Indocyanine Green—Mediated Photodynamic Therapy Reduces Methicillin-Resistant Staphylococcus aureus Drug Resistance

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Photodynamic Therapy System and ICG Photoactivation
2.3. Growth Curves
2.4. Antibiotic Susceptibility Testing
2.5. Transmission Electron Microscopy
2.6. Polymerase Chain Reaction
2.7. Quantitative Reverse Transcription Polymerase Chain Reaction
2.8. Western Blot
2.9. In Vivo
2.9.1. Murine Skin Infection Model
2.9.2. Murine Pneumonia Model
2.10. Statistics
3. Results
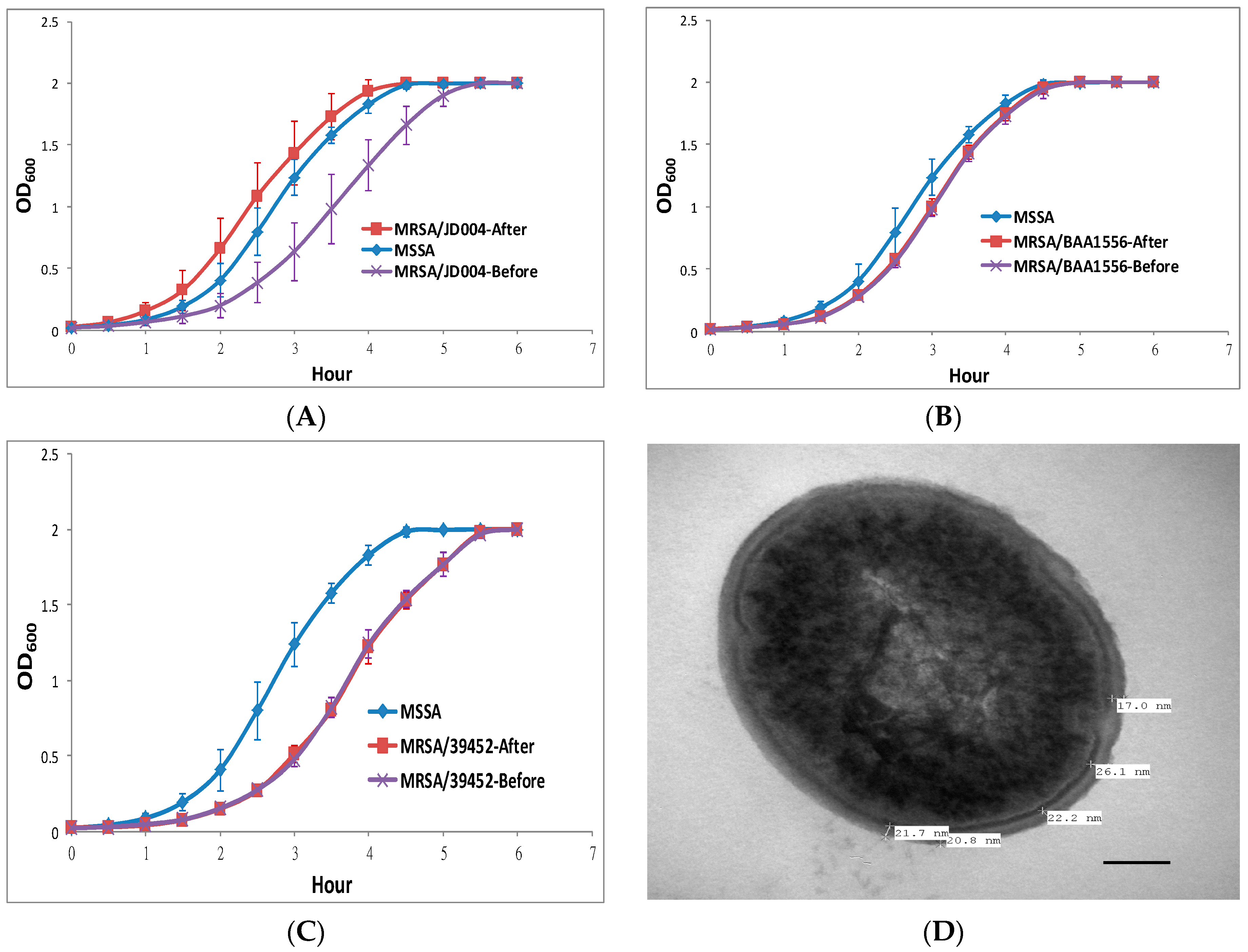
3.1. Photodynamic Inactivation on Three MRSA Strains
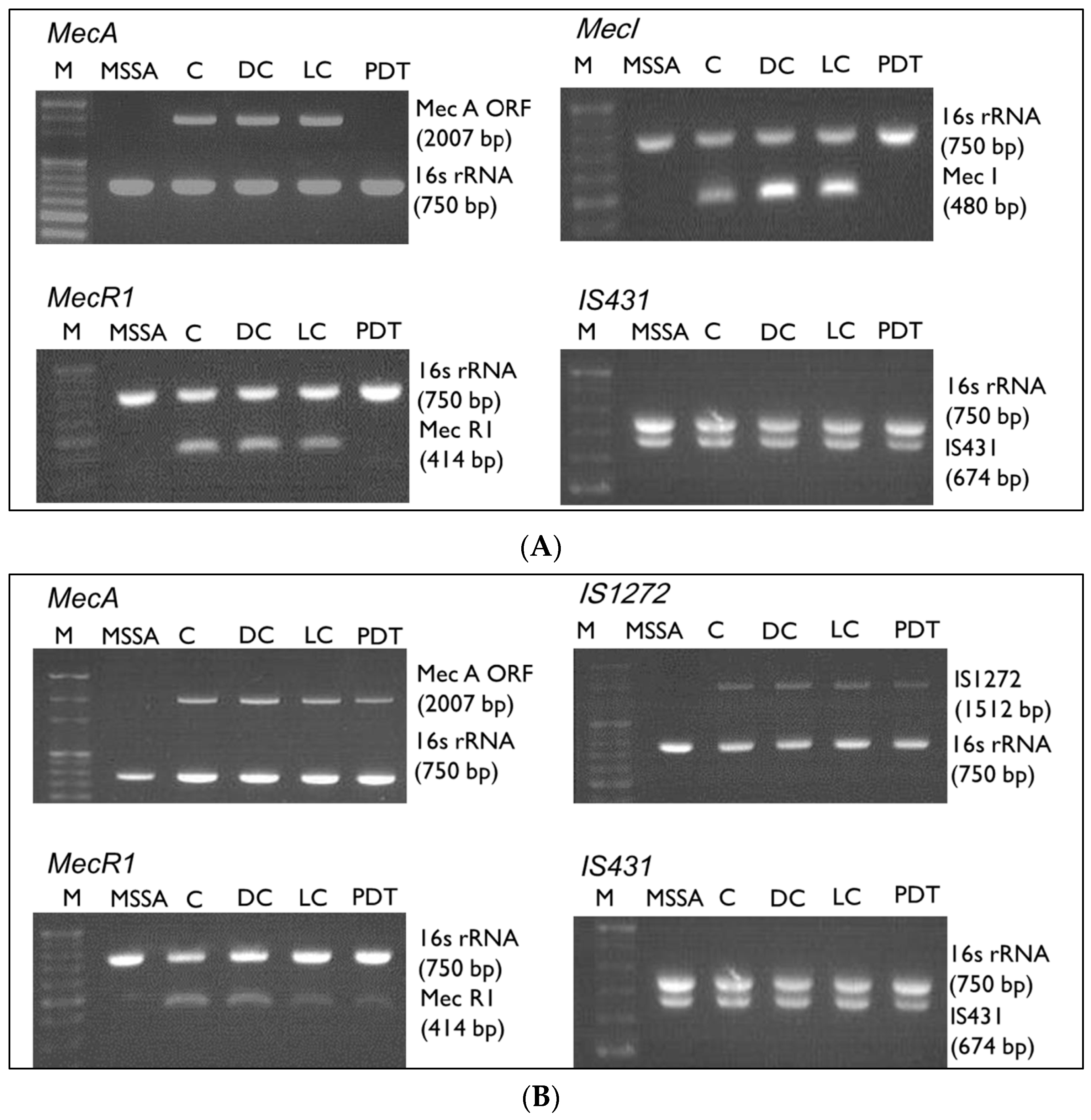
3.2. mec Complex Typing of MRSA Strains
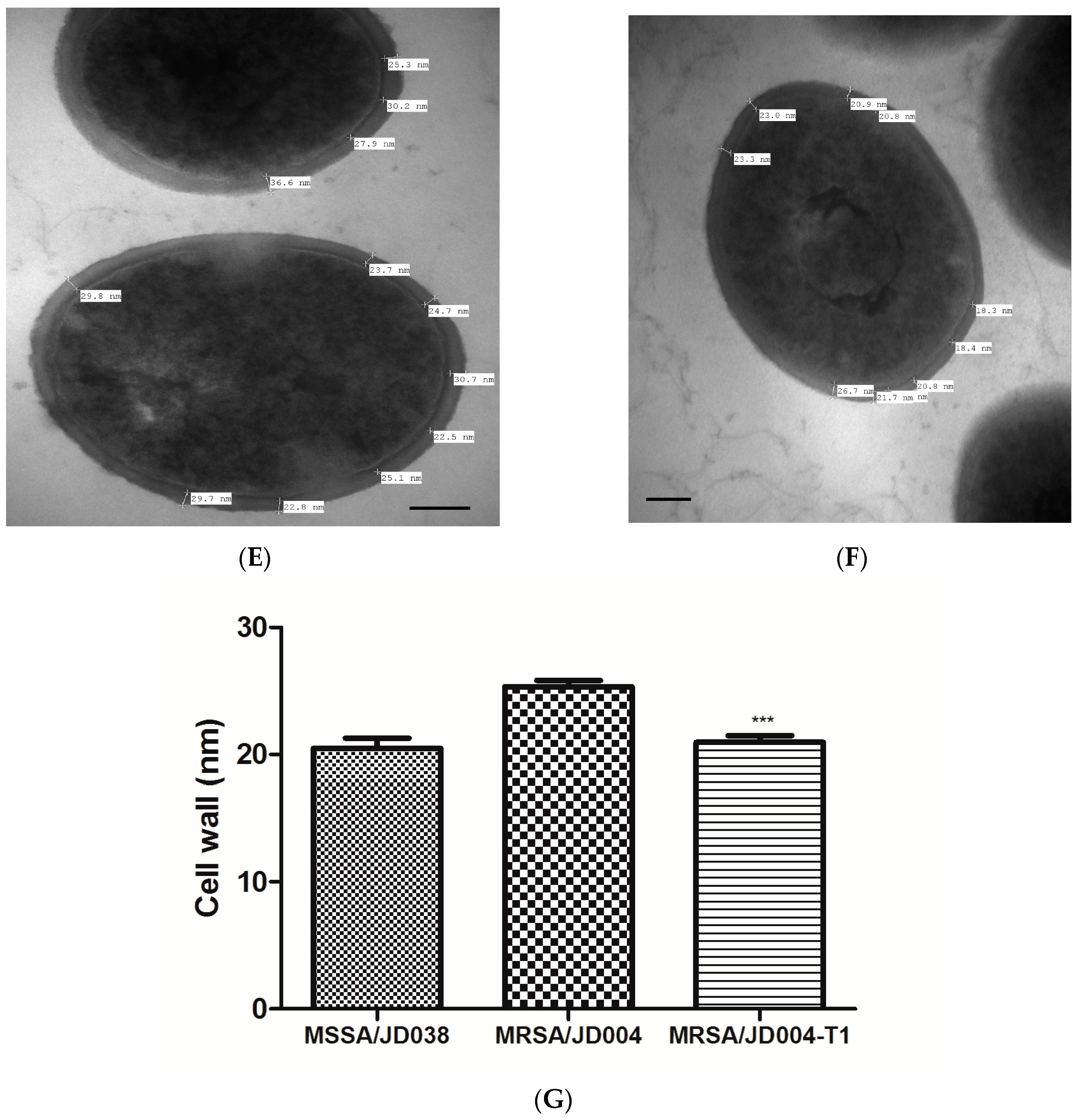
3.3. Physiological MRSA Changes after ICG-PDT
3.4. Antimicrobial Susceptibility Phenotype Altered by ICG-PDT
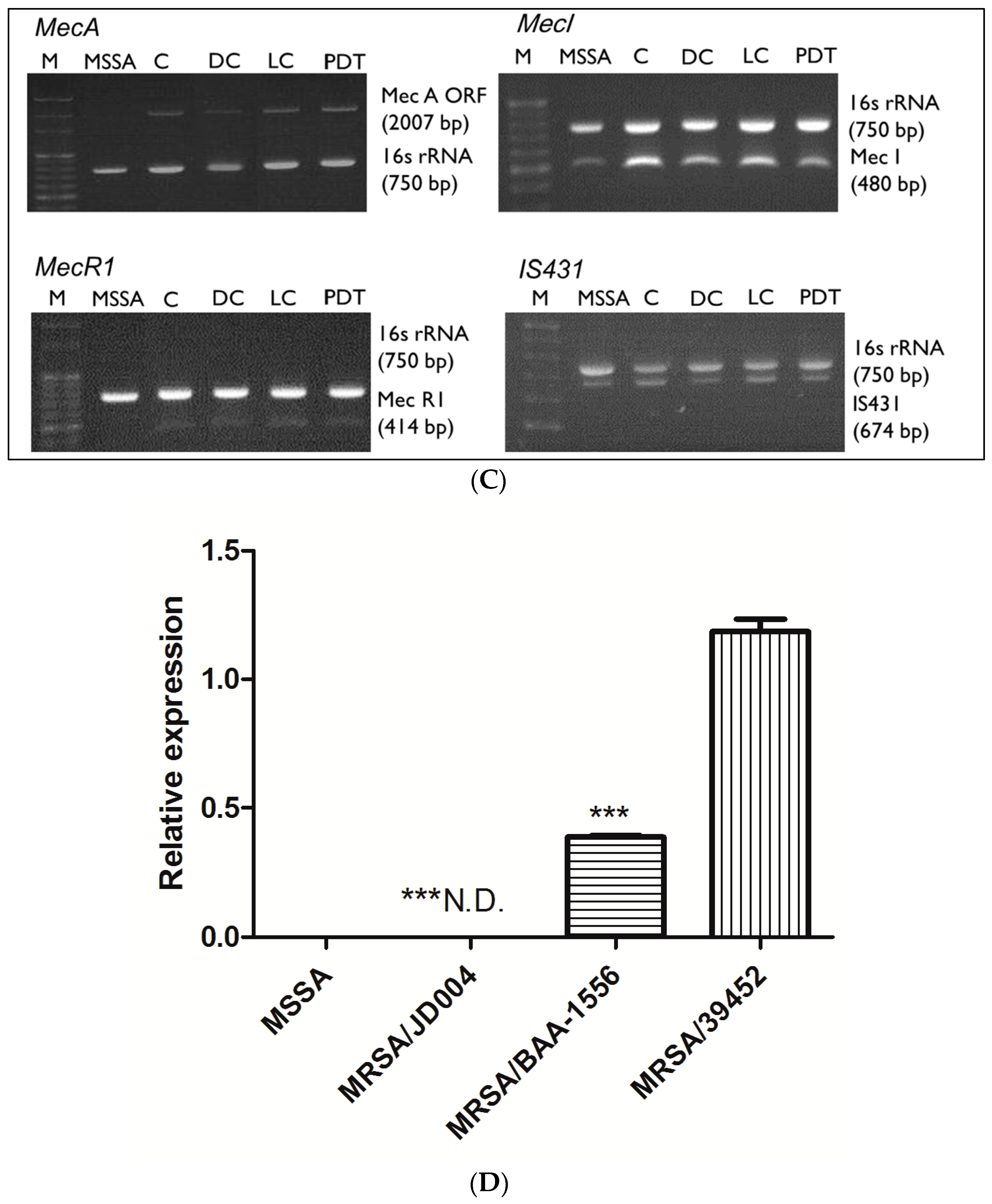
3.5. Gene Alterations after ICG-PDT
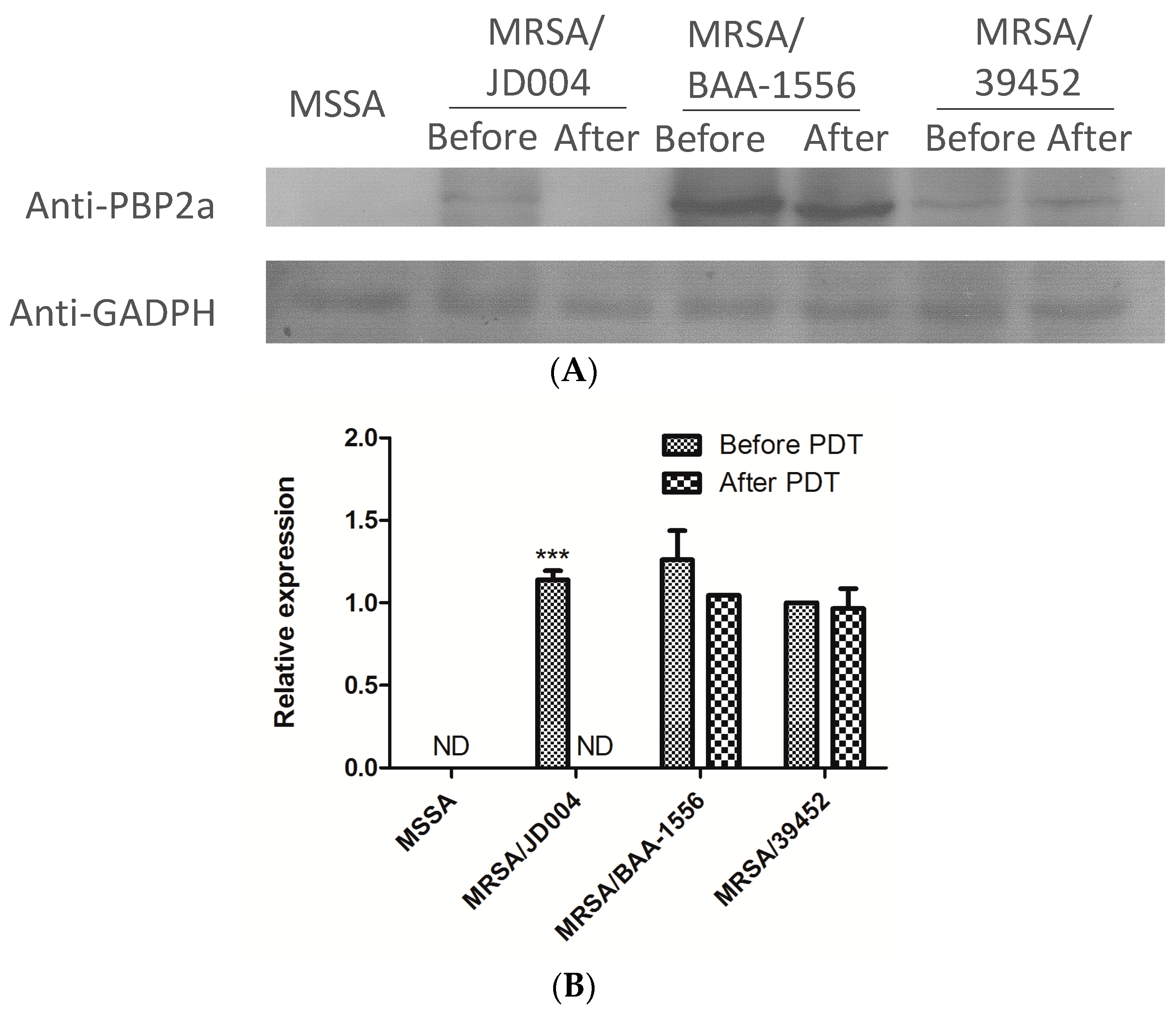
3.6. Reduction of PBP2a by ICG-PDT
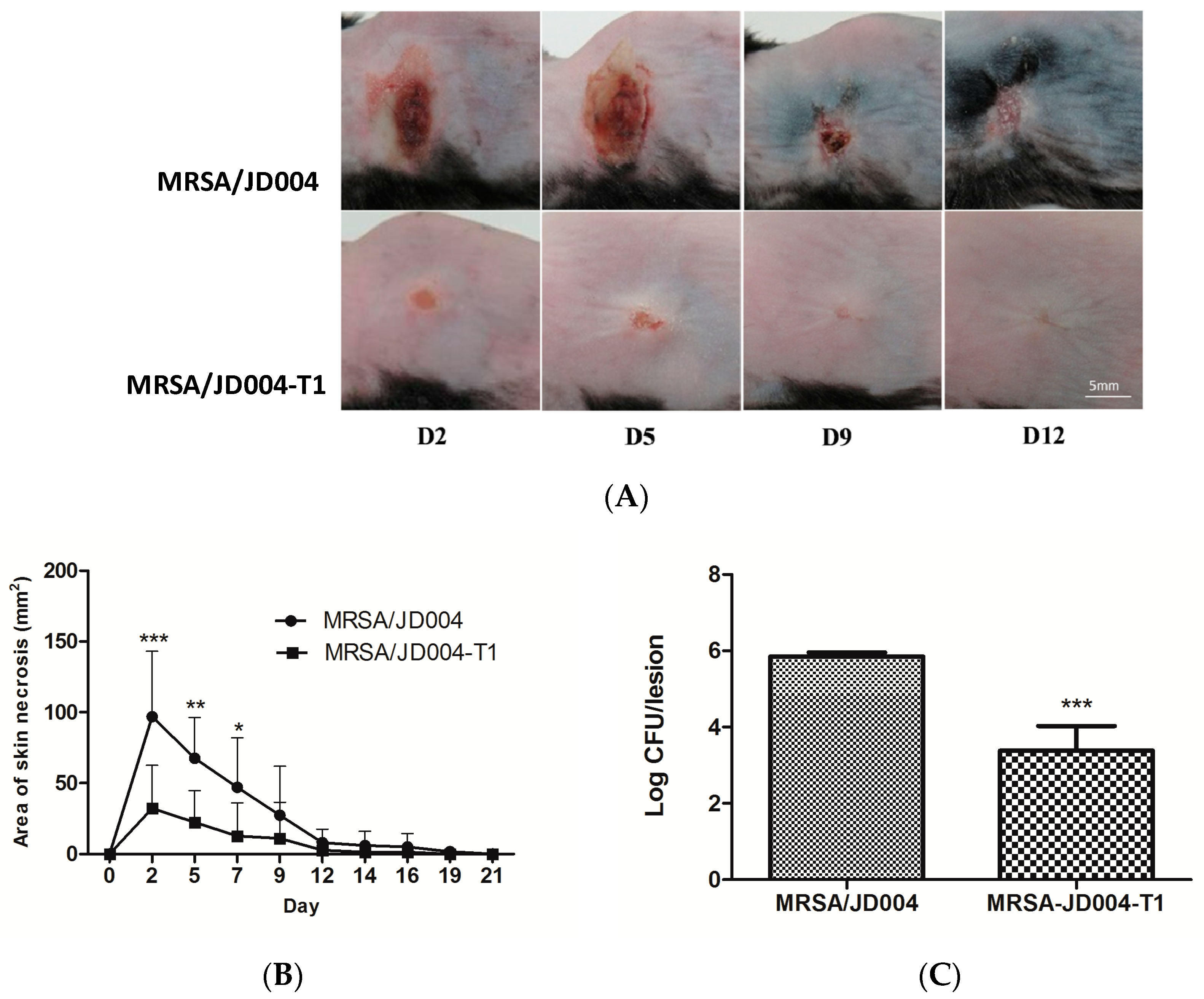
3.7. Oxacillin Decreased Skin Necrosis in Mice Infected with MRSA post-ICG-PDT
3.8. Oxacillin Decreased Pulmonary Bacterial Burden Following Infection with ICG-PDT/PTT-Treated MRSA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [PubMed]
- Anantha, R.V.; Jegatheswaran, J.; Pepe, D.L.; Priestap, F.; Delport, J.; Haeryfar, S.M.; McCormick, J.K.; Mele, T. Risk factors for mortality among patients with Staphylococcus aureus bacteremia: A single-centre retrospective cohort study. Can. Med. Assoc. Open Access J. 2014, 2, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Tipper, D. Mode of action of β-lactam antibiotics. Rev. Infect. Dis. 1979, 1, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F. The changing epidemiology of Staphylococcus aureus? Emerg. Infect. Dis. 2001, 7, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Hartman, B.J.; Tomasz, A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J. Bacteriol. 1984, 158, 513–516. [Google Scholar] [PubMed]
- Shang, W.; Davies, T.A.; Flamm, R.K.; Bush, K. Effects of ceftobiprole and oxacillin on mecA expression in methicillin-resistant Staphylococcus aureus clinical isolates. Antimicrob. Agents Chemother. 2010, 54, 956–959. [Google Scholar] [CrossRef]
- Oppegaard, H.; Steinum, T.M.; Wasteson, Y. Horizontal transfer of a multi-drug resistance plasmid between coliform bacteria of human and bovine origin in a farm environment. Appl. Environ. Microbiol. 2001, 67, 3732–3734. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Weigel, L.M.; Appelbaum, P.C.; McDougal, L.K.; Chaitram, J.; McAllister, S.; Clark, N.; Killgore, G.; O‘Hara, C.M.; Jevitt, L.; et al. Vancomycin-resistant Staphylococcus aureus isolate from a patient in Pennsylvania. Antimicrob. Agents Chemother. 2004, 48, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Sievert, D.M.; Hageman, J.C.; Boulton, M.L.; Tenover, F.C.; Downes, F.P.; Shah, S.; Rudrik, J.T.; Pupp, G.R.; Brown, W.J.; et al. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N. Engl. J. Med. 2003, 348, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Maisch, T.; Baier, J.; Franz, B.; Maier, M.; Landthaler, M.; Szeimies, R.M.; Baumler, W. The role of singlet oxygen and oxygen concentration in photodynamic inactivation of bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 7223–7228. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial photodynamic therapy-what we know and what we don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Kashef, N.; Hamblin, M.R. Can microbial cells develop resistance to oxidative stress in antimicrobial photodynamic inactivation? Drug Resist. Update 2017, 31, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, M.; Maisch, T.; Nonell, S.; Plaetzer, K.; Almeida, A.; Tegos, G.P.; Hamblin, M.R. Photoantimicrobials-are we afraid of the light? Lancet Infect. Dis. 2017, 17, 49–55. [Google Scholar] [CrossRef]
- Dai, T.; Huang, Y.-Y.; Hamblin, M.R. Photodynamic therapy for localized infections—State of the art. Photodiagn. Photodyn. Ther. 2009, 6, 170–188. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.; Capella, M.; Caldas, L. Photodynamic action of methylene blue: Repair and mutation in Escherichia coli. J. Photochem. Photobiol. B Biol. 1990, 5, 505–517. [Google Scholar] [CrossRef]
- Wakayama, Y.; Takagi, M.; Yano, K. Photosensitized inactivation of e. coli cells in toluidine blue-light system. Photochem. Photobiol. 1980, 32, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.-W.; Wang, Y.-Y.; Sheu, H.-M.; Chuang, Y.-C. Bactericidal effects of toluidine blue-mediated photodynamic action on Vibrio vulnificus. Antimicrob. Agents Chemoth. 2005, 49, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Kömerik, N.; Wilson, M.; Poole, S. The Effect of Photodynamic Action on Two Virulence Factors of Gram-negative Bacteria. Photochem. Photobiol. 2000, 72, 676–680. [Google Scholar] [CrossRef]
- Guyer, D.R.; Duker, J.S.; Puliafito, C.A. Indocyanine green angiography and dye-enhanced diode laser photocoagulation. Semin. Ophthalmol. 1992, 7, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Desmettre, T.; Devoisselle, J.; Mordon, S. Fluorescence properties and metabolic features of indocyanine green (ICG) as related to angiography. Surv. Ophthalmol. 2000, 45, 15–27. [Google Scholar] [CrossRef]
- Urbanska, K.; Romanowska-Dixon, B.; Matuszak, Z.; Oszajca, J.; Nowak-Sliwinska, P.; Stochel, G. Indocyanine green as a prospective sensitizer for photodynamic therapy of melanomas. Acta Biochim. Polonica 2001, 49, 387–391. [Google Scholar]
- Okochi, O.; Kaneko, T.; Sugimoto, H.; Inoue, S.; Takeda, S.; Nakao, A. ICG pulse spectrophotometry for perioperative liver function in hepatectomy. J. Surg. Res. 2002, 103, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Allison, R.R.; Downie, G.H.; Cuenca, R.; Hu, X.-H.; Childs, C.J.H.; Sibata, C.H. Photosensitizers in clinical PDT. Photodiagn. Photodyn. Ther. 2004, 1, 27–42. [Google Scholar] [CrossRef]
- Wong, T.W.; Wu, E.C.; Ko, W.C.; Lee, C.C.; Hor, L.I.; Huang, I.H. Photodynamic inactivation of methicillin-resistant Staphylococcus aureus by indocyanine green and near infrared light. Dematolog. Sin. 2017, 36, 1–32. [Google Scholar] [CrossRef]
- Ruhi, M.K.; Ak, A.; Gulsoy, M. Dose-dependent photochemical/photothermal toxicity of indocyanine green-based therapy on three different cancer cell lines. Photodiagn. Photodyn. Ther. 2018, 21, 334–343. [Google Scholar] [CrossRef]
- Giraudeau, C.; Moussaron, A.; Stallivieri, A.; Mordon, S.; Frochot, C. Indocyanine green: Photosensitizer or chromophore? Still a debate. Curr. Med. Chem. 2014, 21, 1871–1897. [Google Scholar] [CrossRef] [PubMed]
- Abels, C.; Karrer, S.; Baumler, W.; Goetz, A.E.; Landthaler, M.; Szeimies, R.M. Indocyanine green and laser light for the treatment of AIDS-associated cutaneous Kaposi’s sarcoma. Br. J. Cancer 1998, 77, 1021–1024. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Chiniforush, N.; Shahabi, S.; Ghorbanzadeh, R.; Bahador, A. Sub-lethal doses of photodynamic therapy affect biofilm formation ability and metabolic activity of Enterococcus faecalis. Photodiagn. Photodyn. Ther. 2016, 15, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Beltes, C.; Sakkas, H.; Economides, N.; Papadopoulou, C. Antimicrobial photodynamic therapy using Indocyanine green and near-infrared diode laser in reducing Entrerococcus faecalis. Photodiagn. Photodyn. Ther. 2017, 17, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Hayashi, J.I.; Fujimura, T.; Iwamura, Y.; Yamamoto, G.; Nishida, E.; Ohno, T.; Okada, K.; Yamamoto, H.; Kikuchi, T.; et al. New Irradiation Method with Indocyanine Green-Loaded Nanospheres for Inactivating Periodontal Pathogens. Int. J. Mol. Sci. 2017, 18, 154. [Google Scholar] [CrossRef] [PubMed]
- Omar, G.S.; Wilson, M.; Nair, S.P. Lethal photosensitization of wound-associated microbes using indocyanine green and near-infrared light. BMC Microbiol. 2008, 8, 111. [Google Scholar] [CrossRef]
- Topaloglu, N.; Gulsoy, M.; Yuksel, S. Antimicrobial photodynamic therapy of resistant bacterial strains by indocyanine green and 809-nm diode laser. Photomed. Laser Surg. 2013, 31, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Weller, T.M. The distribution of mecA, mecR1 and mecI and sequence analysis of mecI and the mec promoter region in staphylococci expressing resistance to methicillin. J. Antimicrob. Chemother. 1999, 43, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Meng, J.R.; Niu, C.; Wang, H.F.; Liu, J.; Hu, B.Q.; Jia, M.; Luo, X.X. Restoration of antibiotic susceptibility in methicillin-resistant Staphylococcus aureus by targeting mecR1 with a phosphorothioate deoxyribozyme. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1160–1164. [Google Scholar] [CrossRef]
- Jo, D.S.; Montgomery, C.P.; Yin, S.; Boyle-Vavra, S.; Daum, R.S. Improved oxacillin treatment outcomes in experimental skin and lung infection by a methicillin-resistant Staphylococcus aureus isolate with a vraSR operon deletion. Antimicrob. Agents Chemother. 2011, 55, 2818–2823. [Google Scholar] [CrossRef]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef]
- Cui, L.; Ma, X.; Sato, K.; Okuma, K.; Tenover, F.C.; Mamizuka, E.M.; Gemmell, C.G.; Kim, M.N.; Ploy, M.C.; El Solh, N.; et al. Cell Wall Thickening Is a Common Feature of Vancomycin Resistance in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 5–14. [Google Scholar] [CrossRef]
- Ender, M.; McCallum, N.; Adhikari, R.; Berger-Bachi, B. Fitness cost of SCCmec and methicillin resistance levels in Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 2295–2297. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Hu, Q.; Cheng, H.; Shang, W.; Liu, N.; Hua, Z.; Zhu, J.; Hu, Z.; Yuan, J.; Zhang, X.; et al. Cell wall thickening is associated with adaptive resistance to amikacin in methicillin-resistant Staphylococcus aureus clinical isolates. J. Antimicrob. Chemother. 2013, 68, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J.; Huang, I.W.; Wang, C.H.; Chen, P.C.; Wang, H.Y.; Lai, J.F.; Shiau, Y.R.; Lauderdale, T.L.; TSAR Hospitals. mecA-positive Staphylococcus aureus with low-level oxacillin MIC in Taiwan. J. Clin. Microbiol. 2012, 50, 1679–1683. [Google Scholar] [CrossRef] [PubMed]
- Iluz, N.; Maor, Y.; Keller, N.; Malik, Z. The synergistic effect of PDT and oxacillin on clinical isolates of Staphylococcus aureus. Lasers Surg. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Standards, N.C.f.C.L. Performance Standards for Antimicrobial Susceptibility Testing; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 1992. [Google Scholar]
- Fernandes, C.J.; Fernandes, L.A.; Collignon, P.; Resistance, A.G.o.A. Cefoxitin resistance as a surrogate marker for the detection of methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2005, 55, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.P.; Teng, L.J.; Chen, C.T.; Lo, T.H.; Hung, W.C.; Chen, H.J.; Hsueh, P.R.; Tsai, J.C. Toluidine blue O photodynamic inactivation on multidrug-resistant Pseudomonas aeruginosa. Lasers Surg. Med. 2009, 41, 391–397. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, T.-W.; Liao, S.-Z.; Ko, W.-C.; Wu, C.-J.; Wu, S.B.; Chuang, Y.-C.; Huang, I.-H. Indocyanine Green—Mediated Photodynamic Therapy Reduces Methicillin-Resistant Staphylococcus aureus Drug Resistance. J. Clin. Med. 2019, 8, 411. https://doi.org/10.3390/jcm8030411
Wong T-W, Liao S-Z, Ko W-C, Wu C-J, Wu SB, Chuang Y-C, Huang I-H. Indocyanine Green—Mediated Photodynamic Therapy Reduces Methicillin-Resistant Staphylococcus aureus Drug Resistance. Journal of Clinical Medicine. 2019; 8(3):411. https://doi.org/10.3390/jcm8030411
Chicago/Turabian StyleWong, Tak-Wah, Shu-Zhen Liao, Wen-Chien Ko, Chi-Jung Wu, Shin Bei Wu, Yin-Ching Chuang, and I-Hsiu Huang. 2019. "Indocyanine Green—Mediated Photodynamic Therapy Reduces Methicillin-Resistant Staphylococcus aureus Drug Resistance" Journal of Clinical Medicine 8, no. 3: 411. https://doi.org/10.3390/jcm8030411
APA StyleWong, T. -W., Liao, S. -Z., Ko, W. -C., Wu, C. -J., Wu, S. B., Chuang, Y. -C., & Huang, I. -H. (2019). Indocyanine Green—Mediated Photodynamic Therapy Reduces Methicillin-Resistant Staphylococcus aureus Drug Resistance. Journal of Clinical Medicine, 8(3), 411. https://doi.org/10.3390/jcm8030411