms that resolve proactive interference. Cereb. Cortex 2005, 15, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, M.; Postle, B.R.; Jonides, J.; Smith, E.E. The neural substrate and temporal dynamics of interference effects in working memory as revealed by event-related functional MRI. Proc. Natl. Acad Sci. USA 1999, 96, 7514–7519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonides, J.; Smith, E.E.; Marshuetz, C.; Koeppe, R.A.; Reuter-Lorenz, P.A. Inhibition in verbal working memory revealed by brain activation. Proc. Natl. Acad Sci. USA 1998, 95, 8410–8413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, J.K.; Reuter-Lorenz, P.A.; Sylvester, C.Y.; Jonides, J.; Smith, E.E. Dissociable neural mechanisms underlying response-based and familiarity-based conflict in working memory. Proc. Natl. Acad Sci. USA 2003, 100, 11171–11175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jay, T.M. Dopamine: A potential substrate for synaptic plasticity and memory mechanisms. Prog. Neurobiol. 2003, 69, 375–390. [Google Scholar] [CrossRef]
- Winters, B.D.; Bartko, S.J.; Saksida, L.M.; Bussey, T.J. Scopolamine infused into perirhinal cortex improves object recognition memory by blocking the acquisition of interfering object information. Learn Mem. 2007, 14, 590–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.D.; Desmond, J.E.; Glover, G.H.; Gabrieli, J.D. Prefrontal cortex and recognition memory. Functional-MRI evidence for context-dependent retrieval processes. Brain 1998, 121 Pt 10, 1985–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNerney, M.W.; Radvansky, G.A. Mind racing: The influence of exercise on long-term memory consolidation. Memory 2015, 23, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Colcombe, S.J.; Erickson, K.I.; Raz, N.; Webb, A.G.; Cohen, N.J.; McAuley, E.; Kramer, A.F. Aerobic fitness reduces brain tissue loss in aging humans. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Brockett, A.T.; LaMarca, E.A.; Gould, E. Physical exercise enhances cognitive flexibility as well as astrocytic and synaptic markers in the medial prefrontal cortex. PLoS ONE 2015, 10, e0124859. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Davey, C.P. Physical exertion and mental performance. Ergonomics 1973, 16, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Tomporowski, P.D.; Cureton, K.; Armstrong, L.E.; Kane, G.M.S.P.B.; Millard-Stafford, M. Short-term effects of aerobic exercise on executive processes and emotional reactivity. Int. J. Sport Exerc. Psychol. 2005, 3, 131–146. [Google Scholar] [CrossRef]
- Etnier, J.L.; Nowell, P.M.; Landers, D.M.; Sibley, B.A. A meta-regression to examine the relationship between aerobic fitness and cognitive performance. Brain Res. Rev. 2006, 52, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Colcombe, S.J.; Kramer, A.F.; McAuley, E.; Erickson, K.I.; Scalf, P. Neurocognitive aging and cardiovascular fitness: Recent findings and future directions. J. Mol. Neurosci. 2004, 24, 9–14. [Google Scholar] [CrossRef]
- Labban, J.D.; Etnier, J.L. Effects of acute exercise on long-term memory. Res. Q. Exerc. Sport. 2011, 82, 712–721. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.; Sproule, J.; Turner, A.; Hale, B.J. Acute, intermediate intensity exercise, and speed and accuracy in working memory tasks: A meta-analytical comparison of effects. Physiol. Behav. 2011, 102, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A. Functional neuroanatomy of altered states of consciousness: The transient hypofrontality hypothesis. Cons. Cogn. 2003, 12, 231–256. [Google Scholar] [CrossRef] [Green Version]
- Loprinzi, P.D.; Kane, C.J. Exercise and cognitive function: A randomized controlled trial examining acute exercise and free-living physical activity and sedentary effects. Mayo Clinic Proc. 2015, 90, 450–460. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Magalhães, S.; Malloy-Diniz, L.F.; Hamdan, A.C. Validity convergent and reliability test-retest of the Rey Auditory Verbal Learning Test. Clinic. Neuropsychiatr. 2012, 9, 129–137. [Google Scholar]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Glanzer, M. Short-Term Memory; Academic Press: New York, NY, USA, 1982. [Google Scholar]
- Murdock, B.B. The serial position effect of free recall. J. Exp. Psychol. 1962, 64, 482–488. [Google Scholar] [CrossRef]
- Kiss, G.R.; Armstrong, C.; Milroy, R.; Piper, J. An associative thesaurus of English and its computer analysis. In The Computer and Literary Studies Edinburgh; Edinburgh University Press: Edinburgh, UK, 1973. [Google Scholar]
- Nielson, K.A.; Radtke, R.C.; Jensen, R.A. Arousal-induced modulation of memory storage processes in humans. Neurobiol. Learn Mem. 1996, 66, 133–142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study 1 (n = 88) Walking | Study 2 (n = 88) Jogging | |
|---|---|---|
| Age (years) | M = 22.3 ± 3.7 | M = 21.9 ± 2.4 |
| Body Mass Index (kg/m2) | M = 25.3 ± 4.5 | M = 24.2 ± 4.2 |
| Sex | ||
| Male | 52.3% | 45.5% |
| Female | 47.7% | 54.5% |
| Race | ||
| Caucasian/Nonhispanic White | 65.9% | 68.2% |
| African American/Nonhispanic Black | 20.5% | 25.0% |
| Other Hispanic | - | 1.1% |
| Other/multirace (Asian) | 13.6% | 5.7% |
| Speed (mph; n = 66) | ||
| Walking | M = 3.2 ± 5 | - |
| 0–5 min into jogging | - | M = 4.6 ± 9 |
| 6–10 min into jogging | - | M = 5.7 ± 9 |
| 11–15 min into jogging | - | M = 7.0 ± 1.3 |
| Heart rate | ||
| Resting (n = 88) | M = 73.5 ± 13.1 | M = 71.9 ± 10.9 |
| 7.5 min into walking/jogging (n = 66; control group omitted) | M = 113.3 ± 21.1 | M = 163.6 ± 16.5 |
| 14 min into walking/jogging | M = 116.0 ± 20.1 | M = 183.7 ± 12.9 |
| 5 min after walking/jogging | M = 80.6 ± 13.7 | M = 106.4 ± 15.8 |
| Rating of perceived exertion | ||
| 7.5 min into walking (n = 66; control group omitted) | M = 9.1 ± 1.4 | - |
| 14 min into walking | M = 11.9 ± 14.5 | - |
| 0–5 min into jogging | - | M = 11.4 ± 7 |
| 6–10 min into jogging | - | M = 14.7 ± 1.2 |
| 11–15 min into jogging | - | M = 18.7 ± 9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frith, E.; Sng, E.; Loprinzi, P.D. Randomized Controlled Trial Considering Varied Exercises for Reducing Proactive Memory Interference. J. Clin. Med. 2018, 7, 147. https://doi.org/10.3390/jcm7060147
Frith E, Sng E, Loprinzi PD. Randomized Controlled Trial Considering Varied Exercises for Reducing Proactive Memory Interference. Journal of Clinical Medicine. 2018; 7(6):147. https://doi.org/10.3390/jcm7060147
Chicago/Turabian StyleFrith, Emily, Eveleen Sng, and Paul D. Loprinzi. 2018. "Randomized Controlled Trial Considering Varied Exercises for Reducing Proactive Memory Interference" Journal of Clinical Medicine 7, no. 6: 147. https://doi.org/10.3390/jcm7060147
APA StyleFrith, E., Sng, E., & Loprinzi, P. D. (2018). Randomized Controlled Trial Considering Varied Exercises for Reducing Proactive Memory Interference. Journal of Clinical Medicine, 7(6), 147. https://doi.org/10.3390/jcm7060147