Presbycusis: An Update on Cochlear Mechanisms and Therapies



Abstract
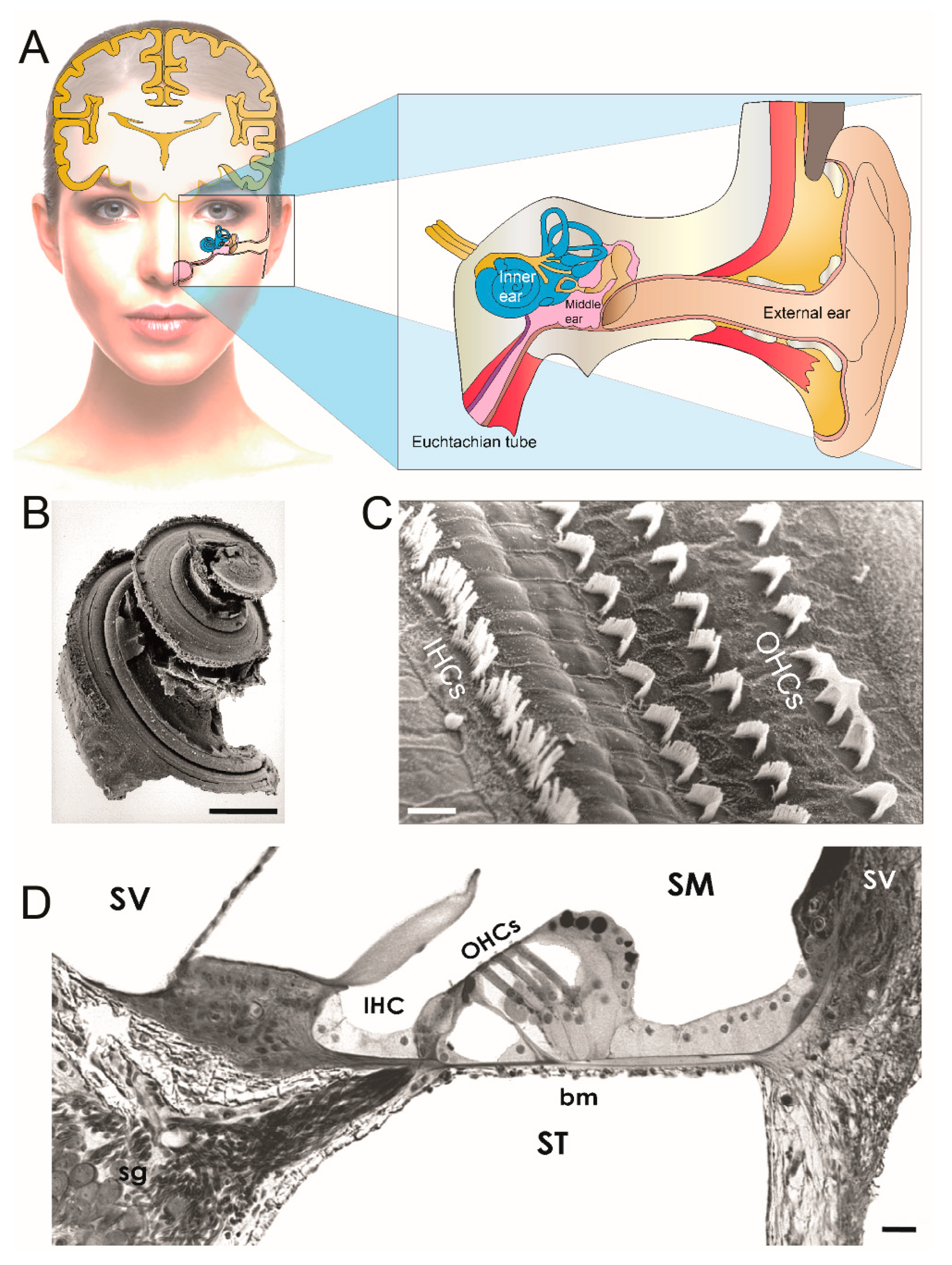
:1. Introduction
2. Major Causal Factors of Age-Related Hearing Loss
2.1. Biological Aging on Hearing
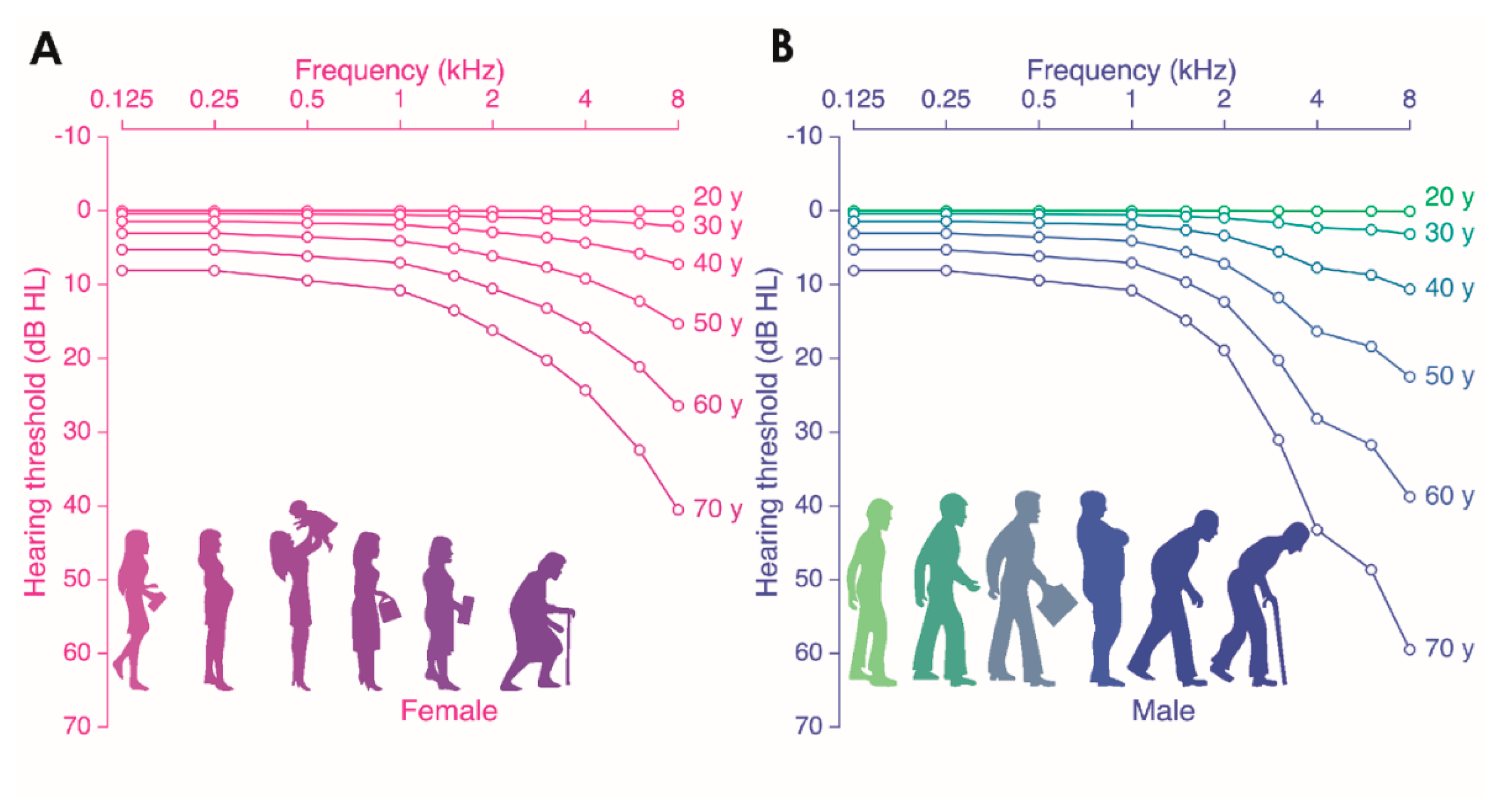
2.1.1. Aging and Hearing in Healthy People
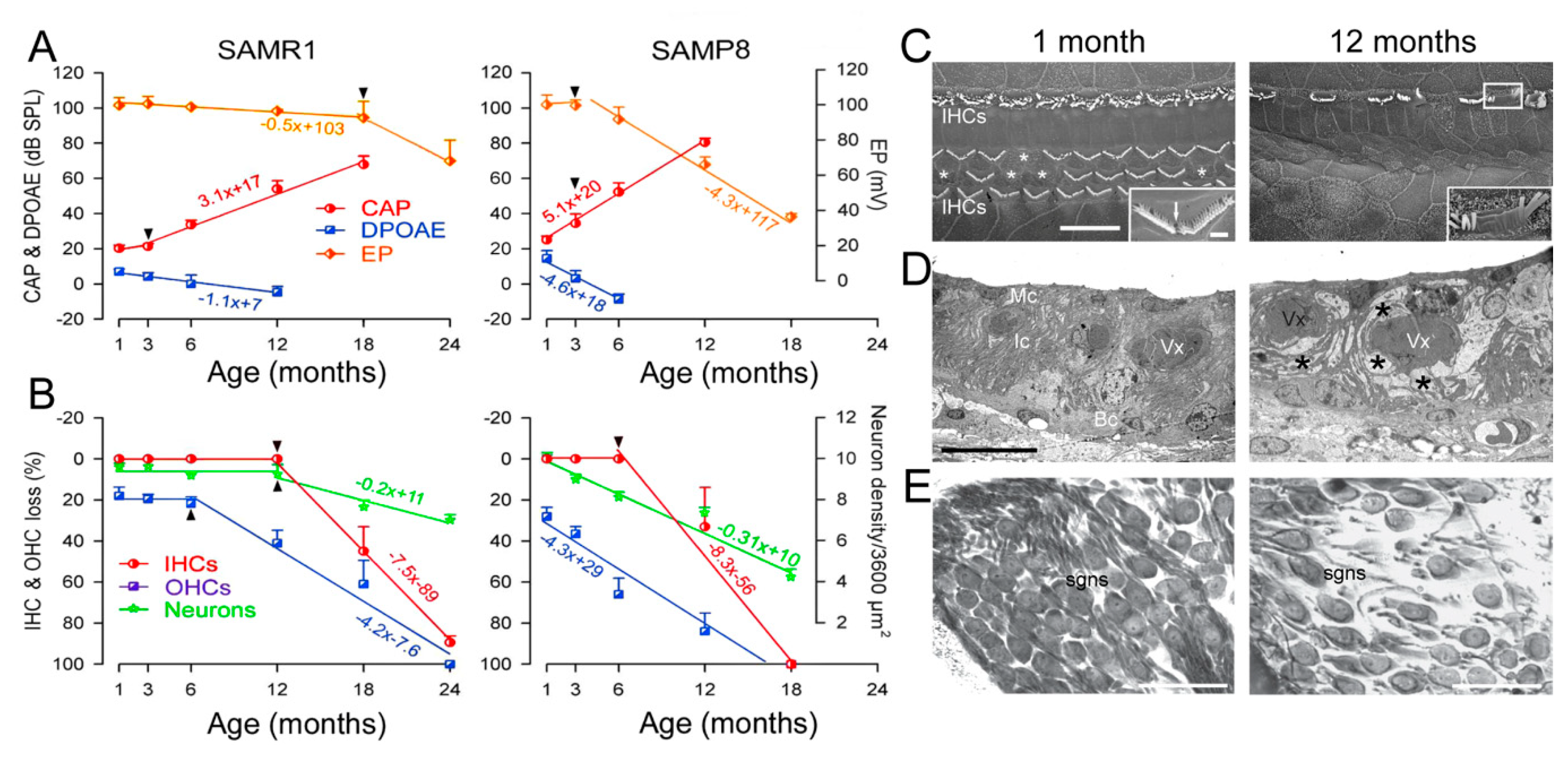
2.1.2. Aging and Hearing in Animals
2.2. Genetic Predispositions
2.3. Epigenetic Factors
2.3.1. DNA Methylation
2.3.2. Histone Modification
2.4. Environmental Factors
2.4.1. Noise Exposure
2.4.2. Ototoxic Medications
3. Molecular Mechanisms of Presbycusis
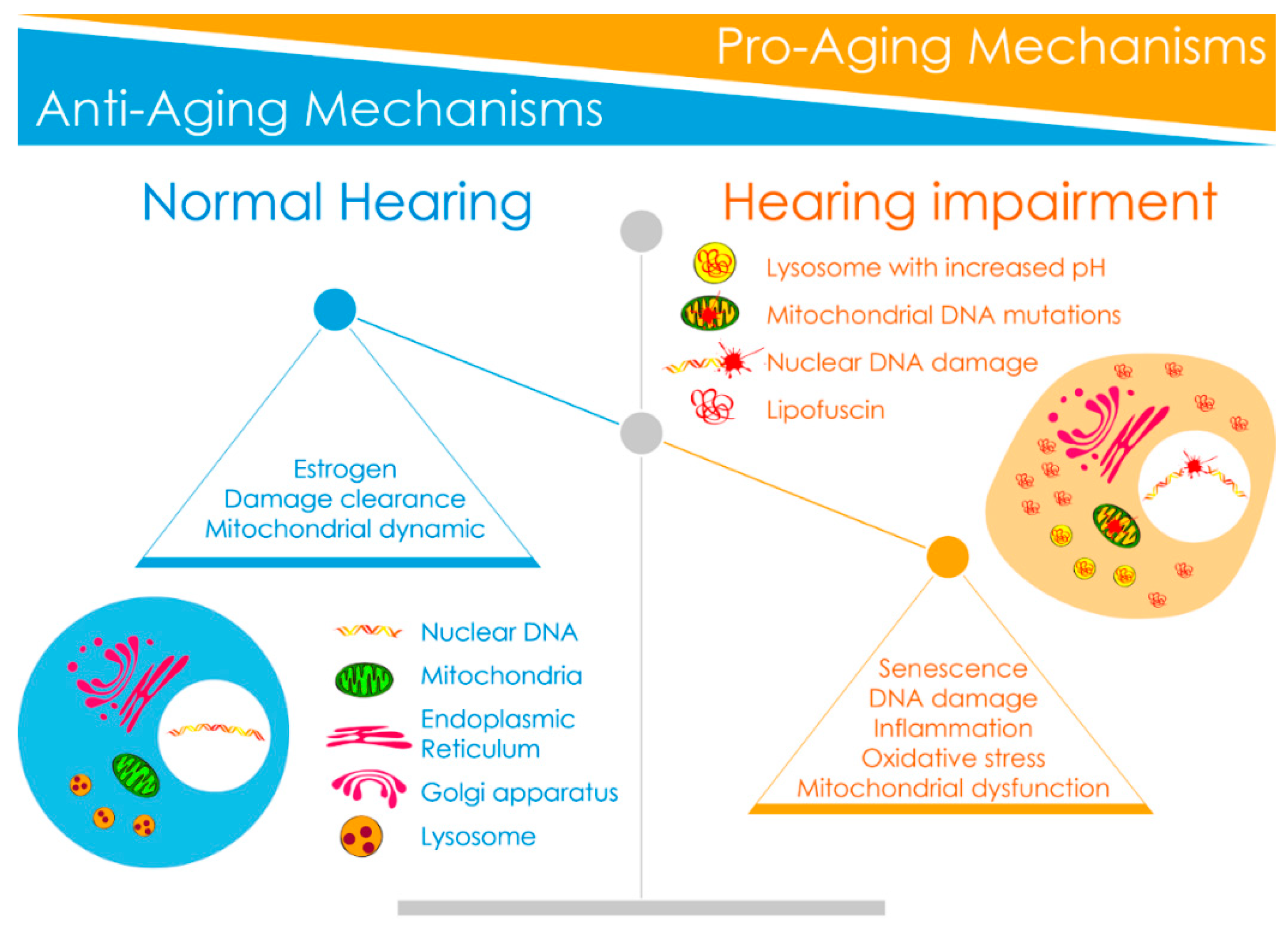
3.1. Pro-Aging Mechanisms Identified in the Cochlea
3.1.1. Oxidative Stress
3.1.2. DNA Damage and DNA Damage Responses
3.1.3. Mitochondrial DNA Mutations and Dysfunction
3.1.4. Impaired Mitochondrial Biogenesis
3.1.5. Senescence-Like Phenotype
3.1.6. Pro-Inflammatory Cytokines
3.1.7. Apoptosis
3.2. Anti-Aging Mechanisms
3.2.1. Mitochondrial Quality Control and Autophagy
3.2.2. Estrogen
4. Pharmacotherapies
4.1. Antioxidants, Free Radical Scavengers, and Anti-Inflammatories
4.2. Regulators of Mitochondrial Function and Metabolism
4.3. Caspase Inhibitors
4.4. Neurotrophins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cochlear Mechanisms of Presbycusis | Ref | Pre-Clinical and Clinical Trials (Tested Molecules) | Ref |
|---|---|---|---|
Oxidative stress & DNA damage
| [57,58,59,60,61,62,63,64,65,66,67,68,69,70] | Pre-clinical in animals
| [62,70,107,109,110,111,112] |
| Mitochondrial DNA mutation & dysfunction | [71,72,73,74,75,76,77,78,79,80,81,82] | Pre-clinical in animals SIRT1 activator resveratrol | [97] |
| Senescence-like phenotype & Pro-inflammatory cytokines (IL-1β and TNF) | [35,59,60,70,80,83,84,85,86,87,88,89] | Clinical trial Aspirin | [114] |
| Apoptosis Ratio pro-apoptotic /anti-apoptotic gene | [51,62,90,91,92] | Pre-clinical in animals Pan-caspase-inhibitor z-VAD-FMK Z-VAD-FMK. | [120,121] |
5. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front. Med. 2018, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Addressing the Rising Prevalence of Hearing Loss; World Health Organization: Geneva, Switzerland, 2018.
- Woodcock, K.; Pole, J.D. Educational attainment, labour force status and injury: A comparison of Canadians with and without deafness and hearing loss. Int. J. Rehabil. Res. 2008, 31, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Moscicki, E.K.; Elkins, E.F.; Baum, H.M.; McNamara, P.M. Hearing loss in the elderly: An epidemiologic study of the Framingham Heart Study Cohort. Ear Hear. 1985, 6, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Cruickshanks, K.J.; Wiley, T.L.; Tweed, T.S.; Klein, B.E.; Klein, R.; Mares-Perlman, J.A.; Nondahl, D.M. Prevalence of hearing loss in older adults in Beaver Dam, Wisconsin. The Epidemiology of Hearing Loss Study. Am. J. Epidemiol. 1998, 148, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Schuknecht, H.F.; Gacek, M.R. Cochlear pathology in presbycusis. Ann. Otol. Rhinol. Laryngol. 1993, 102, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; Gagnon, P.M. Apical-to-basal gradients in age-related cochlear degeneration and their relationship to “primary” loss of cochlear neurons. J. Comp. Neurol. 2004, 479, 103–116. [Google Scholar] [CrossRef]
- Fernandez, K.A.; Jeffers, P.W.; Lall, K.; Liberman, M.C.; Kujawa, S.G. Aging after noise exposure: Acceleration of cochlear synaptopathy in “recovered” ears. J. Neurosci. 2015, 35, 7509–7520. [Google Scholar] [CrossRef] [Green Version]
- Kujawa, S.G.; Liberman, M.C. Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hear. Res. 2015, 330, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Liberman, M.C.; Epstein, M.J.; Cleveland, S.S.; Wang, H.; Maison, S.F. Toward a Differential Diagnosis of Hidden Hearing Loss in Humans. PLoS ONE 2016, 11, e0162726. [Google Scholar] [CrossRef]
- Ralli, M.; Greco, A.; De Vincentiis, M.; Sheppard, A.; Cappelli, G.; Neri, I.; Salvi, R. Tone-in-noise detection deficits in elderly patients with clinically normal hearing. Am. J. Otolaryngol. 2019, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gates, G.A.; Couropmitree, N.N.; Myers, R.H. Genetic associations in age-related hearing thresholds. Arch. Otolaryngol. Head Neck Surg. 1999, 125, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Guest, M.; Boggess, M.; Attia, J.; SHOAMP study team and Scientific Advisory Committee. Relative risk of elevated hearing threshold compared to ISO1999 normative populations for Royal Australian Air Force male personnel. Hear. Res. 2012, 285, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Rosen, S.; Bergman, M.; Plester, D.; El-Mofty, A.; Satti, M.H. Presbycusis study of a relatively noise-free population in the Sudan. Ann. Otol. Rhinol. Laryngol. 1962, 71, 727–743. [Google Scholar] [CrossRef]
- Bergman, M. Hearing in the Mabaans. A critical review of related literature. Arch. Otolaryngol. 1966, 84, 411–415. [Google Scholar] [CrossRef]
- Goycoolea, M.V.; Goycoolea, H.G.; Farfan, C.R.; Rodriguez, L.G.; Martinez, G.C.; Vidal, R. Effect of life in industrialized societies on hearing in natives of Easter Island. Laryngoscope 1986, 96, 1391–1396. [Google Scholar] [CrossRef]
- Bielefeld, E.C.; Tanaka, C.; Chen, G.D.; Henderson, D. Age-related hearing loss: Is it a preventable condition? Hear. Res. 2010, 264, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Ciorba, A.; Benatti, A.; Bianchini, C.; Aimoni, C.; Volpato, S.; Bovo, R.; Martini, A. High frequency hearing loss in the elderly: Effect of age and noise exposure in an Italian group. J. Laryngol. Otol. 2011, 125, 776–780. [Google Scholar] [CrossRef]
- Schmiedt, R.A.; Mills, J.H.; Adams, J.C. Tuning and suppression in auditory nerve fibers of aged gerbils raised in quiet or noise. Hear. Res. 1990, 45, 221–236. [Google Scholar] [CrossRef]
- Schuknecht, H.F.; Watanuki, K.; Takahashi, T.; Belal, A.A., Jr.; Kimura, R.S.; Jones, D.D.; Ota, C.Y. Atrophy of the stria vascularis, a common cause for hearing loss. Laryngoscope 1974, 84, 1777–1821. [Google Scholar] [CrossRef]
- Gratton, M.A.; Schmiedt, R.A.; Schulte, B.A. Age-related decreases in endocochlear potential are associated with vascular abnormalities in the stria vascularis. Hear. Res. 1996, 102, 181–190. [Google Scholar] [CrossRef]
- Gratton, M.A.; Smyth, B.J.; Lam, C.F.; Boettcher, F.A.; Schmiedt, R.A. Decline in the endocochlear potential corresponds to decreased Na,K-ATPase activity in the lateral wall of quiet-aged gerbils. Hear. Res. 1997, 108, 9–16. [Google Scholar] [CrossRef]
- Hellstrom, L.I.; Schmiedt, R.A. Compound action potential input/output functions in young and quiet-aged gerbils. Hear. Res. 1990, 50, 163–174. [Google Scholar] [CrossRef]
- Schulte, B.A.; Schmiedt, R.A. Lateral wall Na,K-ATPase and endocochlear potentials decline with age in quiet-reared gerbils. Hear. Res. 1992, 61, 35–46. [Google Scholar] [CrossRef]
- Gratton, M.A.; Schulte, B.A. Alterations in microvasculature are associated with atrophy of the stria vascularis in quiet-aged gerbils. Hear. Res. 1995, 82, 44–52. [Google Scholar] [CrossRef]
- Christensen, K.; Frederiksen, H.; Hoffman, H.J. Genetic and environmental influences on self-reported reduced hearing in the old and oldest old. J. Am. Geriatr. Soc. 2001, 49, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Viljanen, A.; Era, P.; Kaprio, J.; Pyykko, I.; Koskenvuo, M.; Rantanen, T. Genetic and environmental influences on hearing in older women. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2007, 62, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Unal, M.; Tamer, L.; Dogruer, Z.N.; Yildirim, H.; Vayisoglu, Y.; Camdeviren, H. N-acetyltransferase 2 gene polymorphism and presbycusis. Laryngoscope 2005, 115, 2238–2241. [Google Scholar] [CrossRef]
- Van Eyken, E.; Van Camp, G.; Fransen, E.; Topsakal, V.; Hendrickx, J.J.; Demeester, K.; Van de Heyning, P.; Maki-Torkko, E.; Hannula, S.; Sorri, M.; et al. Contribution of the N-acetyltransferase 2 polymorphism NAT2*6A to age-related hearing impairment. J. Med. Genet. 2007, 44, 570–578. [Google Scholar] [CrossRef]
- Bared, A.; Ouyang, X.; Angeli, S.; Du, L.L.; Hoang, K.; Yan, D.; Liu, X.Z. Antioxidant enzymes, presbycusis, and ethnic variability. Otolaryngol. Head Neck Surg. 2010, 143, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, L.S.; Cadge, B.A.; Gomez-Dorado, M.; Dawson, S.J. A functional and genetic analysis of SOD2 promoter variants and their contribution to age-related hearing loss. Mech. Ageing Dev. 2013, 134, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Arsenijevic, D.; Onuma, H.; Pecqueur, C.; Raimbault, S.; Manning, B.S.; Miroux, B.; Couplan, E.; Alves-Guerra, M.C.; Goubern, M.; Surwit, R.; et al. Disruption of the uncoupling protein-2 gene in mice reveals a role in immunity and reactive oxygen species production. Nat. Genet. 2000, 26, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, S.; Uchida, Y.; Nakashima, T.; Ando, F.; Shimokata, H. The association between gene polymorphisms in uncoupling proteins and hearing impairment in Japanese elderly. Acta Oto-Laryngol. 2010, 130, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Sugiura, S.; Sone, M.; Ueda, H.; Nakashima, T. Progress and prospects in human genetic research into age-related hearing impairment. BioMed Res. Int. 2014, 2014, 390601. [Google Scholar] [CrossRef] [Green Version]
- Van Laer, L.; Van Eyken, E.; Fransen, E.; Huyghe, J.R.; Topsakal, V.; Hendrickx, J.J.; Hannula, S.; Maki-Torkko, E.; Jensen, M.; Demeester, K.; et al. The grainyhead like 2 gene (GRHL2), alias TFCP2L3, is associated with age-related hearing impairment. Hum. Mol. Genet. 2008, 17, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Van Eyken, E.; Van Laer, L.; Fransen, E.; Topsakal, V.; Lemkens, N.; Laureys, W.; Nelissen, N.; Vandevelde, A.; Wienker, T.; Van De Heyning, P.; et al. KCNQ4: A gene for age-related hearing impairment? Hum. Mutat. 2006, 27, 1007–1016. [Google Scholar] [CrossRef]
- Friedman, R.A.; Van Laer, L.; Huentelman, M.J.; Sheth, S.S.; Van Eyken, E.; Corneveaux, J.J.; Tembe, W.D.; Halperin, R.F.; Thorburn, A.Q.; Thys, S.; et al. GRM7 variants confer susceptibility to age-related hearing impairment. Hum. Mol. Genet. 2009, 18, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.L.; Fisher, L.M.; Ohmen, J.; Parody, R.; Fong, C.T.; Frisina, S.T.; Mapes, F.; Eddins, D.A.; Robert Frisina, D.; Frisina, R.D.; et al. GRM7 variants associated with age-related hearing loss based on auditory perception. Hear. Res. 2012, 294, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.R.; Zheng, Q.Y. Ahl2, a second locus affecting age-related hearing loss in mice. Genomics 2002, 80, 461–464. [Google Scholar] [CrossRef]
- Morita, Y.; Hirokawa, S.; Kikkawa, Y.; Nomura, T.; Yonekawa, H.; Shiroishi, T.; Takahashi, S.; Kominami, R. Fine mapping of Ahl3 affecting both age-related and noise-induced hearing loss. Biochem. Biophys. Res. Commun. 2007, 355, 117–121. [Google Scholar] [CrossRef]
- Vaiserman, A.; Lushchak, O. Developmental origins of type 2 diabetes: Focus on epigenetics. Ageing Res. Rev. 2019, 55, 100957. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, M.J.; Domann, F.E. A role for epigenetics in hearing: Establishment and maintenance of auditory specific gene expression patterns. Hear. Res. 2007, 233, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, F.H.; Kong, Q.P.; Perry, B.; He, Y.H. Progress on the role of DNA methylation in aging and longevity. Brief. Funct. Genom. 2016, 15, 454–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wang, Y.; Sun, Y.; Chen, S.; Zhang, S.; Shen, L.; Huang, X.; Lin, X.; Kong, W. Reduced expression of Connexin26 and its DNA promoter hypermethylation in the inner ear of mimetic aging rats induced by d-galactose. Biochem. Biophys. Res. Commun. 2014, 452, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zheng, J.; Shen, W.; Ma, L.; Zhao, M.; Wang, X.; Tang, J.; Yan, J.; Wu, Z.; Zou, Z.; et al. Elevated SLC26A4 gene promoter methylation is associated with the risk of presbycusis in men. Mol. Med. Rep. 2017, 16, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, A.; Smeti, I.; Dhouib, L.; Roche, M.; Achour, I.; Khalfallah, A.; Gibriel, A.A.; Charfeddine, I.; Ayadi, H.; Lachuer, J.; et al. Down-expression of P2RX2, KCNQ5, ERBB3 and SOCS3 through DNA hypermethylation in elderly women with presbycusis. Biomarkers 2018, 23, 347–356. [Google Scholar] [CrossRef]
- Watanabe, K.; Bloch, W. Histone methylation and acetylation indicates epigenetic change in the aged cochlea of mice. Eur. Arch. Oto-Rhino-Laryngol. 2013, 270, 1823–1830. [Google Scholar] [CrossRef]
- Fransen, E.; Lemkens, N.; Van Laer, L.; Van Camp, G. Age-related hearing impairment (ARHI): Environmental risk factors and genetic prospects. Exp. Gerontol. 2003, 38, 353–359. [Google Scholar] [CrossRef]
- Gates, G.A.; Schmid, P.; Kujawa, S.G.; Nam, B.; D’Agostino, R. Longitudinal threshold changes in older men with audiometric notches. Hear. Res. 2000, 141, 220–228. [Google Scholar] [CrossRef]
- Kujawa, S.G.; Liberman, M.C. Acceleration of age-related hearing loss by early noise exposure: Evidence of a misspent youth. J. Neurosci. 2006, 26, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Fetoni, A.R.; Picciotti, P.M.; Paludetti, G.; Troiani, D. Pathogenesis of presbycusis in animal models: A review. Exp. Gerontol. 2011, 46, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.C.; Fuentes-Santamaria, V.; Gabaldon-Ull, M.C.; Juiz, J.M. Age-Related Hearing Loss Is Accelerated by Repeated Short-Duration Loud Sound Stimulation. Front. Neurosci. 2019, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.C.; Fuentes-Santamaria, V.; Gabaldon-Ull, M.C.; Blanco, J.L.; Juiz, J.M. Wistar rats: A forgotten model of age-related hearing loss. Front. Aging Neurosci. 2014, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, Y.; Cruickshanks, K.J.; Klein, B.E.K.; Klein, R.; Hong, O.; Wallhagen, M. The Contribution of Ototoxic Medications to Hearing Loss among Older Adults. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Someya, S. Maintaining good hearing: Calorie restriction, Sirt3, and glutathione. Exp. Gerontol. 2013, 48, 1091–1095. [Google Scholar] [CrossRef] [Green Version]
- Menardo, J.; Tang, Y.; Ladrech, S.; Lenoir, M.; Casas, F.; Michel, C.; Bourien, J.; Ruel, J.; Rebillard, G.; Maurice, T.; et al. Oxidative stress, inflammation, and autophagic stress as the key mechanisms of premature age-related hearing loss in SAMP8 mouse Cochlea. Antioxid. Redox Signal. 2012, 16, 263–274. [Google Scholar] [CrossRef]
- Lasisi, A.O.; Fehintola, F.A. Correlation between plasma levels of radical scavengers and hearing threshold among elderly subjects with age-related hearing loss. Acta Oto-Laryngol. 2011, 131, 1160–1164. [Google Scholar] [CrossRef]
- Jiang, H.; Talaska, A.E.; Schacht, J.; Sha, S.H. Oxidative imbalance in the aging inner ear. Neurobiol. Aging 2007, 28, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Someya, S.; Xu, J.; Kondo, K.; Ding, D.; Salvi, R.J.; Yamasoba, T.; Rabinovitch, P.S.; Weindruch, R.; Leeuwenburgh, C.; Tanokura, M.; et al. Age-related hearing loss in C57BL/6J mice is mediated by Bak-dependent mitochondrial apoptosis. Proc. Natl. Acad. Sci. USA 2009, 106, 19432–19437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keithley, E.M.; Canto, C.; Zheng, Q.Y.; Wang, X.; Fischel-Ghodsian, N.; Johnson, K.R. Cu/Zn superoxide dismutase and age-related hearing loss. Hear. Res. 2005, 209, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Y.L.; Balaban, C.D. Regional distribution of manganese superoxide dismutase 2 (Mn SOD2) expression in rodent and primate spiral ganglion cells. Hear. Res. 2009, 253, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Mantha, A.K.; Sarkar, B.; Tell, G. A short review on the implications of base excision repair pathway for neurons: Relevance to neurodegenerative diseases. Mitochondrion 2014, 16, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Breen, A.P.; Murphy, J.A. Reactions of oxyl radicals with DNA. Free Radic. Biol. Med. 1995, 18, 1033–1077. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Benkafadar, N.; Menardo, J.; Bourien, J.; Nouvian, R.; Francois, F.; Decaudin, D.; Maiorano, D.; Puel, J.L.; Wang, J. Reversible p53 inhibition prevents cisplatin ototoxicity without blocking chemotherapeutic efficacy. EMBO Mol. Med. 2017, 9, 7–26. [Google Scholar] [CrossRef]
- Kamogashira, T.; Hayashi, K.; Fujimoto, C.; Iwasaki, S.; Yamasoba, T. Functionally and morphologically damaged mitochondria observed in auditory cells under senescence-inducing stress. NPJ Aging Mech. Dis. 2017, 3, 2. [Google Scholar] [CrossRef]
- Benkafadar, N.; Francois, F.; Affortit, C.; Casas, F.; Ceccato, J.C.; Menardo, J.; Venail, F.; Malfroy-Camine, B.; Puel, J.L.; Wang, J. ROS-Induced Activation of DNA Damage Responses Drives Senescence-Like State in Postmitotic Cochlear Cells: Implication for Hearing Preservation. Mol. Neurobiol. 2019, 56, 5950–5969. [Google Scholar] [CrossRef] [Green Version]
- Trifunovic, A.; Wredenberg, A.; Falkenberg, M.; Spelbrink, J.N.; Rovio, A.T.; Bruder, C.E.; Bohlooly, Y.M.; Gidlof, S.; Oldfors, A.; Wibom, R.; et al. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature 2004, 429, 417–423. [Google Scholar] [CrossRef]
- Chen, H.; Tang, J. The role of mitochondria in age-related hearing loss. Biogerontology 2014, 15, 13–19. [Google Scholar] [CrossRef]
- Markaryan, A.; Nelson, E.G.; Hinojosa, R. Quantification of the mitochondrial DNA common deletion in presbycusis. Laryngoscope 2009, 119, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Bai, U.; Seidman, M.D.; Hinojosa, R.; Quirk, W.S. Mitochondrial DNA deletions associated with aging and possibly presbycusis: A human archival temporal bone study. Am. J. Otol. 1997, 18, 449–453. [Google Scholar] [PubMed]
- Fischel-Ghodsian, N.; Bykhovskaya, Y.; Taylor, K.; Kahen, T.; Cantor, R.; Ehrenman, K.; Smith, R.; Keithley, E. Temporal bone analysis of patients with presbycusis reveals high frequency of mitochondrial mutations. Hear. Res. 1997, 110, 147–154. [Google Scholar] [CrossRef]
- Markaryan, A.; Nelson, E.G.; Hinojosa, R. Major arc mitochondrial DNA deletions in cytochrome c oxidase-deficient human cochlear spiral ganglion cells. Acta Oto-Laryngol. 2010, 130, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Filosto, M.; Mancuso, M.; Nishigaki, Y.; Pancrudo, J.; Harati, Y.; Gooch, C.; Mankodi, A.; Bayne, L.; Bonilla, E.; Shanske, S.; et al. Clinical and genetic heterogeneity in progressive external ophthalmoplegia due to mutations in polymerase gamma. Arch. Neurol. 2003, 60, 1279–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarzi, E.; Angebault, C.; Seveno, M.; Gueguen, N.; Chaix, B.; Bielicki, G.; Boddaert, N.; Mausset-Bonnefont, A.L.; Cazevieille, C.; Rigau, V.; et al. The human OPA1delTTAG mutation induces premature age-related systemic neurodegeneration in mouse. Brain 2012, 135, 3599–3613. [Google Scholar] [CrossRef]
- Trifunovic, A.; Hansson, A.; Wredenberg, A.; Rovio, A.T.; Dufour, E.; Khvorostov, I.; Spelbrink, J.N.; Wibom, R.; Jacobs, H.T.; Larsson, N.G. Somatic mtDNA mutations cause aging phenotypes without affecting reactive oxygen species production. Proc. Natl. Acad. Sci. USA 2005, 102, 17993–17998. [Google Scholar] [CrossRef] [Green Version]
- Kujoth, G.C.; Hiona, A.; Pugh, T.D.; Someya, S.; Panzer, K.; Wohlgemuth, S.E.; Hofer, T.; Seo, A.Y.; Sullivan, R.; Jobling, W.A.; et al. Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science 2005, 309, 481–484. [Google Scholar] [CrossRef]
- Sanchis-Gomar, F.; Garcia-Gimenez, J.L.; Gomez-Cabrera, M.C.; Pallardo, F.V. Mitochondrial biogenesis in health and disease. Molecular and therapeutic approaches. Curr. Pharm. Des. 2014, 20, 5619–5633. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Sun, J.L.; Hu, Y.J.; Yang, Y.; Zhang, W.J.; Hu, Y.; Li, J.; Sun, Y.; Zhong, Y.; Peng, W.; et al. The effect of overexpression of PGC-1alpha on the mtDNA4834 common deletion in a rat cochlear marginal cell senescence model. Hear. Res. 2013, 296, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Herranz, N.; Gil, J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [Green Version]
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Sapieha, P.; Mallette, F.A. Cellular Senescence in Postmitotic Cells: Beyond Growth Arrest. Trends Cell Biol. 2018, 28, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Howcroft, T.K.; Campisi, J.; Louis, G.B.; Smith, M.T.; Wise, B.; Wyss-Coray, T.; Augustine, A.D.; McElhaney, J.E.; Kohanski, R.; Sierra, F. The role of inflammation in age-related disease. Aging 2013, 5, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, S.D.; Cruickshanks, K.J.; Zhan, W.; Tsai, M.Y.; Klein, R.; Chappell, R.; Nieto, F.J.; Klein, B.E.; Schubert, C.R.; Dalton, D.S.; et al. Long-term assessment of systemic inflammation and the cumulative incidence of age-related hearing impairment in the epidemiology of hearing loss study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verschuur, C.A.; Dowell, A.; Syddall, H.E.; Ntani, G.; Simmonds, S.J.; Baylis, D.; Gale, C.R.; Walsh, B.; Cooper, C.; Lord, J.M.; et al. Markers of inflammatory status are associated with hearing threshold in older people: Findings from the Hertfordshire Ageing Study. Age Ageing 2012, 41, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Tadros, S.F.; D’Souza, M.; Zhu, X.; Frisina, R.D. Apoptosis-related genes change their expression with age and hearing loss in the mouse cochlea. Apoptosis Int. J. Program. Cell Death 2008, 13, 1303–1321. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, C.; Yamasoba, T. Oxidative stresses and mitochondrial dysfunction in age-related hearing loss. Oxid. Med. Cell. Longev. 2014, 2014, 582849. [Google Scholar] [CrossRef] [Green Version]
- Falah, M.; Houshmand, M.; Najafi, M.; Balali, M.; Mahmoudian, S.; Asghari, A.; Emamdjomeh, H.; Farhadi, M. The potential role for use of mitochondrial DNA copy number as predictive biomarker in presbycusis. Ther. Clin. Risk Manag. 2016, 12, 1573–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Puel, J.L. Toward Cochlear Therapies. Physiol. Rev. 2018, 98, 2477–2522. [Google Scholar] [CrossRef] [PubMed]
- Aburto, M.R.; Sanchez-Calderon, H.; Hurle, J.M.; Varela-Nieto, I.; Magarinos, M. Early otic development depends on autophagy for apoptotic cell clearance and neural differentiation. Cell Death Dis. 2012, 3, e394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Iriarte Rodriguez, R.; Pulido, S.; Rodriguez-de la Rosa, L.; Magarinos, M.; Varela-Nieto, I. Age-regulated function of autophagy in the mouse inner ear. Hear. Res. 2015, 330, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Landreh, M.; Cao, K.; Abe, M.; Hendriks, G.J.; Kennerdell, J.R.; Zhu, Y.; Wang, L.S.; Bonini, N.M. The microRNA miR-34 modulates ageing and neurodegeneration in Drosophila. Nature 2012, 482, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Xiong, H.; Pang, J.; Yang, H.; Dai, M.; Liu, Y.; Ou, Y.; Huang, Q.; Chen, S.; Zhang, Z.; Xu, Y.; et al. Activation of miR-34a/SIRT1/p53 signaling contributes to cochlear hair cell apoptosis: Implications for age-related hearing loss. Neurobiol. Aging 2015, 36, 1692–1701. [Google Scholar] [CrossRef]
- Pang, J.; Xiong, H.; Lin, P.; Lai, L.; Yang, H.; Liu, Y.; Huang, Q.; Chen, S.; Ye, Y.; Sun, Y.; et al. Activation of miR-34a impairs autophagic flux and promotes cochlear cell death via repressing ATG9A: Implications for age-related hearing loss. Cell Death Dis. 2017, 8, e3079. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Zarate, S.; Stevnsner, T.; Gredilla, R. Role of Estrogen and Other Sex Hormones in Brain Aging. Neuroprotection and DNA Repair. Front. Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, R.; Rosenhall, U.; Gause-Nilsson, I.; Steen, B. Auditory function in 70- and 75-year-olds of four age cohorts. A cross-sectional and time-lag study of presbyacusis. Scand. Audiol. 1998, 27, 81–93. [Google Scholar] [CrossRef]
- Stenberg, A.E.; Nylen, O.; Windh, M.; Hultcrantz, M. Otological problems in children with Turner’s syndrome. Hear. Res. 1998, 124, 85–90. [Google Scholar] [CrossRef]
- Hultcrantz, M.; Simonoska, R.; Stenberg, A.E. Estrogen and hearing: A summary of recent investigations. Acta Oto-Laryngol. 2006, 126, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Balogova, Z.; Popelar, J.; Chiumenti, F.; Chumak, T.; Burianova, J.S.; Rybalko, N.; Syka, J. Age-Related Differences in Hearing Function and Cochlear Morphology between Male and Female Fischer 344 Rats. Front. Aging Neurosci. 2017, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimaraes, P.; Zhu, X.; Cannon, T.; Kim, S.; Frisina, R.D. Sex differences in distortion product otoacoustic emissions as a function of age in CBA mice. Hear. Res. 2004, 192, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.R. Males lose hearing earlier in mouse models of late-onset age-related hearing loss; females lose hearing earlier in mouse models of early-onset hearing loss. Hear. Res. 2004, 190, 141–148. [Google Scholar] [CrossRef]
- Seidman, M.D.; Khan, M.J.; Bai, U.; Shirwany, N.; Quirk, W.S. Biologic activity of mitochondrial metabolites on aging and age-related hearing loss. Am. J. Otol. 2000, 21, 161–167. [Google Scholar] [CrossRef]
- Sha, S.H.; Kanicki, A.; Halsey, K.; Wearne, K.A.; Schacht, J. Antioxidant-enriched diet does not delay the progression of age-related hearing loss. Neurobiol. Aging 2012, 33, 1010.e15–1010.e16. [Google Scholar] [CrossRef] [Green Version]
- Seidman, M.D.; Khan, M.J.; Tang, W.X.; Quirk, W.S. Influence of lecithin on mitochondrial DNA and age-related hearing loss. Otolaryngol. Head Neck Surg. 2002, 127, 138–144. [Google Scholar] [CrossRef]
- Nevado, J.; Sanz, R.; Sanchez-Rodriguez, C.; Garcia-Berrocal, J.R.; Martin-Sanz, E.; Gonzalez-Garcia, J.A.; Esteban-Sanchez, J.; Ramirez-Camacho, R. Ginkgo biloba extract (EGb761) protects against aging-related caspase-mediated apoptosis in rat cochlea. Acta Oto-Laryngol. 2010, 130, 1101–1112. [Google Scholar] [CrossRef]
- Kang, J.W.; Choi, H.S.; Kim, K.; Choi, J.Y. Dietary vitamin intake correlates with hearing thresholds in the older population: The Korean National Health and Nutrition Examination Survey. Am. J. Clin. Nutr. 2014, 99, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Polanski, J.F.; Cruz, O.L. Evaluation of antioxidant treatment in presbyacusis: Prospective, placebo-controlled, double-blind, randomised trial. J. Laryngol. Otol. 2013, 127, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Di Matteo, V.; Benigno, A.; Pierucci, M.; Crescimanno, G.; Di Giovanni, G. Non-steroidal anti-inflammatory drugs in Parkinson’s disease. Exp. Neurol. 2007, 205, 295–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowthian, J.A.; Britt, C.J.; Rance, G.; Lin, F.R.; Woods, R.L.; Wolfe, R.; Nelson, M.R.; Dillon, H.A.; Ward, S.; Reid, C.M.; et al. Slowing the progression of age-related hearing loss: Rationale and study design of the ASPIRIN in HEARING, retinal vessels imaging and neurocognition in older generations (ASPREE-HEARING) trial. Contemp. Clin. Trials 2016, 46, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, H.Y.; Yu, M.; Yeom, E.; Lee, J.H.; Yoon, A.; Lee, K.S.; Min, K.J. Extension of Drosophila lifespan by Korean red ginseng through a mechanism dependent on dSir2 and insulin/IGF-1 signaling. Aging 2019, 11, 9369–9387. [Google Scholar] [CrossRef] [PubMed]
- Rascon, B.; Hubbard, B.P.; Sinclair, D.A.; Amdam, G.V. The lifespan extension effects of resveratrol are conserved in the honey bee and may be driven by a mechanism related to caloric restriction. Aging 2012, 4, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, B.; Baur, J.A. Resveratrol and life extension. Ann. N. Y. Acad. Sci. 2011, 1215, 138–143. [Google Scholar] [CrossRef]
- Han, C.; Linser, P.; Park, H.J.; Kim, M.J.; White, K.; Vann, J.M.; Ding, D.; Prolla, T.A.; Someya, S. Sirt1 deficiency protects cochlear cells and delays the early onset of age-related hearing loss in C57BL/6 mice. Neurobiol. Aging 2016, 43, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Someya, S.; Tanokura, M.; Weindruch, R.; Prolla, T.A.; Yamasoba, T. Effects of caloric restriction on age-related hearing loss in rodents and rhesus monkeys. Curr. Aging Sci. 2010, 3, 20–25. [Google Scholar] [CrossRef]
- Han, X.; Ge, R.; Xie, G.; Li, P.; Zhao, X.; Gao, L.; Zhang, H.; Wang, O.; Huang, F.; Han, F. Caspase-mediated apoptosis in the cochleae contributes to the early onset of hearing loss in A/J mice. ASN Neuro 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhang, H.; Han, X.; Zhao, X.; Hu, F.; Li, P.; Xie, G.; Gao, L.; Cheng, L.; Song, X.; et al. Attenuation of hearing loss in DBA/2J mice by anti-apoptotic treatment. Hear. Res. 2015, 327, 109–116. [Google Scholar] [CrossRef]
- Lee, H.; Shin, E.A.; Lee, J.H.; Ahn, D.; Kim, C.G.; Kim, J.H.; Kim, S.H. Caspase inhibitors: A review of recently patented compounds (2013–2015). Expert Opin. Ther. Pat. 2018, 28, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Viana, L.M.; O’Malley, J.T.; Burgess, B.J.; Jones, D.D.; Oliveira, C.A.; Santos, F.; Merchant, S.N.; Liberman, L.D.; Liberman, M.C. Cochlear neuropathy in human presbycusis: Confocal analysis of hidden hearing loss in post-mortem tissue. Hear. Res. 2015, 327, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makary, C.A.; Shin, J.; Kujawa, S.G.; Liberman, M.C.; Merchant, S.N. Age-related primary cochlear neuronal degeneration in human temporal bones. J. Assoc. Res. Otolaryngol. 2011, 12, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xing, Y.; Xia, L.; Chen, Z.; Yin, S.; Wang, J. AAV-mediated NT-3 overexpression protects cochleae against noise-induced synaptopathy. Gene Ther. 2018. [Google Scholar] [CrossRef] [Green Version]
- Pyykko, I.; Zou, J.; Schrott-Fischer, A.; Glueckert, R.; Kinnunen, P. An Overview of Nanoparticle Based Delivery for Treatment of Inner Ear Disorders. Methods Mol. Biol. 2016, 1427, 363–415. [Google Scholar] [CrossRef]
- Miller, R.A. Evaluating evidence for aging. Science 2005, 310, 441–443. [Google Scholar] [CrossRef]
- Ren, H.; Chen, J.; Wang, Y.; Zhang, S.; Zhang, B. Intracerebral neural stem cell transplantation improved the auditory of mice with presbycusis. Int. J. Clin. Exp. Pathol. 2013, 6, 230–241. [Google Scholar]
- Hu, Z.; Ulfendahl, M. Cell replacement therapy in the inner ear. Stem Cells Dev. 2006, 15, 449–459. [Google Scholar] [CrossRef]
- Corrales, C.E.; Pan, L.; Li, H.; Liberman, M.C.; Heller, S.; Edge, A.S. Engraftment and differentiation of embryonic stem cell-derived neural progenitor cells in the cochlear nerve trunk: Growth of processes into the organ of Corti. J. Neurobiol. 2006, 66, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jongkamonwiwat, N.; Abbas, L.; Eshtan, S.J.; Johnson, S.L.; Kuhn, S.; Milo, M.; Thurlow, J.K.; Andrews, P.W.; Marcotti, W.; et al. Restoration of auditory evoked responses by human ES-cell-derived otic progenitors. Nature 2012, 490, 278–282. [Google Scholar] [CrossRef]
- Davidsohn, N.; Pezzone, M.; Vernet, A.; Graveline, A.; Oliver, D.; Slomovic, S.; Punthambaker, S.; Sun, X.; Liao, R.; Bonventre, J.V.; et al. A single combination gene therapy treats multiple age-related diseases. Proc. Natl. Acad. Sci. USA 2019, 116, 23505–23511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantz, B.J.; Turner, C. Combining acoustic and electrical speech processing: Iowa/Nucleus hybrid implant. Acta Oto-Laryngol. 2004, 124, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Skarzynski, H.; Lorens, A.; Piotrowska, A.; Podskarbi-Fayette, R. Results of partial deafness cochlear implantation using various electrode designs. Audiol. Neuro-Otol. 45. [CrossRef]
- Liu, Y.; Jolly, C.; Braun, S.; Stark, T.; Scherer, E.; Plontke, S.K.; Kiefer, J. In vitro and in vivo pharmacokinetic study of a dexamethasone-releasing silicone for cochlear implants. Eur. Arch. Oto-Rhino-Laryngol. 2016, 273, 1745–1753. [Google Scholar] [CrossRef]
- Douchement, D.; Terranti, A.; Lamblin, J.; Salleron, J.; Siepmann, F.; Siepmann, J.; Vincent, C. Dexamethasone eluting electrodes for cochlear implantation: Effect on residual hearing. Cochlear Implant. Int. 2015, 16, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, L.; Simoni, E.; Giarbini, N.; Giordano, P.; Pannella, M.; Hatzopoulos, S.; Martini, A. Cochlear implant and inflammation reaction: Safety study of a new steroid-eluting electrode. Hear. Res. 2016, 336, 44–52. [Google Scholar] [CrossRef]
- Bas, E.; Bohorquez, J.; Goncalves, S.; Perez, E.; Dinh, C.T.; Garnham, C.; Hessler, R.; Eshraghi, A.A.; Van De Water, T.R. Electrode array-eluted dexamethasone protects against electrode insertion trauma induced hearing and hair cell losses, damage to neural elements, increases in impedance and fibrosis: A dose response study. Hear. Res. 2016, 337, 12–24. [Google Scholar] [CrossRef]
- Plontke, S.K.; Gotze, G.; Rahne, T.; Liebau, A. Intracochlear drug delivery in combination with cochlear implants : Current aspects. Hno 2016, 64, 797–807. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Puel, J.-L. Presbycusis: An Update on Cochlear Mechanisms and Therapies. J. Clin. Med. 2020, 9, 218. https://doi.org/10.3390/jcm9010218
Wang J, Puel J-L. Presbycusis: An Update on Cochlear Mechanisms and Therapies. Journal of Clinical Medicine. 2020; 9(1):218. https://doi.org/10.3390/jcm9010218
Chicago/Turabian StyleWang, Jing, and Jean-Luc Puel. 2020. "Presbycusis: An Update on Cochlear Mechanisms and Therapies" Journal of Clinical Medicine 9, no. 1: 218. https://doi.org/10.3390/jcm9010218
APA StyleWang, J., & Puel, J. -L. (2020). Presbycusis: An Update on Cochlear Mechanisms and Therapies. Journal of Clinical Medicine, 9(1), 218. https://doi.org/10.3390/jcm9010218