Immunomodulation by an Omega-6 Fatty Acid Reduced Mixed Lipid Emulsion in Murine Acute Respiratory Distress Syndrome



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals and Experimental Protocol
2.3. LPS-induced Acute Lung Injury in the Murine Model
2.4. Assessment of Lung Edema
2.5. BAL Leukocyte Counts
2.6. Enzyme-linked Immunosorbent Assay (ELISA)
2.7. Myeloperoxidase Assay
2.8. Assessment of Apoptosis by Flow Cytometry
2.9. Flow Cytometry and Staining Procedure
2.10. Statistics
3. Results
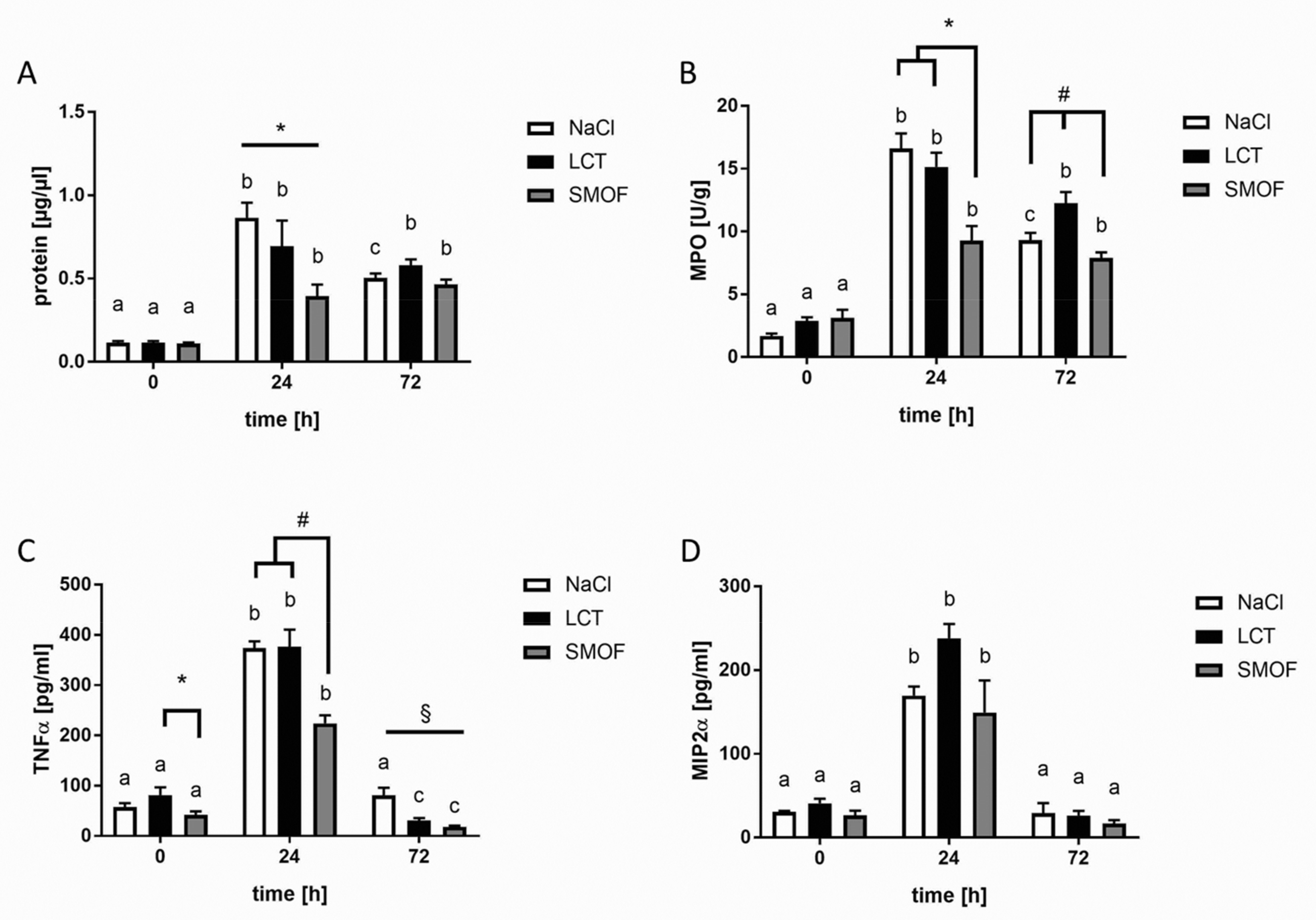
3.1. Protein Concentrations of BAL Fluids
3.2. Accumulation of Neutrophils in Lung Tissue
3.3. Generation of Cytokines in ARDS
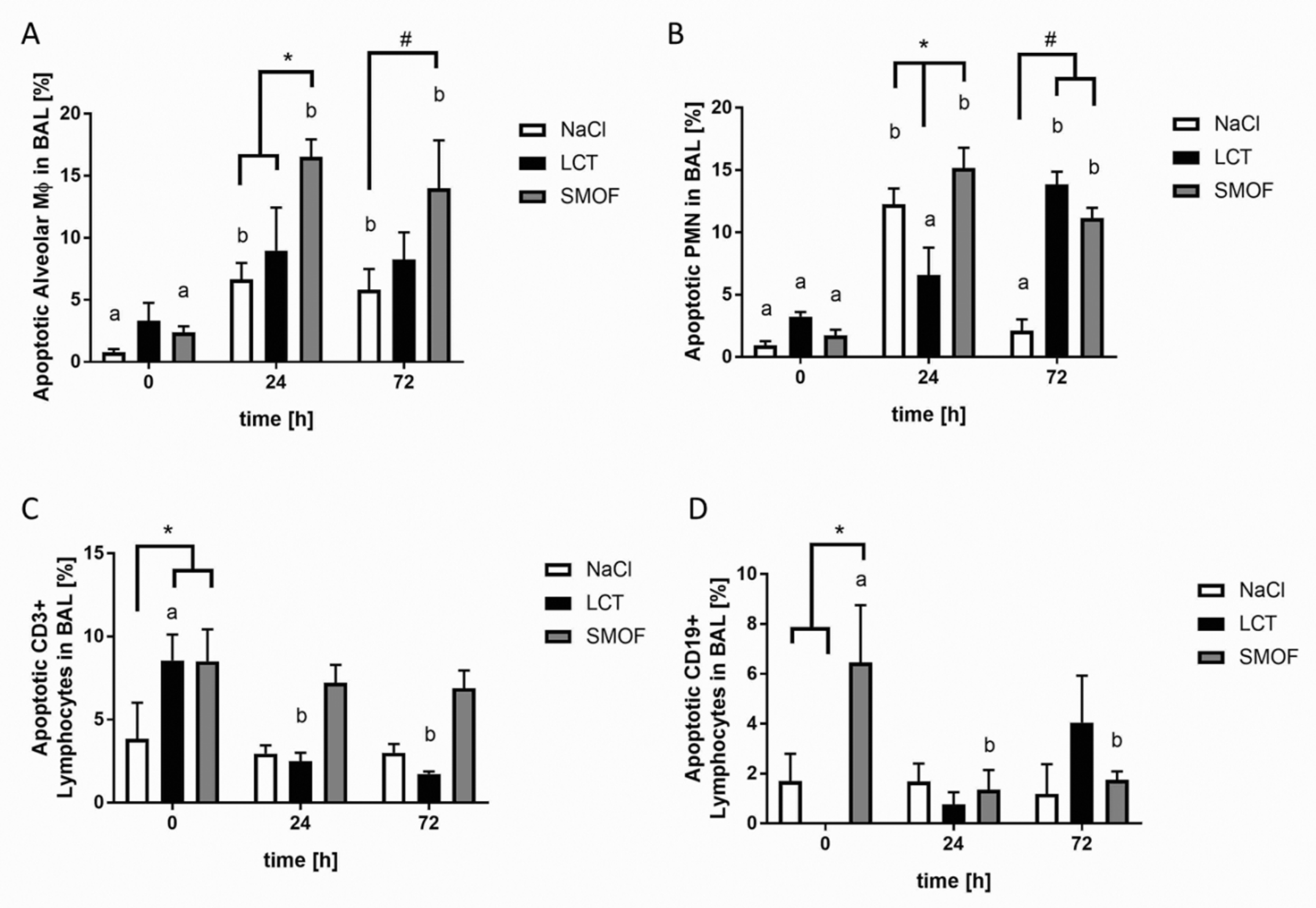
3.4. Effect of Lipid Emulsions on Alveolar Recruitment of Leukocytes in LPS-induced Acute Respiratory Distress Syndrome
3.5. Assessment of Apoptosis in LPS-induced Acute Respiratory Distress Syndrome
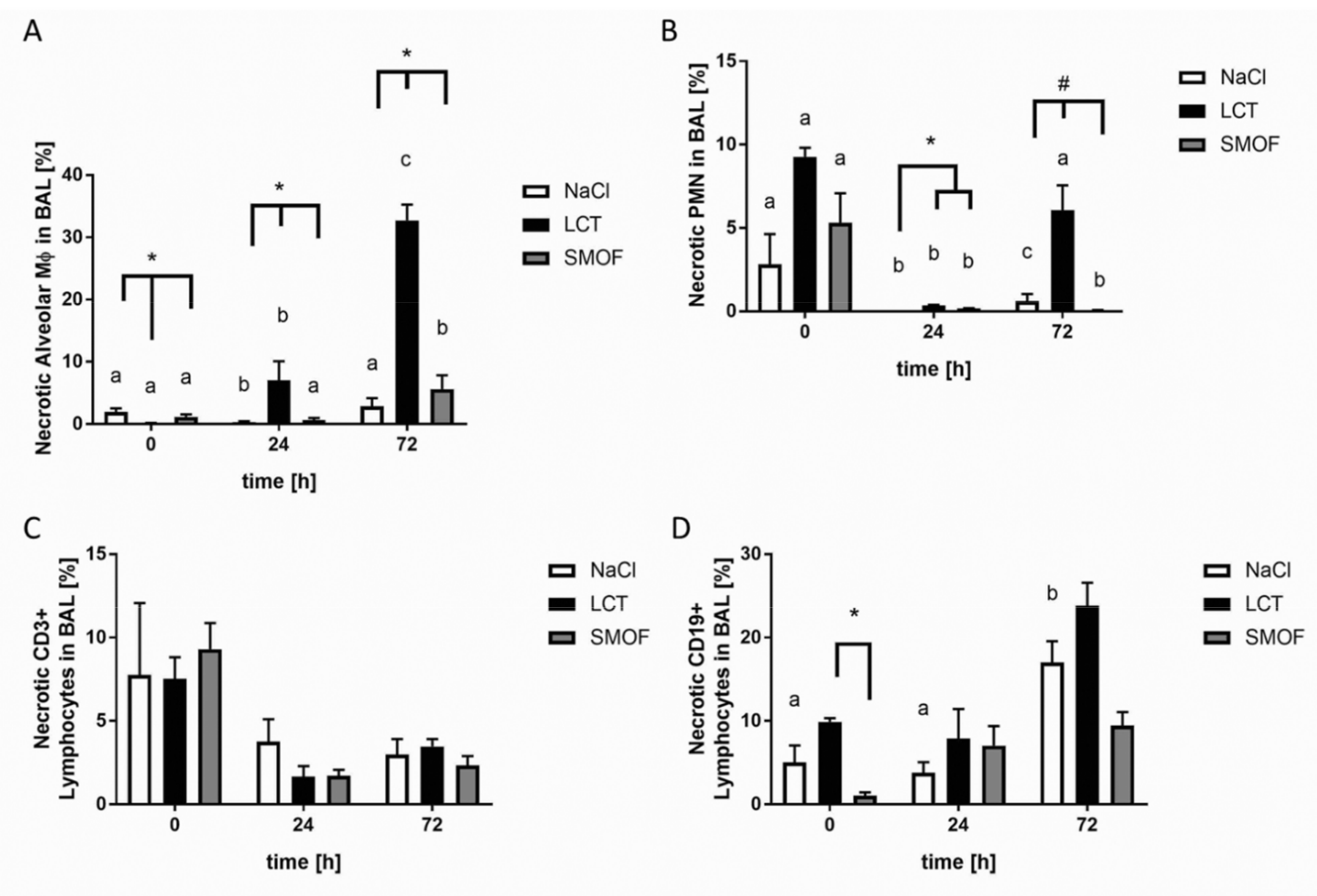
3.6. Analysis of Necrosis in LPS-induced Acute Respiratory Distress Syndrome
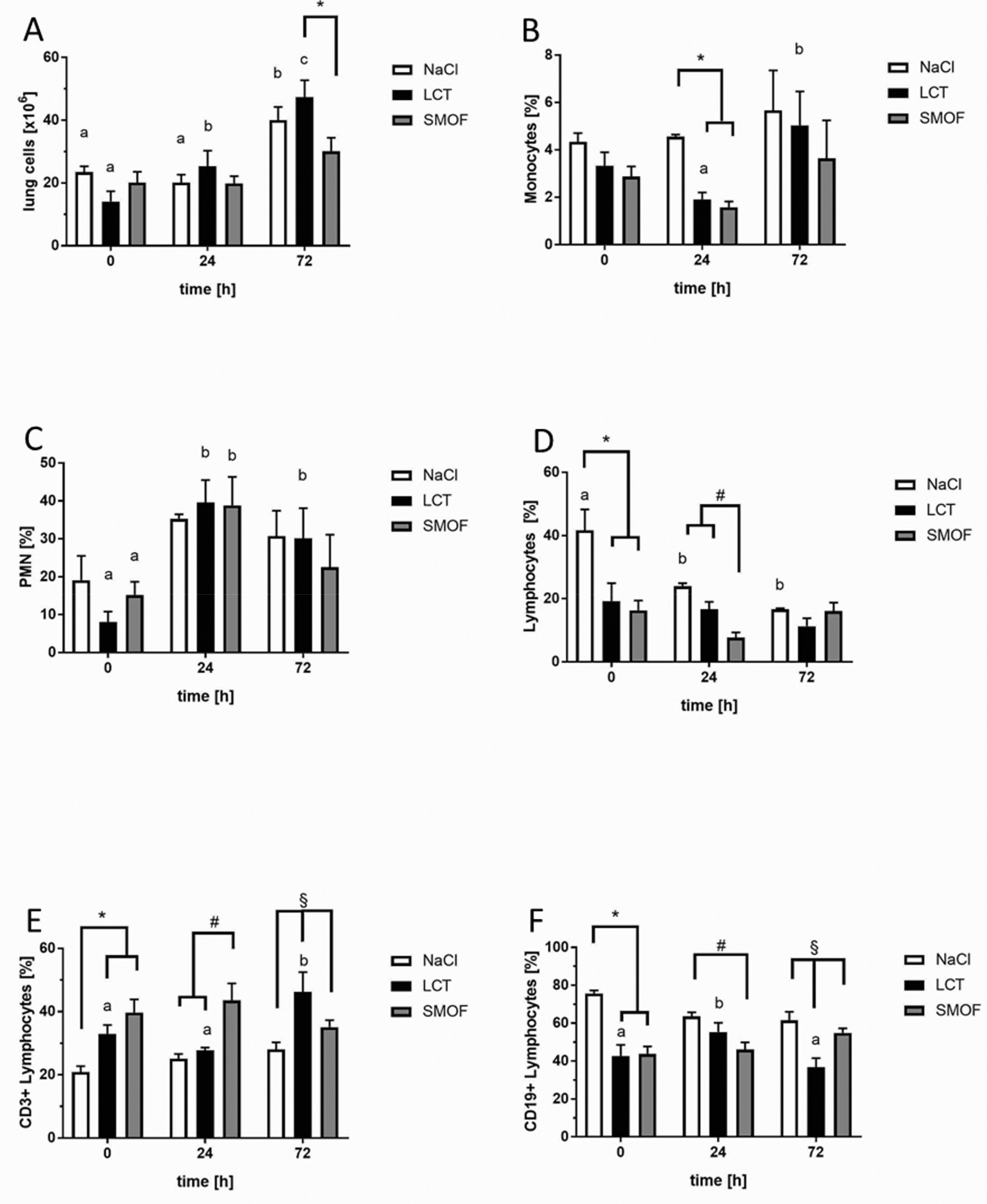
3.7. Effect of Lipid Emulsions on Lung-tissue Leukocytes in LPS-induced Acute Respiratory Distress Syndrome
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ware, L.; Matthay, M. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.M.A.; Ware, L.L.B.; Zimmerman, G.A. The acute respiratory distress syndrome. J. Clin. Investig. 2012, 122, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthay, M.A.; Zemans, R.L. The Acute Respiratory Distress Syndrome: Pathogenesis and Treatment. Annu. Rev. Pathol. 2010, 6, 147–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, M.; Walmrath, H.D.; Seeger, W.; Mayer, K. Clinical aspects of acute lung insufficiency (ALI/TRALI). Transfus. Med. Hemotherapy 2008, 35, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun-Buisson, C.; Minelli, C.; Bertolini, G.; Brazzi, L.; Pimentel, J.; Lewandowski, K.; Bion, J.; Romand, J.-A.; Villar, J.; Thorsteinsson, A.; et al. Epidemiology and outcome of acute lung injury in European intensive care units. Results from the ALIVE study. Intensive Care Med. 2004, 30, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, M.; Mayer, K. Intravenous lipids in adult intensive care unit patients. World Rev. Nutr. Diet. 2015, 112, 120–126. [Google Scholar]
- Hecker, M.; Felbinger, T.; Mayer, K. Nutrition in intensive care medicine. Anaesthesist 2012, 61, 553–561. [Google Scholar] [CrossRef]
- Boisramé-Helms, J.; Toti, F.; Hasselmann, M.; Meziani, F. Lipid emulsions for parenteral nutrition in critical illness. Prog. Lipid Res. 2015, 60, 1–16. [Google Scholar] [CrossRef]
- Mayer, K.; Seeger, W. Fish oil-containing lipid emulsions in patients with sepsis. Crit. Care 2010, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Heller, A.R.; Rössler, S.; Litz, R.J.; Steh, S.N.; Heller, S.C.; Koch, R.; Koch, T. Omega-3 fatty acids improve the diagnosis-related clinical outcome. Crit. Care Med. 2006, 34, 972–979. [Google Scholar] [CrossRef]
- Hecker, M.; Linder, T.; Ott, J.; Walmrath, H.-D.; Lohmeyer, J.; Vadász, I.; Marsh, L.M.; Herold, S.; Reichert, M.; Buchbinder, A.; et al. Immunomodulation by lipid emulsions in pulmonary inflammation: A randomized controlled trial. Crit. Care 2015, 19, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanten, G.J.; Calder, P.C. Immune modulation by parenteral lipid emulsions. Am. J. Clin. Nutr. 2007, 85, 1171–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cury-Boaventura, M.F.; Gorjão, R.; de Lima, T.M.; Fiamoncini, J.; Torres, R.P.; Mancini-Filho, J.; Soriano, F.G.; Curi, R. Effect of olive oil-based emulsion on human lymphocyte and neutrophil death. JPEN J. Parenter. Enteral Nutr. 2008, 32, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Cury-Boaventura, M.F.; Gorjão, R.; Martins de Lima, T.; Fiamoncini, J.; Godoy, A.B.P.; Deschamphs, F.C.; Soriano, F.G.; Curi, R. Effect of medium/ω-6 long chain triglyceride-based emulsion on leucocyte death and inflammatory gene expression. Clin. Exp. Immunol. 2011, 165, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Bi, M.H.; Ott, J.; Fischer, T.; Hecker, M.; Dietrich, H.; Schaefer, M.B.; Markart, P.; Wang, B.E.; Seeger, W.; Mayer, K. Induction of lymphocyte apoptosis in a murine model of acute lung injury-modulation by lipid emulsions. Shock 2010, 33, 179–188. [Google Scholar] [CrossRef]
- Mayer, K.; Schaefer, M.B.; Hecker, M. Intravenous n-3 fatty acids in the critically ill. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 124–128. [Google Scholar] [CrossRef]
- Jones, C.J.; Calder, P.C. Influence of different intravenous lipid emulsions on fatty acid status and laboratory and clinical outcomes in adult patients receiving home parenteral nutrition: A systematic review. Clin. Nutr. 2018, 37, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Hecker, M.; Ott, J.; Sondermann, C.; Schaefer, M.M.B.; Obert, M.; Hecker, A.; Morty, R.E.; Vadasz, I.; Herold, S.; Rosengarten, B.; et al. Immunomodulation by fish-oil containing lipid emulsions in murine acute respiratory distress syndrome. Crit. Care 2014, 18, R85. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosenbough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement With the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Singer, P.; Shapiro, H.; Theilla, M.; Anbar, R.; Singer, J.; Cohen, J. Anti-inflammatory properties of omega-3 fatty acids in critical illness: Novel mechanisms and an integrative perspective. Intensive Care Med. 2008, 34, 1580–1592. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M.B.; Wenzel, A.; Fischer, T.; Braun-Dullaeus, R.C.; Renner, F.; Dietrich, H.; Schaefer, C.A.; Seeger, W.; Mayer, K. Fatty acids differentially influence phosphatidylinositol 3-kinase signal transduction in endothelial cells: Impact on adhesion and apoptosis. Atherosclerosis 2008, 197, 630–637. [Google Scholar] [CrossRef]
- Denys, A.; Hichami, A.; Khan, N.A. n-3 PUFAs modulate T-cell activation via protein kinase C-alpha and -epsilon and the NF-kappaB signaling pathway. J. Lipid Res. 2005, 46, 752–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camandola, S.; Leonarduzzi, G.; Musso, T.; Varesio, L.; Carini, R.; Scavazza, A.; Chiarpotto, E.; Baeuerle, P.A.; Poli, G. Nuclear Factor kB Is Activated by Arachidonic Acid but Not by Eicosapentaenoic Acid. Biochem. Biophys. Res. Commun. 1996, 229, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Baro, L.; Fonolla, J.; Pena, J.L.; Martinez-Ferez, A.; Lucena, A.; Jimenez, J.; Boza, J.J.; Lopez-Huertas, E. n-3 Fatty Acids Plus Oleic Acid and Vitamin Supplemented Milk Consumption Reduces Total and LDL Cholesterol, Homocysteine and Levels of Endothelial Adhesion Molecules in Healthy Humans. Clin. Nutr. 2003, 22, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.F.; Deem, T.L.; Bruce, A.C.; Reutershan, J.; Wu, D.; Ley, K. Leukocyte phosphoinositide-3 kinase γ is required for chemokine-induced, sustained adhesion under flow in vivo. J. Leukoc. Biol. 2006, 80, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, M.B.; Schaefer, C.A.; Schifferings, S.; Kuhlmann, C.R.W.; Urban, A.; Benscheid, U.; Fischer, T.; Hecker, M.; Morty, R.E.; Vadasz, I.; et al. N-3 vs. n-6 fatty acids differentially influence calcium signalling and adhesion of inflammatory activated monocytes: Impact of lipid rafts. Inflamm. Res. 2016, 65, 881–894. [Google Scholar] [CrossRef]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Hecker, M.; Sommer, N.; Foch, S.; Hecker, A.; Hackstein, H.; Witzenrath, M.; Weissmann, N.; Seeger, W.; Mayer, K. Resolvin E1 and its precursor 18 R-HEPE restore mitochondrial function in inflammation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1016–1028. [Google Scholar] [CrossRef]
- Mayer, K.; Sommer, N.; Hache, K.; Hecker, A.; Reiche, S.; Schneck, E.; Weissmann, N.; Seeger, W.; Hecker, M. Resolvin E1 Improves Mitochondrial Function in Human Alveolar Epithelial Cells during Severe Inflammation. Lipids 2019, 54, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massaro, M.; Scoditti, E.; Carluccio, M.A.; Calabriso, N.; Santarpino, G.; Verri, T.; De Caterina, R. Effects of Olive Oil on Blood Pressure: Epidemiological, Clinical, and Mechanistic Evidence. Nutrients 2020, 12, 1548. [Google Scholar] [CrossRef] [PubMed]
- Reimund, J.-M.; Scheer, O.; Muller, C.D.; Pinna, G.; Duclos, B.; Baumann, R. In vitro modulation of inflammatory cytokine production by thee lipid emulsions with different fatty acid compositions. Clin. Nutr. 2004, 23, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Blum, S.; Rössle, C.; Le Boucher, J.; Malnoë, A.; Dutot, G. Effects of Parenteral Lipid Emulsions With Different Fatty Acid Composition on Immune Cell Functions In Vitro. J. Parenter. Enter. Nutr. 2000, 24, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Buenestado, A.; Cortijo, J.; Sanz, M.-J.; Naim-Abu-Nabah, Y.; Martinez-Losa, M.; Mata, M.; Issekutz, A.C.; Martí-Bonmatí, E.; Morcillo, E.J. Olive Oil–Based Lipid Emulsion’s Neutral Effects on Neutrophil Functions and Leukocyte–Endothelial Cell Interactions. J. Parenter. Enter. Nutr. 2006, 30, 286–296. [Google Scholar] [CrossRef]
- Mateu-de Antonio, J.; Grau, S.; Luque, S.; Marín-Casino, M.; Albert, I.; Ribes, E. Comparative effects of olive oil-based and soyabean oil-based emulsions on infection rate and leucocyte count in critically ill patients receiving parenteral nutrition. Br. J. Nutr. 2008, 99, 846–854. [Google Scholar] [CrossRef]
- García-de-Lorenzo, A.; Denia, R.; Atlan, P.; Martinez-Ratero, S.; Le Brun, A.; Evard, D.; Bereziat, G. Parenteral nutrition providing a restricted amount of linoleic acid in severely burned patients: A randomised double-blind study of an olive oil-based lipid emulsion v. medium/long-chain triacylglycerols. Br. J. Nutr. 2005, 94, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimund, J.-M.; Rahmi, G.; Escalin, G.; Pinna, G.; Finck, G.; Muller, C.D.; Duclos, B.; Baumann, R. Efficacy and safety of an olive oil-based intravenous fat emulsion in adult patients on home parenteral nutrition. Aliment. Pharmacol. Ther. 2005, 21, 445–454. [Google Scholar] [CrossRef]
- Hernández, C.R.; Ponce, E.C.; Busquets, F.B.; Hernández, D.S.; Oliva, S.M.; Santacruz, E.L.; Pérez, N.; De los Santos Pelegrini, M.; Flaque, M.V. Hypersensitivity reaction to components of parenteral nutrition in pediatrics. Nutrition 2016, 32, 1303–1305. [Google Scholar] [CrossRef]
- Uthaya, S.; Liu, X.; Babalis, D.; Dore, C.J.; Warwick, J.; Bell, J.; Thomas, L.; Ashby, D.; Durighel, G.; Ederies, A.; et al. Nutritional Evaluation and Optimisation in Neonates: A randomized, double-blind controlled trial of amino acid regimen and intravenous lipid composition in preterm parenteral nutrition. Am. J. Clin. Nutr. 2016, 103, 1443–1452. [Google Scholar] [CrossRef] [Green Version]
- Pichler, J.; Simchowitz, V.; Macdonald, S.; Hill, S. Comparison of liver function with two new/mixed intravenous lipid emulsions in children with intestinal failure. Eur. J. Clin. Nutr. 2014, 68, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, G.; Simmer, K.; Deshmukh, M.; Mori, T.A.; Croft, K.D.; Kristensen, J. Fish Oil (SMOFlipid) and olive oil lipid (Clinoleic) in very preterm neonates. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 177–182. [Google Scholar] [CrossRef]
- Dai, Y.-J.; Sun, L.-L.; Li, M.-Y.; Ding, C.-L.; Su, Y.-C.; Sun, L.-J.; Xue, S.-H.; Yan, F.; Zhao, C.-H.; Wang, W. Comparison of Formulas Based on Lipid Emulsions of Olive Oil, Soybean Oil, or Several Oils for Parenteral Nutrition: A Systematic Review and Meta-Analysis. Adv. Nutr. Int. Rev. J. 2016, 7, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, K.A.; Xu, Z.; Pavlina, T.M.; Zaloga, G.P.; Siddiqui, R.A. Modulation of endothelial cell integrity and inflammatory activation by commercial lipid emulsions. Lipids Health Dis. 2015, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisramé-Helms, J.; Delabranche, X.; Klymchenko, A.; Drai, J.; Blond, E.; Zobairi, F.; Mely, Y.; Hasselmann, M.; Toti, F.; Meziani, F. Lipid emulsions differentially affect LPS-induced acute monocytes inflammation: In vitro effects on membrane remodeling and cell viability. Lipids 2014, 49, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-K.; Zhu, J.; Gong, Z.-Z.; Wen, J.; Xiao, Y.-T.; Cai, W.; Zhang, T. Olive Oil-Supplemented Lipid Emulsion Induces CELF1 Expression and Promotes Apoptosis in Caco-2 Cells. Cell. Physiol. Biochem. 2017, 41, 711–721. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid (g/100g) | LCT | SMOF |
|---|---|---|
| Medium chain: caprylic acid (C8:0) | 10 | |
| Medium chain: capric acid (C10:0) | 10 | |
| Medium chain: lauric acid (C12:0) | 10 | |
| Long chain (LC) saturated: palmitic acid (C16:0) | 10.4 | 5.9 |
| LC saturated: stearic acid (C18:0) | 4.4 | 1.8 |
| LC mono-unsaturated: oleic acid (C18:1) n-9 | 22.5 | 25 |
| LC poly-unsaturated: linoleic acid (C18:2) n-6 | 54.4 | 18.7 |
| LC poly-unsaturated: α-Linolenic acid (C18:3) n-3 | 6.7 | 8.7 |
| LC poly-unsaturated: eicosapentaenoic acid (C22:5) n-3 | 4.8 | |
| LC poly-unsaturated: docosahexaenoic acid (C22:6) n-3 | 3.5 | |
| Ratio n-6/n-3 | 8:1 | 1.3:1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hecker, M.; Rose, M.; Hecker, A.; Dietrich, H.; Schaefer, M.B.; Sommer, N.; Seeger, W.; Mayer, K. Immunomodulation by an Omega-6 Fatty Acid Reduced Mixed Lipid Emulsion in Murine Acute Respiratory Distress Syndrome. J. Clin. Med. 2020, 9, 2048. https://doi.org/10.3390/jcm9072048
Hecker M, Rose M, Hecker A, Dietrich H, Schaefer MB, Sommer N, Seeger W, Mayer K. Immunomodulation by an Omega-6 Fatty Acid Reduced Mixed Lipid Emulsion in Murine Acute Respiratory Distress Syndrome. Journal of Clinical Medicine. 2020; 9(7):2048. https://doi.org/10.3390/jcm9072048
Chicago/Turabian StyleHecker, Matthias, Matthias Rose, Andreas Hecker, Hartmut Dietrich, Martina B. Schaefer, Natascha Sommer, Werner Seeger, and Konstantin Mayer. 2020. "Immunomodulation by an Omega-6 Fatty Acid Reduced Mixed Lipid Emulsion in Murine Acute Respiratory Distress Syndrome" Journal of Clinical Medicine 9, no. 7: 2048. https://doi.org/10.3390/jcm9072048
APA StyleHecker, M., Rose, M., Hecker, A., Dietrich, H., Schaefer, M. B., Sommer, N., Seeger, W., & Mayer, K. (2020). Immunomodulation by an Omega-6 Fatty Acid Reduced Mixed Lipid Emulsion in Murine Acute Respiratory Distress Syndrome. Journal of Clinical Medicine, 9(7), 2048. https://doi.org/10.3390/jcm9072048