Circulating miRNAs as a Predictive Biomarker of the Progression from Prediabetes to Diabetes: Outcomes of a 5-Year Prospective Observational Study

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Sample Collection and Measurement
2.3. Detection of the miRNA Profiles
2.4. Validation of the NanoString Results by Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
2.5. Data Analysis
3. Results
3.1. Characteristics of the Patients
3.2. Baseline Concentrations of Circulating miRNAs
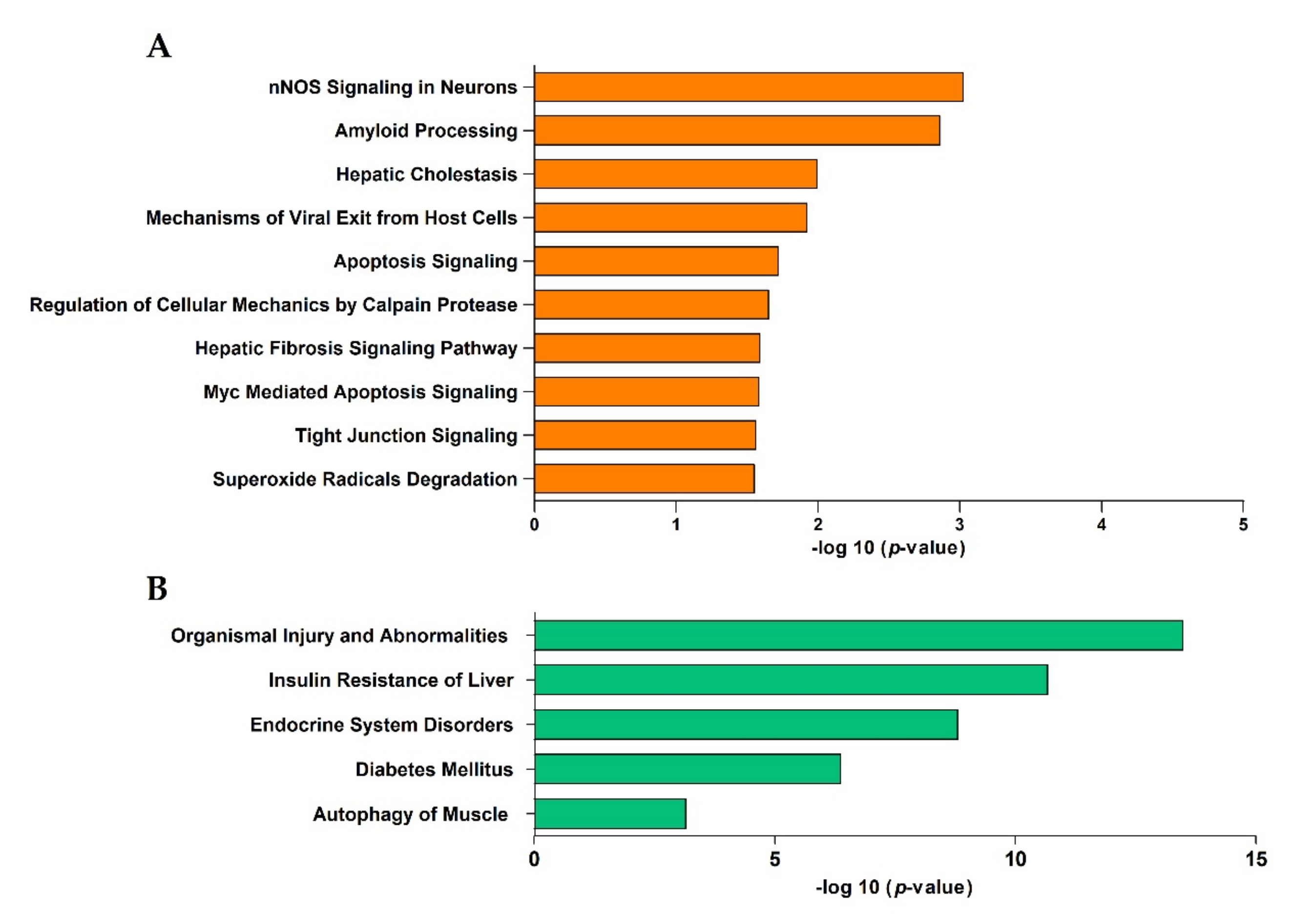
3.3. Canonical Pathway Analysis
3.4. Functional Enrichment Analysis
3.5. Hub Gene Identification
3.6. Relationship between miRNA Serum Levels and Anthropometric and Biochemical Measurements
3.7. Receiver Operating Characteristic Curve Analysis
3.8. Logistic Regression Model
3.9. Data Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharroubi, A.T. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef]
- Dorcely, B.; Katz, K.; Jagannathan, R.; Chiang, S.S.; Oluwadare, B.; Goldberg, I.J.; Bergman, M. Novel biomarkers for prediabetes, diabetes, and associated complications. Diabetes, Metab. Syndr. Obes. Targets Ther. 2017, 10, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabak, A.G.; Herder, C.; Rathman, W.; Brunner, E.J.; Kivimaki, M. Prediabetes: A high-risk state for developing diabetes. Lancet 2014, 379, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Nathan, D.M.; Davidson, M.B.; DeFronzo, R.A.; Heine, R.J.; Henry, R.R.; Pratley, R.; Zinman, B.; Kahn, R. Impaired fasting glucose and impaired glucose tolerance: Implications for care. Diabetes Care 2007, 30, 753–759. [Google Scholar] [CrossRef] [Green Version]
- Nsiah, K.; Shang, V.; Boateng, K.; Mensah, F. Prevalence of metabolic syndrome in type 2 diabetes mellitus patients. Int. J. Appl. Basic Med. Res. 2015, 5, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Salinas, C.A.; Viveros-Ruiz, T. Recent advances in managing/understanding the metabolic syndrome. F1000Research 2019, 8, F1000 Faculty Rev-370. [Google Scholar] [CrossRef] [Green Version]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Zafar, U.; Khaliq, S.; Ahmad, H.U.; Manzoor, S.; Lone, K.P. Metabolic syndrome: an update on diagnostic criteria, pathogenesis, and genetic links. Hormones 2018, 17, 299–313. [Google Scholar] [CrossRef]
- Díaz-Redondo, A.; Giráldez-García, C.; Carrillo, L.; Serrano, R.; García-Soidán, F.J.; Artola, S.; Franch, J.; Díez, J.; Ezkurra, P.; Millaruelo, J.M.; et al. Modifiable risk factors associated with prediabetes in men and women: A cross-sectional analysis of the cohort study in primary health care on the evolution of patients with prediabetes. BMC Fam. Pract. 2015, 16, 5. [Google Scholar] [CrossRef] [Green Version]
- Fowler, M.J. Microvascular and macrovascular complications of diabetes. Clin. Diabetes 2011, 29, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Satake, E.; Pezzolesi, M.G.; Dom, Z.I.M.; Smiles, A.M.; Niewczas, M.A.; Krolewski, A.S. Circulating miRNA profiles associated with hyperglycemia in patients with type 1 diabetes. Diabetes 2018, 67, 1013–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Diagnosis and classification of diabetes mellitus. Diabetes Care 2006, Suppl. 1, s43–s48.
- Keating, B.J. Advances in risk prediction of type 2 diabetes: Integrating genetic scores with framingham risk models. Diabetes 2015, 64, 1495–1497. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lee, E.S.; Yoo, J.; Kim, Y. Predicting risk of type 2 diabetes mellitus in Korean adults aged 40–69 by integrating clinical and genetic factors. Prim. Care Diabetes 2019, 13, 3–10. [Google Scholar] [CrossRef]
- Scirica, B.M. Use of biomarkers in predicting the onset, monitoring the progression, and risk stratification for patients with type 2 diabetes mellitus. Clin. Chem. 2017, 63, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xing, W.; Xie, L. Regulatory roles of microRNAs in diabetes. Int. J. Mol. Sci. 2016, 17, 1729. [Google Scholar] [CrossRef] [PubMed]
- Kamalden, T.A.; Macgregor-Das, A.M.; Kannan, S.M.; Dunkerly-Eyring, B.; Khaliddin, N.; Xu, Z.; Fusco, A.P.; Yazib, S.A.; Chow, R.C.; Duh, E.J.; et al. Exosomal MicroRNA-15a Transfer from the Pancreas Augments Diabetic Complications by Inducing Oxidative Stress. Antioxidants Redox Signal. 2017, 27, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, G.; Po, A.; Miele, E.; Ventriglia, G.; Ceccarelli, E.; Bugliani, M.; Marselli, L.; Marchetti, P.; Gulino, A.; Ferretti, E.; et al. MicroRNA-124a is hyperexpressed in type 2 diabetic human pancreatic islets and negatively regulates insulin secretion. Acta Diabetol. 2015, 52, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Li, X. MiR-375, a microRNA related to diabetes. Gene 2014, 533, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Barutta, F.; Bellini, S.; Mastrocola, R.; Bruno, G.; Gruden, G. MicroRNA and microvascular complications of diabetes. Int. J. Endocrinol. 2018, 2018, 6890501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Lucena, R.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; López-Moreno, J.; Roncero-Ramos, I.; Molina-Abril, H.; Yubero-Serrano, E.M.; Caballero-Villarraso, J.; Delgado-Lista, J.; Castaño, J.P.; et al. Circulating miRNAs as Predictive Biomarkers of Type 2 Diabetes Mellitus Development in Coronary Heart Disease Patients from the CORDIOPREV Study. Mol. Ther. - Nucleic Acids 2018, 12, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, G.; Yang, C.; Zhou, K.; Shen, B.; Liang, H.; Jiang, X. The role of circulating microRNA-126 (miR-126): A novel biomarker for screening prediabetes and newly diagnosed type 2 diabetes mellitus. Int. J. Mol. Sci. 2014, 15, 10567–10577. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.L.; Jiang, B.G.; Li, W.T.; Zou, J.J.; Shi, Y.Q.; Liu, Z.M. MicroRNA-15a positively regulates insulin synthesis by inhibiting uncoupling protein-2 expression. Diabetes Res. Clin. Pract. 2011, 91, 94–100. [Google Scholar] [CrossRef]
- Kong, L.; Zhu, J.; Han, W.; Jiang, X.; Xu, M.; Zhao, Y.; Dong, Q.; Pang, Z.; Guan, Q.; Gao, L.; et al. Significance of serum microRNAs in pre-diabetes and newly diagnosed type 2 diabetes: A clinical study. Acta Diabetol. 2011, 48, 61–69. [Google Scholar] [CrossRef]
- Nigi, L.; Grieco, G.E.; Ventriglia, G.; Brusco, N.; Mancarella, F.; Formichi, C.; Dotta, F.; Sebastiani, G. MicroRNAs as regulators of insulin signaling: Research updates and potential therapeutic perspectives in type 2 diabetes. Int. J. Mol. Sci. 2018, 19, 3705. [Google Scholar] [CrossRef] [Green Version]
- Deiuliis, J.A. MicroRNAs as regulators of metabolic disease: Pathophysiologic significance and emerging role as biomarkers and therapeutics. Int. J. Obes. 2016, 40, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, F.; D’Souza, R.F.; Durainayagam, B.R.; Milan, A.M.; Markworth, J.F.; Miranda-Soberanis, V.; Sequeira, I.R.; Roy, N.C.; Poppitt, S.D.; Mitchell, C.J.; et al. Circulatory miRNA biomarkers of metabolic syndrome. Acta Diabetol. 2020, 57, 203–214. [Google Scholar] [CrossRef]
- Lin, X.; Luo, C.; He, D.; Matro, E.; Chen, Q.; Li, H.; Zhou, J. Urinary miRNA-29a-3p levels are associated with metabolic parameters via regulation of IGF1 in patients with metabolic syndrome. Biomed. Reports 2019, 10, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Carrasco-Wong, I.; Dominguez, A.; Arnaiz, P.; Farías, M.; Barja, S.; Mardones, F.; Casanello, P. Micro-RNAs Let7e and 126 in plasma as markers of metabolic dysfunction in 10 to 12 years old children. PLoS One 2015, 10, e0128140. [Google Scholar] [CrossRef]
- Al-Rawaf, H.A. Circulating microRNAs and adipokines as markers of metabolic syndrome in adolescents with obesity. Clin. Nutr. 2019, 38, 2231–2238. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hong, J.; Cao, Y.; Shi, J.; Gu, W.; Ning, G.; Zhang, Y.; Wang, W. Elevated circulating microRNA-122 is associated with obesity and insulin resistance in young adults. Eur. J. Endocrinol. 2015, 172, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Zaiou, M.; El Amri, H.; Bakillah, A. The clinical potential of adipogenesis and obesity-related microRNAs. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrián, S.; González-Muniesa, P.; Milagro, F.I.; Alfredo Martínez, J. MicroRNAs and other non-coding RNAs in adipose tissue and obesity: Emerging roles as biomarkers and therapeutic targets. Clin. Sci. 2019, 133, 23–40. [Google Scholar] [CrossRef]
- Bansal, N. Prediabetes diagnosis and treatment: A review. World J. Diabetes 2015, 6, 296–303. [Google Scholar] [CrossRef]
- 2018 Guidelines on the management of diabetic patients. Clin. Diabetol. 2018, 7, 1–90. [CrossRef]
- Radziuk, J. Homeostastic model assessment and insulin sensitivity/resistance. Diabetes 2014, 63, 1850–1854. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M. Quantification strategies in real-time PCR. In A-Z of quantitative PCR; Bustin, S.A., Ed.; International University Line : San Diego, CA, USA, 2004; pp. 87–120. ISBN 0963681788. [Google Scholar]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics 2011, 12, 1–8. [Google Scholar] [CrossRef]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinformatics 2009, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Incani, M.; Sentinelli, F.; Perra, L.; Pani, M.G.; Porcu, M.; Lenzi, A.; Cavallo, M.G.; Cossu, E.; Leonetti, F.; Baroni, M.G. Glycated hemoglobin for the diagnosis of diabetes and prediabetes: Diagnostic impact on obese and lean subjects, and phenotypic characterization. J. Diabetes Investig. 2015, 6, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostalek, U. Global epidemiology of prediabetes - present and future perspectives. Clin. Diabetes Endocrinol. 2019, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farr, O.M.; Mantzoros, C.S. Treatment options to prevent diabetes in subjects with prediabetes: Efficacy, cost effectiveness and future outlook. Metabolism 2017, 70, 192–195. [Google Scholar] [CrossRef]
- Ezquerra-Lázaro, I.; Cea-Soriano, L.; Giraldez-García, C.; Ruiz, A.; Franch-Nadal, J.; Diez-Espino, J.; Nogales, P.; Carramiñana, F.; Javier Sangros, F.; Regidor, E. Lifestyle factors do not explain the difference on diabetes progression according to type of prediabetes: Results from a Spanish prospective cohort of prediabetic patients. Diabetes Res. Clin. Pract. 2019, 153, 66–75. [Google Scholar] [CrossRef]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA Signatures as Future Biomarkers for Diagnosis of Diabetes States. Cells 2019, 8, 1533. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Li, L.; Shang, Q.; Lv, C.F.; Wang, C.Y.; Su, B. Circulating miR-126 is a potential biomarker to predict the onset of type 2 diabetes mellitus in susceptible individuals. Biochem. Biophys. Res. Commun. 2015, 463, 60–63. [Google Scholar] [CrossRef]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma MicroRNA profiling reveals loss of endothelial MiR-126 and other MicroRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef]
- Pordzik, J.; Jakubik, D.; Jarosz-Popek, J.; Wicik, Z.; Eyileten, C.; De Rosa, S.; Indolfi, C.; Siller-Matula, J.M.; Czajka, P.; Postula, M. Significance of circulating microRNAs in diabetes mellitus type 2 and platelet reactivity: Bioinformatic analysis and review. Cardiovasc. Diabetol. 2019, 18, 113. [Google Scholar] [CrossRef] [Green Version]
- Chuang, T.Y.; Wu, H.L.; Chen, C.C.; Gamboa, G.M.; Layman, L.C.; Diamond, M.P.; Azziz, R.; Chen, Y.H. MicroRNA-223 expression is upregulated in insulin resistant human adipose tissue. J. Diabetes Res. 2015, 943659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanyam, M.; Aravind, S.; Gokulakrishnan, K.; Prabu, P.; Sathishkumar, C.; Ranjani, H.; Mohan, V. Impaired miR-146a expression links subclinical inflammation and insulin resistance in Type 2 diabetes. Mol. Cell. Biochem. 2011, 351, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Lamadrid-Romero, M.; Solís, K.H.; Cruz-Reséndiz, M.S.; Pérez, J.E.; Díaz, N.F.; Flores-Herrera, H.; García-López, G.; Perichart, O.; Reyes-Muñoz, E.; Arenas-Huertero, F.; et al. Central nervous system development-related microRNAs levels increase in the serum of gestational diabetic women during the first trimester of pregnancy. Neurosci. Res. 2018, 130, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, H.; Si, H.; Li, X.; Ding, X.; Sheng, Q.; Chen, P.; Zhang, H. Serum miR-23a, a potential biomarker for diagnosis of pre-diabetes and type 2 diabetes. Acta Diabetol. 2014, 51, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Boissonneault, V.; Plante, I.; Rivest, S.; Provost, P. MicroRNA-298 and microRNA-328 regulate expression of mouse β-amyloid precursor protein-converting enzyme 1. J. Biol. Chem. 2009, 284, 1971–1981. [Google Scholar] [CrossRef] [Green Version]
- Provost, P. Interpretation and applicability of microrna datato the context of Alzheimer’s and age-related diseases. Aging (Albany. NY) 2010, 2, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Motti, D.; Lerch, J.K.; Danzi, M.C.; Gans, J.H.; Kuo, F.; Slepak, T.I.; Bixby, J.L.; Lemmon, V.P. Identification of miRNAs involved in DRG neurite outgrowth and their putative targets. FEBS Lett. 2017, 591, 2091–2105. [Google Scholar] [CrossRef] [Green Version]
- De Felice, F.G.; Ferreira, S.T. Inflammation, defective insulin signaling, and mitochondrial dysfunction as common molecular denominators connecting type 2 diabetes to Alzheimer Disease. Diabetes 2014, 63, 2262–2272. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Mudher, A. Alzheimer’s disease and type 2 diabetes: A critical assessment of the shared pathological traits. Front. Neurosci. 2018, 12, 383. [Google Scholar] [CrossRef] [Green Version]
- Tumminia, A.; Vinciguerra, F.; Parisi, M.; Frittitta, L. Type 2 diabetes mellitus and alzheimer’s disease: Role of insulin signalling and therapeutic implications. Int. J. Mol. Sci. 2018, 19, 3306. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.I.; Carvalho, C.; Zhu, X.; Smith, M.A.; Perry, G. Mitochondrial dysfunction is a trigger of Alzheimer’s disease pathophysiology. Biochim. Biophys. Acta - Mol. Basis Dis. 2010, 1802, 2–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Gonzalez, I.; Edwards, G.; Salvadores, N.; Shahnawaz, M.; Diaz-Espinoza, R.; Soto, C. Molecular interaction between type 2 diabetes and Alzheimer’s disease through cross-seeding of protein misfolding. Mol. Psychiatry 2017, 22, 1327–1334. [Google Scholar] [CrossRef] [Green Version]
- Donath, M.Y.; Shoelson, S.E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef]
- Masters, S.L.; Dunne, A.; Subramanian, S.L.; Hull, R.L.; Tannahill, G.M.; Sharp, F.A.; Becker, C.; Franchi, L.; Yoshihara, E.; Chen, Z.; et al. Activation of the NLRP3 inflammasome by islet amyloid polypeptide provides a mechanism for enhanced IL-1β 2 in type 2 diabetes. Nat. Immunol. 2010, 11, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhai, Y.X.; Liu, H.Q.; Shi, Y.A.; Li, X.B. MicroRNA-491-5p suppresses cervical cancer cell growth by targeting hTERT. Oncol. Rep. 2015, 34, 979–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Liu, Z.; Tong, D.; Yang, Y.; Guo, B.; Wang, X.; Zhao, L.; Huang, C. MiR-491-5p, mediated by Foxi1, functions as a tumor suppressor by targeting Wnt3a/β-catenin signaling in the development of gastric cancer. Cell Death Dis. 2017, 30, e2714. [Google Scholar] [CrossRef] [PubMed]
- Hui, Z.; Yiling, C.; Wenting, Y.; Xuqun, H.; Chuanyi, Z.; Hui, L. miR-491-5p functions as a tumor suppressor by targeting JMJD2B in ERα-positive breast cancer. FEBS Lett. 2015, 589, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Ding, H.; He, E.; Chen, J.; Li, M. Up-regulation of microRNA-491-5p suppresses cell proliferation and promotes apoptosis by targeting FOXP4 in human osteosarcoma. Cell Prolif. 2017, 50, e12308. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, L.; Yao, B.; Liu, Q.; Guoa, C. miR-1307-3p promotes tumor growth and metastasis of hepatocellular carcinoma by repressing DAB2 interacting protein. Biomed. Pharmacother. 2019, 117, 109055. [Google Scholar] [CrossRef]
- Han, S.; Zou, H.; Lee, J.-W.; Han, J.; Kim, H.C.; Cheol, J.J.; Kim, L.-S.; Kim, H. miR-1307-3p Stimulates Breast Cancer Development and Progression by Targeting SMYD4. J. Cancer 2019, 10, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Zheng, Y.; Lei, W.; Xiang, L.; Chen, M. miR-1307–3p overexpression inhibits cell proliferation and promotes cell apoptosis by targeting ISM1 in colon cancer. Mol. Cell. Probes 2019, 48, 101445. [Google Scholar] [CrossRef] [PubMed]
- Srodulski, S.; Sharma, S.; Bachstetter, A.B.; Brelsfoard, J.M.; Pascual, C.; Xie, X.S.; Saatman, K.E.; Van Eldik, L.J.; Despa, F. Neuroinflammation and neurologic deficits in diabetes linked to brain accumulation of amylin. Mol. Neurodegener. 2014, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Morales-Scheihing, D.; Salvadores, N.; Moreno-Gonzalez, I.; Gonzalez, C.; Taylor-Presse, K.; Mendez, N.; Shahnawaz, M.; Gaber, A.O.; Sabek, O.M.; et al. Induction of IAPP amyloid deposition and associated diabetic abnormalities by a prion-like mechanism. J. Exp. Med. 2017, 214, 2591–2610. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Cao, P.; Noor, H.; Ridgway, Z.; Tu, L.H.; Wang, H.; Wong, A.G.; Zhang, X.; Abedini, A.; Schmidt, A.M.; et al. Islet Amyloid Polypeptide: Structure, Function, and Pathophysiology. J. Diabetes Res. 2016, 2016, 2798269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimundo, A.F.; Ferreira, S.; Martins, I.C.; Menezes, R. Islet Amyloid Polypeptide: A Partner in Crime With Aβ in the Pathology of Alzheimer’s Disease. Front. Mol. Neurosci. 2020, 13, 35. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, D.Y. Neuronal nitric oxide synthase: Structure, subcellular localization, regulation, and clinical implications. Nitric Oxide - Biol. Chem. 2009, 20, 223–230. [Google Scholar] [CrossRef]
- Giove, T.J.; Deshpande, M.M.; Gagen, C.S.; Eldred, W.D. Increased neuronal nitric oxide synthase activity in retinal neurons in early diabetic retinopathy. Mol. Vis. 2009, 15, 2249–2258. [Google Scholar]
- Fonseca, V.A.; Handelsman, Y.; Staels, B. Colesevelam lowers glucose and lipid levels in type 2 diabetes: The clinical evidence. Diabetes Obes. Metab. 2010, 12, 384–392. [Google Scholar] [CrossRef]
- Hansen, M.; Sonne, D.P.; Mikkelsen, K.H.; Gluud, L.L.; Vilsbøll, T.; Knop, F.K. Bile acid sequestrants for glycemic control in patients with type 2 diabetes: A systematic review with meta-analysis of randomized controlled trials. J. Diabetes Complications 2017, 31, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Cao, J.; Liu, G.; Huo, J.R. Function and mechanism of F-box proteins in gastric cancer (Review). Int. J. Oncol. 2015, 47, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, C.; Holubowska, A.; Schwedhelm-Domeyer, N.; Mitkovski, M.; Lee, S.J.; Kannan, M.; Matz, A.; Vadhvani, M.; Stegmüller, J. Loss of the neuron-specific F-box protein FBXO41 models an ataxia-like phenotype in mice with neuronal migration defects and degeneration in the cerebellum. J. Neurosci. 2015, 35, 8701–8717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhipa, A.S.; Borse, S.P.; Baksi, R.; Lalotra, S.; Nivsarkar, M. Targeting receptors of advanced glycation end products (RAGE): Preventing diabetes induced cancer and diabetic complications. Pathol. Res. Pract. 2019, 11, 152643. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.; Yan, S.F.; Schmidt, A.M. Receptor for AGE (RAGE): Signaling mechanisms in the pathogenesis of diabetes and its complications. Ann. N. Y. Acad. Sci. 2011, 1243, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, B.T.; Bhardwaj, G.; Penniman, C.M.; Krumpoch, M.T.; Suarez Beltran, P.A.; Klaus, K.; Poro, K.; Li, M.; Pan, H.; Dreyfuss, J.M.; et al. FOXO transcription factors are critical regulators of diabetes-related muscle atrophy. Diabetes 2019, 68, 556–570. [Google Scholar] [CrossRef] [Green Version]
- Perry, B.D.; Caldow, M.K.; Brennan-Speranza, T.C.; Sbaraglia, M.; Jerums, G.; Garnham, A.; Wong, C.; Levinger, P.; Asrar Ul Haq, M.; Hare, D.L.; et al. Muscle atrophy in patients with Type 2 Diabetes Mellitus: Roles of inflammatory pathways, physical activity and exercise. Exerc. Immunol. Rev. 2016, 22, 94–109. [Google Scholar]
- Patil, G.; Li, S. Tripartite motif proteins: An emerging antiviral protein family. Future Virol. 2019, 14, 107–122. [Google Scholar] [CrossRef]
- Patil, G.; Zhao, M.; Song, K.; Hao, W.; Bouchereau, D.; Wang, L.; Li, S. TRIM41-Mediated Ubiquitination of Nucleoprotein Limits Influenza A Virus Infection. J. Virol. 2018, 92, e00905–e00918. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Pan, Z.; Yang, X. Key genes and co-expression network analysis in the livers of type 2 diabetes patients. J. Diabetes Investig. 2019, 10, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Dastsooz, H.; Cereda, M.; Donna, D.; Oliviero, S. A comprehensive bioinformatics analysis of UBE2C in cancers. Int. J. Mol. Sci. 2019, 20, 2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlicher, S.E.; Stierwalt, H.D.; Newsom, S.A.; Robinson, M.M. Skeletal muscle autophagy remains responsive to hyperinsulinemia and hyperglycemia at higher plasma insulin concentrations in insulin-resistant mice. Physiol. Rep. 2018, 6, e13810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, M. Protein breakdown in muscle wasting: Role of autophagy-lysosome and ubiquitin-proteasome. Int. J. Biochem. Cell Biol. 2013, 45, 2121–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambashivaiah, S.; Harridge, S.D.R.; Sharma, N.; Selvam, S.; Rohatgi, P.; Kurpad, A.V. Asian Indians With Prediabetes Have Similar Skeletal Muscle Mass and Function to Those With Type 2 Diabetes. Front. Nutr. 2019, 6, 179. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Purohit, P.; Sharma, P. MicroRNAs and Diseases: Promising Biomarkers for Diagnosis and Therapeutics. Indian J. Clin. Biochem. 2019, 34, 243–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genomics 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Faruq, O.; Vecchione, A. microRNA: Diagnostic perspective. Front. Med. 2015, 2, 51. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control N = 18 | T2DM N = 24 | |||||
|---|---|---|---|---|---|---|
| Variable | Visit 1 a | Visit 2 b | Visit 1 a | Visit 2 b | p-Value Visit 1 c | p-Value Visit 2 d |
| Age [years] | 54.57 (33.43–65.91) | 56. 36 (37.36–70.96) | 58.30 (28.88–65.12) | 62.50 (41.16–69.20) | 0.157 | 0.146 |
| Female/Male | 9/9 | 11/13 | ||||
| BMI [kg/m2] | 32.40 (25.51–46.05) | 32.48 (23.66–47.05) | 31.52 (22.85–47.22) | 32.10 (26.71–49.35) | 0.187 | 0.219 |
| Weight [kg] | 98.75 (68.8–145.9) | 101.55 (64.00–144.10) | 89.30 (60.20–162.10) | 90.55 (65.80–176.30) | 0.113 | 0.160 |
| Fasting glucose 0 min [mg/dL] | 106.50 (100.00–117.00) | 108.00 (101.00–121.00) | 107.00 (84.00–128.00) | 129.00 (100.00–171.00) | 0.876 | 0.0018 |
| Glucose 120 min [mg/dL] | 124.50 (68.00–182.00) | 127.00 (72.00–190.00) | 141.40 (69.00–186.00) | 206 (160.00–229.00) | 0.132 | 0.0001 |
| Insulin [µU/mL] | 13.49 (5.60–29.00) | 15.00 (10.00–35.00) | 12.00 (6.00–35.00) | 16.00 (4.70–59.00) | 0.618 | 0.88 |
| HbA1c [%] | 5.80 (4.10–6.40) | 5.80 (5.10–6.40) | 5.95 (4.90–6.60) | 6.15 (5.30–7.70) | 0.308 | 0.0057 |
| LDL cholesterol [mg/dL] | 123.10 (94.00–228.00) | 105.00 (53.00–222.00) | 118.50 (57.00–213.00) | 95.50 (60.00–213.00 | 0.471 | 0.348 |
| Total cholesterol [mg/dL] | 196.00 (166.00–324.00) | 181.00 (125.00–284.00) | 191.00 (129.00–321.00) | 174.00 (138.00–310.00) | 0.458 | 0.723 |
| HDL cholesterol [mg/dL] | 49.50 (32.00–107.00) | 49.70 (29.00–125.00) | 51.70 (40.00–71.00) | 53.00 (36.00–89.00) | 0.517 | 0.319 |
| Triglyceride [mg/dL] | 132.00 (41.00–227.00) | 107.00 (33.00–229.00) | 112.50 (45.00–491.00) | 124.50 (44.00–232.00) | 0.131 | 0.875 |
| HOMA-IR | 3.40 (1.50–7.80) | 4.30 (2.80–10.40) | 2.95 (1.52–9.40) | 5.20 (1.10–20.00) | 0.783 | 0.479 |
| HOMA-B | 112.38 (43.00–275.00) | 112.00 (71.00–216.00) | 97.00 (42.23–349.00) | 85.00 (19.00–277.00) | 0.687 | 0.112 |
| miRNA | FC | FDR |
|---|---|---|
| miR-298 | 1.95 | 0.05 |
| miR-491-5p | 1.95 | 0.01 |
| miR-1307-3p | 1.85 | 0.02 |
| AUC (CI) | Cut-Off Point | Specificity | Sensitivity | Accuracy | TP/TN/FP/FN | Intercept (a0) | Coefficients | |
|---|---|---|---|---|---|---|---|---|
| x1 = miR-298 | 95.70% | 1.5 | 100% | 91.30% | 0.95 | 17/2/0/21 | –649.78 | a1 = 31.25 |
| x2 = miR-1307-3p | a2 = 27.99 | |||||||
| x3 = miR-491-5p | a3 = 81.50 | |||||||
| x1 = miR-298 | 82.50% | 1.5 | 82.35% | 82.60% | 0.83 | 14/4/3/19 | −5.99 | a1 = 0.64 |
| x2 = miR-1307-3p | a2 = 0.70 | |||||||
| x1 = miR-298 | 92.70% | 1.5 | 94.20% | 91.30% | 0.93 | 16/2/1/21 | −16.81 | a1 = 1.32 |
| x2 = miR-491-5p | a2 = 2.53 | |||||||
| x1 = miR-1307-3p | 87.60% | 1.5 | 88.23% | 86.96% | 0.86 | 15/3/2/20 | −23.09 | a1 = 1.63 |
| x2 = miR-491-5p | a2 = 3.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorkiewicz, I.; Niemira, M.; Maliszewska, K.; Erol, A.; Bielska, A.; Szalkowska, A.; Adamska-Patruno, E.; Szczerbinski, L.; Gorska, M.; Kretowski, A. Circulating miRNAs as a Predictive Biomarker of the Progression from Prediabetes to Diabetes: Outcomes of a 5-Year Prospective Observational Study. J. Clin. Med. 2020, 9, 2184. https://doi.org/10.3390/jcm9072184
Sidorkiewicz I, Niemira M, Maliszewska K, Erol A, Bielska A, Szalkowska A, Adamska-Patruno E, Szczerbinski L, Gorska M, Kretowski A. Circulating miRNAs as a Predictive Biomarker of the Progression from Prediabetes to Diabetes: Outcomes of a 5-Year Prospective Observational Study. Journal of Clinical Medicine. 2020; 9(7):2184. https://doi.org/10.3390/jcm9072184
Chicago/Turabian StyleSidorkiewicz, Iwona, Magdalena Niemira, Katarzyna Maliszewska, Anna Erol, Agnieszka Bielska, Anna Szalkowska, Edyta Adamska-Patruno, Lukasz Szczerbinski, Maria Gorska, and Adam Kretowski. 2020. "Circulating miRNAs as a Predictive Biomarker of the Progression from Prediabetes to Diabetes: Outcomes of a 5-Year Prospective Observational Study" Journal of Clinical Medicine 9, no. 7: 2184. https://doi.org/10.3390/jcm9072184
APA StyleSidorkiewicz, I., Niemira, M., Maliszewska, K., Erol, A., Bielska, A., Szalkowska, A., Adamska-Patruno, E., Szczerbinski, L., Gorska, M., & Kretowski, A. (2020). Circulating miRNAs as a Predictive Biomarker of the Progression from Prediabetes to Diabetes: Outcomes of a 5-Year Prospective Observational Study. Journal of Clinical Medicine, 9(7), 2184. https://doi.org/10.3390/jcm9072184