
Effects of Phosphorus on Nitrification Process in a Fertile Soil Amended with Urea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Used
2.2. Laboratory Incubation Study
2.3. Soil Analysis and Nutrient Extraction
2.4. DNA Extraction and Quantitative PCR (qPCR) of the amoA Genes
2.5. Statistical Analysis
3. Results
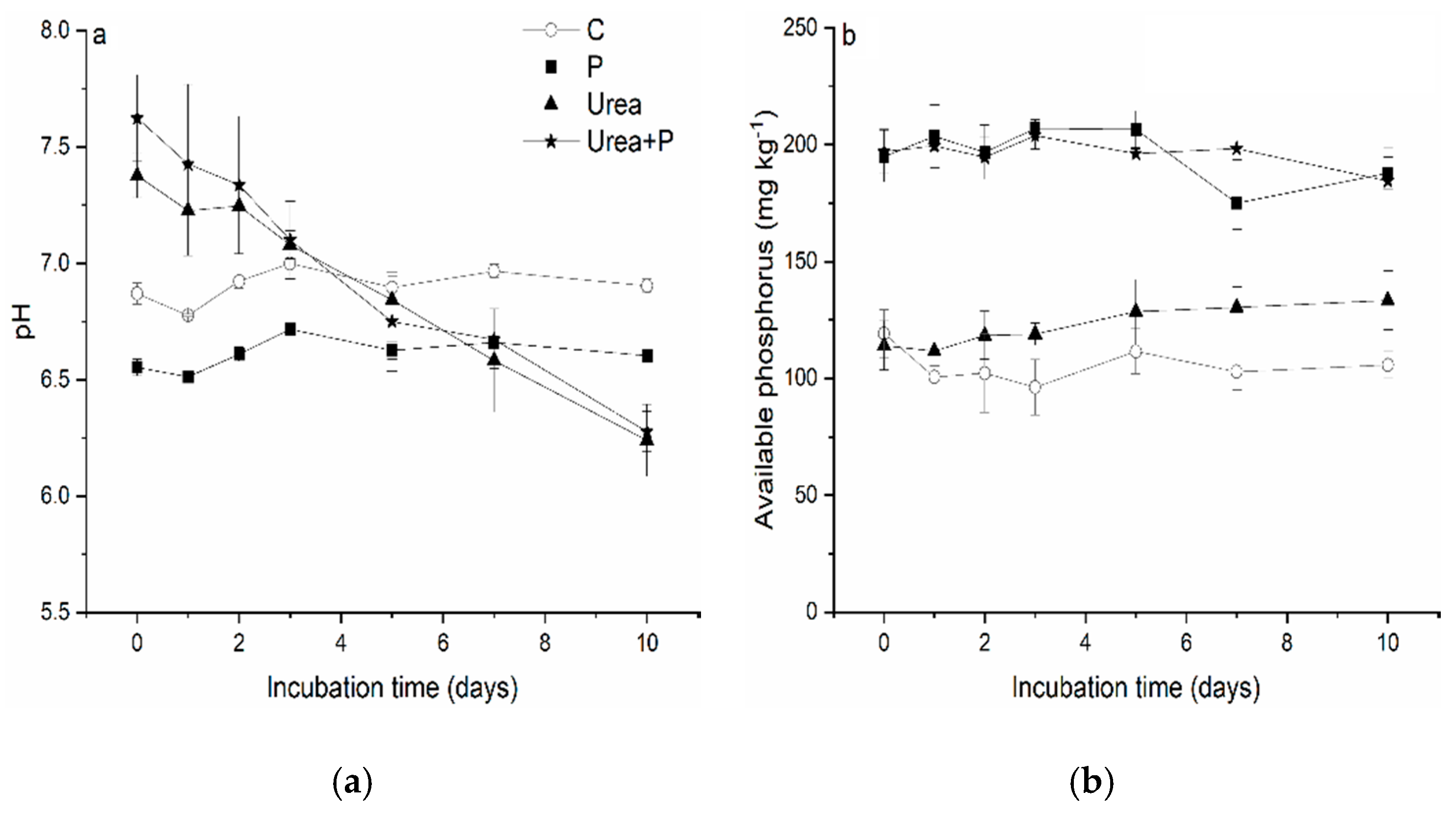
3.1. Soil pH and Available P
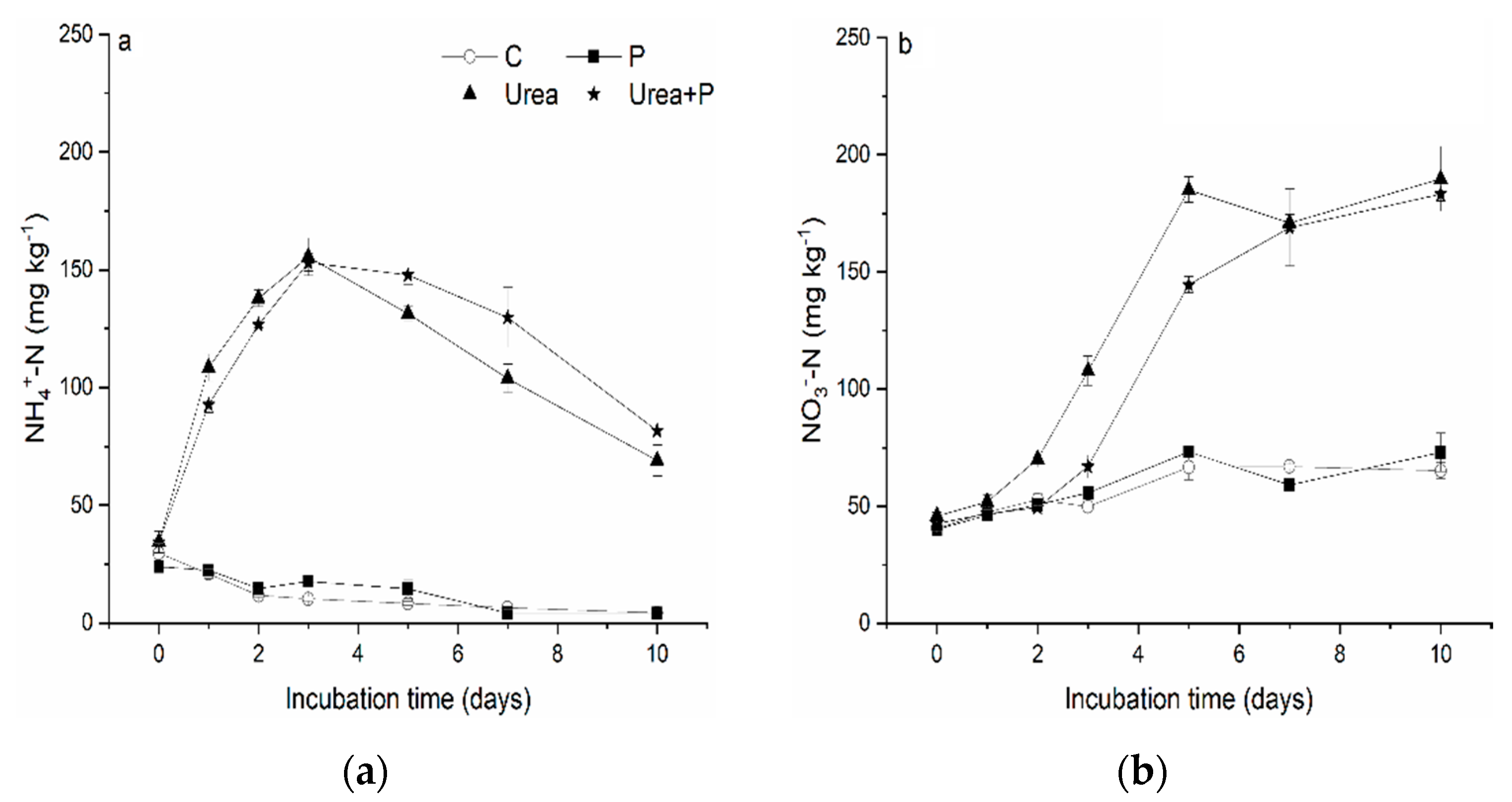
3.2. Changes in NH4+-N
3.3. Soil Nitrification
3.4. Soil Stoichiometry of N:P Ratio
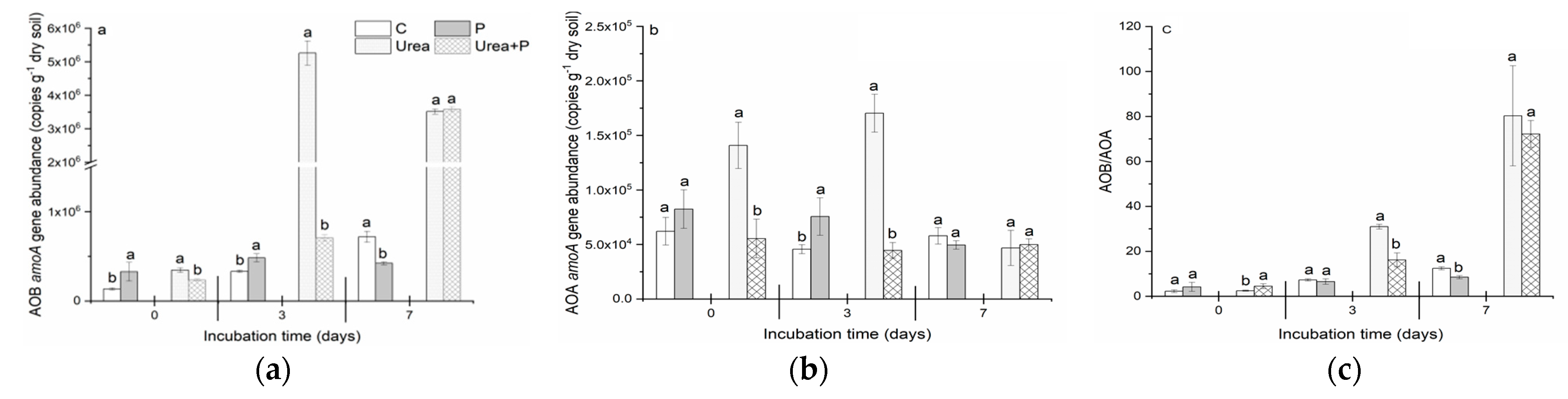
3.5. Abundances of Bacterial and Archaeal amoA Genes
3.6. Correlations between Soil Ammonia-Oxidizing Microorganisms and Soil Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klotz, M.G.; Alzerreca, J.; Norton, J.M. A gene encoding a membrane protein exists upstream of the amoA/amoB genes in ammonia-oxidizing bacteria: A third member of the amo operon? FEMS Microbiol. Lett. 1997, 150, 65–73. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Annu. Rev. Microbiol. 2001, 55, 485–592. [Google Scholar] [CrossRef]
- Li, Y.; Chapman, S.J.; Nicol, G.W.; Yao, H. Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 2018, 116, 290–301. [Google Scholar] [CrossRef]
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G.W.; Prosser, J.I.; Schuster, S.C.; Schleper, C. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 2006, 442, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Deboer, W.; Gunnewiek, P.; Veenhuis, M.; Bock, E.; Laanbroek, H.J. Nitrification at low pH by aggregated chemolithotrophic bacteria. Appl. Environ. Microbiol. 1991, 57, 3600–3604. [Google Scholar] [CrossRef] [PubMed]
- He, J.Z.; Hu, H.W.; Zhang, L.M. Current insights into the autotrophic thaumarchaeal ammonia oxidation in acidic soils. Soil Biol. Biochem. 2012, 55, 146–154. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialisation and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef]
- Qin, H.; Yuan, H.; Zhang, H.; Zhu, Y.; Yin, C.; Tan, Z.; Wu, J.; Wei, W. Ammonia-oxidizing archaea are more important than ammonia-oxidizing bacteria in nitrification and NO3−-N loss in acidic soil of sloped land. Biol. Fertil. Soils 2013, 49, 767–776. [Google Scholar] [CrossRef]
- Wang, X.; Han, C.; Zhang, J.; Huang, Q.; Deng, H.; Deng, Y.; Zhong, W. Long-term fertilization effects on active ammonia oxidizers in an acidic upland soil in China. Soil Biol. Biochem. 2015, 84, 28–37. [Google Scholar] [CrossRef]
- Isobea, K.; Ikutani, J.; Fang, Y.; Yoh, M.; Mo, J.; Suwa, Y.; Yoshida, M.; Senoo, K.; Otsuka, S.; Koba, K. Highly abundant acidophilic ammonia-oxidizing archaea causes high rates of nitrification and nitrate leaching in nitrogen-saturated forest soils. Soil Biol. Biochem. 2018, 122, 220–227. [Google Scholar] [CrossRef]
- Levičnik-Höfferle, Š.; Nicol, G.W.; Ausec, L.; Mandic-Mulec, I.; Prosser, J.I. Stimulation of thaumarchaeal ammonia oxidation by ammonia derived from organic nitrogen but not added inorganic nitrogen. FEMS Microbiol. Ecol. 2012, 80, 114–123. [Google Scholar] [CrossRef]
- Wu, Y.; Conrad, R. Ammonia oxidation-dependent growth of group I.1b Thaumarchaeota in acidic red soil microcosms. FEMS Microbiol. Ecol. 2014, 89, 127–134. [Google Scholar] [CrossRef]
- Stopnisek, N.; Gubry-Rangin, C.; Hofferle, S.; Nicol, G.W.; Mandic-Mulec, I.; Prosser, J.I. Thaumarchaeal ammonia oxidation in an acidic forest peat soil is not influenced by ammonium amendment. Appl. Environ. Microbiol. 2010, 76, 7626–7634. [Google Scholar] [CrossRef]
- Hu, B.L.; Liu, S.; Wang, W.; Shen, L.D.; Lou, L.P.; Liu, W.P.; Tian, G.M.; Xu, X.Y.; Zheng, P. pH-dominated niche segregation of ammonia-oxidising microorganisms in Chinese agricultural soils. FEMS Microbiol. Ecol. 2014, 90, 290–299. [Google Scholar]
- Che, J.; Zhao, X.Q.; Zhou, X.; Jia, Z.J.; Shen, R.F. High pH-enhanced soil nitrification was associated with ammonia-oxidizing bacteria rather than archaea in acidic soils. Appl. Soil Ecol. 2015, 85, 21–29. [Google Scholar] [CrossRef]
- Martens-Habbena, W.M.; Berube, P.; Urakawa, H.; de la Torre, J.R.; Stahl, D.A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 2009, 461, 976–979. [Google Scholar] [CrossRef]
- Taylor, A.E.; Zeglin, L.H.; Dooley, S.; Myrold, D.D.; Bottomley, P.J. Evidence for different contributions of archaea and bacteria to the ammonia-oxidizing potential of diverse Oregon soils. Appl. Environ. Microbiol. 2010, 76, 7691–7698. [Google Scholar] [CrossRef] [PubMed]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.Z. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nat. Geosci. 2009, 2, 621–624. [Google Scholar] [CrossRef]
- Jia, Z.; Conrad, R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil. Environ. Microbiol. 2009, 11, 1658–1671. [Google Scholar] [CrossRef] [PubMed]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W. Different roles of rhizosphere effect and long-term fertilization in the activity and community structure of ammonia oxidizers in a calcareous fluvo-aquic soil. Soil Biol. Biochem. 2013, 57, 30–42. [Google Scholar] [CrossRef]
- Xia, W.; Zhang, C.; Zeng, X.; Feng, Y.; Weng, J.; Lin, X.; Zhu, J.; Xiong, Z.; Xu, J.; Cai, Z.; et al. Autotrophic growth of nitrifying community in an agricultural soil. ISME J. 2011, 5, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Fujii, T.; Morimoto, S.; Lin, X.; Yagi, K.; Hu, J.; Zhang, J. Community structure of ammonia-oxidizing bacteria under long-term application of mineral fertilizer and organic manure in a sandy loam soil. Appl. Environ. Microbiol. 2007, 73, 485–491. [Google Scholar] [CrossRef]
- Wu, Y.; Lu, L.; Wang, B.; Lin, X.; Zhu, J.; Cai, Z.; Yan, X.; Jia, Z. Long-term field fertilization significantly alters community structure of Ammonia-oxidizing Bacteria rather than Archaea in a Paddy soil. Soil Sci. Soc. Am. J. 2011, 75, 1431–1439. [Google Scholar] [CrossRef]
- Tzanakakis, V.A.; Paranychianakis, N.V. Divergent response of ammonia oxidizers to various amino acids. Appl. Soil Ecol. 2017, 114, 45–51. [Google Scholar] [CrossRef]
- Ouyang, Y.; Norton, J.M.; Stark, J.M.; Reeve, J.R.; Habteselassie, M.Y. Ammonia-oxidizing bacteria are more responsive than archaea to nitrogen source in an agricultural soil. Soil Biol. Biochem. 2016, 96, 4–15. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Shen, J.-P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.-Z. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiol. Ecol. 2010, 72, 386–394. [Google Scholar] [CrossRef]
- Erguder, T.H.; Boon, N.; Wittebolle, L.; Marzorati, M.; Verstraete, W. Environmental factors shaping the ecological niches of ammoniaoxidizing archaea. FEMS Microbiol Rev. 2009, 33, 855–869. [Google Scholar] [CrossRef] [PubMed]
- Spohn, M. Element cycling as driven by stoichiometric homeostasis of soil microorganisms. Basic Appl. Ecol. 2016, 17, 471–478. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, X.; Li, D.; Wang, H.; Chen, F.; Fu, X.; Fang, X.; Sun, X.; Yu, G. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, T.T.; Qian, H.Y.; Fan, J.B.; He, Y.Q.; Sun, B. Nitrogen mineralization as a result of phosphorus supplementation in long-term phosphate deficient soil. Appl. Soil Ecol. 2016, 106, 24–32. [Google Scholar] [CrossRef]
- DeForest, J.L.; Otuya, R.K. Soil nitrification increases with elevated phosphorus or soil pH in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 2020, 142, 107716. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Sun, N.; Xu, M.; Zhang, J.; Cai, Z.; Wang, S. Phosphorus addition enhances gross microbial N cycling in phosphorus-poor soils: A 15N study from two long-term fertilization experiments. Biol. Fertil. Soil 2018, 54, 783–789. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.; Gurmesa, G.A.; Zhu, X.; Li, D.; Mo, J. Phosphorus addition affects soil nitrogen dynamics in a nitrogen-saturated and two nitrogen-limited forests. Eur. J. Soil Biol. 2017, 68, 472–479. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Phosphorus addition enhances loss of nitrogen in a phosphorus-poor soil. Soil Biol. Biochem. 2015, 82, 99–106. [Google Scholar] [CrossRef]
- Mehnaz, K.R.; Dijkstra, F.A. Denitrification and associated N2O emissions are limited by phosphorus availability in a grassland soil. Geoderma 2016, 284, 34–41. [Google Scholar] [CrossRef]
- O’Neill, R.M.; Krol, D.J.; Wall, D.; Lanigan, G.J.; Renou-Wilson, F.; Richards, K.G.; Jansen-Willems, A.B.; Müller, C. Assessing the impact of long-term soil phosphorus on N-transformation pathways using 15N tracing. Soil Biol. Biochem. 2021, 152, 108066. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, H.; Wang, R.; Guo, S. Responses of nitrification and denitrification to nitrogen and phosphorus fertilization: Does the intrinsic soil fertility matter? Plant Soil 2019, 440, 443–456. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA Circular 939; US Government Printing Office: Washington, DC, USA, 1954.
- Lu, R.K. Methods of Agricultural Chemical Analysis of Soil; China Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Verchot, L.V.; Holmes, Z.; Mulon, L.; Groffman, P.M.; Lovett, G.M. Gross vs net rates of N mineralization and nitrification as indicators of functional differences between forest types. Soil Biol. Biochem. 2001, 33, 1889–1901. [Google Scholar] [CrossRef]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed]
- Horz, H.P.; Barbrook, A.; Field, C.B.; Bohannan, B.J.M. Ammonia-oxidizing bacteria respond to multifactorial global change. Proc. Natl. Acad. Sci. USA 2004, 101, 15136–15141. [Google Scholar] [CrossRef]
- Jiang, X.; Shi, X.; Liu, W.; Wright, A. Kinetics of net nitrification associated with soil aggregates under conventional and no-tillage in a subtropical rice soil. Plant Soil 2011, 347, 305–312. [Google Scholar] [CrossRef]
- Xin, X.; Jiang, X.; Su, J.; Yan, X.; Ni, J.; Faeflen, S.J.; Huang, X.; Wright, A.L. Manganese oxide affects nitrification and ammonia oxidizers in subtropical and temperate acid forest soils. Catena 2016, 137, 24–30. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Schloter, M.; Zhang, G.; Chen, Q.; Huang, J.; Li, L.; Elser, J.J.; Han, X. Response of the abundance of key soil microbial nitrogen-cycling genes to multi-factorial global changes. PLoS ONE 2013, 8, e76500. [Google Scholar] [CrossRef]
- Benini, S.; Rypniewski, W.R.; Wilson, K.S.; Ciurli, S.; Mangani, S. Structure-based rationalization of urease inhibition by phosphate: Novel insights into the enzyme mechanisms. J. Biol. Inorg. Chem. 2001, 6, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, B.; Zaborska, W. The effect of phosphate buffer in the range of pH 5.80-8.07 on jack bean urease activity. J. Mol. Catal. B Enzym. 1999, 6, 75–81. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, X.; Wang, X.; Shao, H.; Yang, J.; Wang, X. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, H.; Zhao, H.; Xia, H.; Sun, M.; Li, Z.; Li, P.; Zheng, C.; Dong, H.; Liu, J. Phosphorus affects enzymatic activity and chemical properties of cotton soil. Plant Soil Environ. 2019, 65, 361–368. [Google Scholar] [CrossRef]
- Wei, X.; Hu, Y.; Peng, P.; Zhu, Z.; Atere, C.T.; O’Donnell, A.G.; Wu, J.; Ge, T. Effect of P stoichiometry on the abundance of nitrogen-cycle genes in phosphorus-limited paddy soil. Soil Biol. Biochem. 2017, 53, 767–776. [Google Scholar] [CrossRef]
- Mori, T.; Ohta, S.; Ishizuka, S.; Konda, R.; Wicaksono, A.; Heriyanto, J.; Hardjono, A. Effects of phosphorus addition on N2O and NO emissions from soils of an Acacia mangium plantation. Soil Sci. Plant Nutr. 2010, 56, 782–788. [Google Scholar] [CrossRef]
- Fisk, M.; Ratliff, T.; Goswami, S.; Yanai, R. Synergistic soil response to nitrogen plus phosphorus fertilization in hardwood forests. Biogeochemistry 2014, 118, 195–204. [Google Scholar] [CrossRef]
- Wang, F.; Li, J.; Wang, X.; Zhang, W.; Zou, B.; Neher, D.A.; Li, Z. Nitrogen and phosphorus addition impact soil N2O emission in a secondary tropical forest of south China. Sci. Rep. 2014, 4, 5615. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huo, Y.; Guo, D.; Wang, Q.; Bao, Y.; Li, L. Effects of multi-nutrient additions on GHG fluxes in a temperate grassland of northern China. Ecosystems 2014, 17, 657–672. [Google Scholar] [CrossRef]
- White, J.R.; Reddy, K.R. Nitrification and denitrification rates of everglades wetland soils along a phosphorus-impacted gradient. J. Environ. Qual. 2003, 32, 2436–2443. [Google Scholar] [CrossRef]
- Chu, H.; Fujii, T.; Morimoto, S.; Lin, X.; Yagi, K. Population size and specific nitrification potential of soil ammonia-oxidizing bacteria under long-term fertilizer management. Soil Biol. Biochem. 2008, 40, 1960–1963. [Google Scholar] [CrossRef]
- Xue, C.; Penton, C.; Zhang, B.; Zhao, M.; Rothstein, D.E.; Mladenoff, D.J.; Forrester, J.A.; Shen, Q.; Tiedje, A.M. Soil fungal and bacterial responses to conversion of open land to short rotation woody biomass crops. GCB Bioenergy 2015, 8, 723–736. [Google Scholar] [CrossRef]
- He, J.Z.; Shen, J.P.; Zhang, L.M.; Zhu, Y.G.; Zheng, Y.M.; Xu, M.G.; Di, H.J. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environ. Microbiol. 2007, 9, 2364–2374. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.M.; Shen, J.P.; Wei, W.X.; He, J.Z. Abundance and community structure of ammonia-oxidizing archaea and bacteria in an acid paddy soil. Biol. Fertil. Soils 2011, 47, 323–331. [Google Scholar] [CrossRef]
- Yang, X.; Ni, K.; Shi, Y.; Yi, X.; Ji, L.; Ma, L.; Ruan, J. Heavy nitrogen application increases soil nitrification through ammoniaoxidizing bacteria rather than archaea in acidic tea (Camellia sinensis L.) plantation soil. Sci. Total Environ. 2020, 717, 137248. [Google Scholar] [CrossRef]
- He, X.; Li, S.; Wu, F. Responses of Ammonia-oxidizing microorganisms to intercropping systems in different seasons. Agriculture 2021, 11, 195. [Google Scholar] [CrossRef]
- Dong, J.; Che, R.; Jia, S.; Wang, F.; Zhang, B.; Cui, X.; Wang, S.; Wang, S. Responses of ammonia-oxidizing archaea and bacteria to nitrogen and phosphorus amendments in an alpine steppe. Eur. J. Soil Sci. 2020, 71, 94–954. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicol, G.W. Relative contributions of archaea and bacteria to aerobic ammonia oxidation in the environment. Environ. Microbiol. 2008, 10, 2931–2941. [Google Scholar] [CrossRef] [PubMed]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Song, H.; Che, Z.; Cao, W.; Huang, T.; Wang, J.; Dong, Z. Changing roles of ammonia-oxidizing bacteria and archaea in a continuously acidifying soil caused by over-fertilization with nitrogen. Environ. Sci. Pollut. Res. 2016, 23, 11964–11974. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | NO3−-N (mg kg−1d−1) at Different Incubation Time (Days) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 5 | 7 | 10 | |
| C | 6.84 ± 1.01 a | 6.07 ± 1.30 a | 3.08 ± 1.35 b | 5.20 ± 1.12 a | 3.75 ± 0.13 a | 2.45 ± 0.35 a |
| P | 6.05 ± 1.28 a | 5.21 ± 0.46 a | 5.18 ± 0.55 a | 6.63 ± 0.20 a | 2.70 ± 0.29 b | 3.28 ± 0.82 a |
| Urea | 6.04 ± 2.86 a | 12.12 ± 1.72 a | 20.63 ± 2.06 a | 27.84 ± 1.09 a | 17.86 ±0.50 a | 14.39 ± 1.37 a |
| Urea + P | 4.15 ± 0.81 a | 3.38 ± 0.24 b | 8.10 ± 1.63 b | 20.38 ± 0.70 b | 18.03 ± 2.35 a | 14.05 ± 0.30 a |
| Treatments | Np (mg N kg−1) | k1 (day−1) | R2 | Vp (mg N kg−1 day−1) |
|---|---|---|---|---|
| C | 40.15 ± 0.85 | 0.26 ± 0.03 | 0.89 | 7.63 ± 0.83 |
| P | 39.41 ± 2.16 | 0.23 ± 0.17 | 0.77 | 8.17 ± 3.35 |
| Urea | 203.8 ± 18.78 | 0.18 ± 0.02 | 0.91 | 36.18 ± 1.22 |
| Urea + P | 379.91 ± 19.49 | 0.06 ± 0.01 | 0.92 | 22.19 ± 1.92 |
| Items | Treatments | Incubation Time (Days) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 5 | 7 | 10 | ||
| NH4+-N/AP | C | 0.25 ± 0.04 a | 0.21 ± 0.02 a | 0.12 ± 0.04 a | 0.11 ± 0.01 a | 0.07 ± 0.01 a | 0.06 ± 0.01 a | 0.04 ± 0.01 a |
| P | 0.12 ± 0.01 b | 0.11 ± 0.01 b | 0.07 ± 0.01 b | 0.09 ± 0.01 b | 0.07 ± 0.02 a | 0.02 ± 0.01 b | 0.02 ± 0.01 b | |
| Urea | 0.30 ± 0.06 a | 0.98 ± 0.12 a | 1.17 ± 0.10 a | 1.31 ± 0.04 a | 1.03 ± 0.10 a | 0.80 ± 0.09 a | 0.52 ± 0.13 a | |
| Urea + P | 0.17 ± 0.03 b | 0.46 ± 0.01 b | 0.65 ± 0.02 b | 0.75 ± 0.04 b | 0.75 ± 0.03 b | 0.66 ± 0.07 a | 0.45 ± 0.1 1 a | |
| NO3−-N/AP | C | 0.34 ± 0.04 a | 0.47 ± 0.01 a | 0.53 ± 0.09 a | 0.55 ± 0.11 a | 0.60 ± 0.10 a | 0.65 ± 0.05 a | 0.62 ± 0.02 a |
| P | 0.23 ± 0.01 b | 0.23 ± 0.02 b | 0.24 ± 0.02 b | 0.26 ± 0.01 b | 0.36 ± 0.01 b | 0.39 ± 0.03 b | 0.39 ± 0.06 b | |
| Urea | 0.40 ± 0.03 a | 0.47 ± 0.02 a | 0.59 ± 0.04 a | 0.91 ± 0.09 a | 1.45 ± 0.17 a | 1.32 ± 0.11 a | 1.43 ± 0.12 a | |
| Urea + P | 0.22 ± 0.01 b | 0.23 ± 0.01 b | 0.25 ± 0.01 b | 0.33 ± 0.02 b | 0.74 ± 0.02 b | 0.85 ± 0.09 b | 1.00 ± 0.06 b | |
| Variable | pH | AP | NH4+-N | NO3−-N | AOB | AOA | AOB/AOA | NR | NH4+-N/AP | NO3−-N/AP |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1 | |||||||||
| AP | −0.160 | 1 | ||||||||
| NH4+-N | 0.217 * | 0.121 | 1 | |||||||
| NO3−-N | −0.476 ** | 0.016 | 0.506 ** | 1 | ||||||
| AOB | −0.194 | −0.120 | 0.763 ** | 0.831 ** | 1 | |||||
| AOA | 0.288 | −0.273 | 0.215 | −0.062 | 0.380 * | 1 | ||||
| AOB/AOA | −0.399 * | 0.034 | 0.612 ** | 0.963 ** | 0.769 ** | −0.166 | 1 | |||
| NR | −0.195 | −0.033 | 0.686 ** | 0.838 ** | 0.963 ** | 0.493 * | 0.781 ** | 1 | ||
| NH4+-N/AP | 0.232 * | −0.137 | 0.934 ** | 0.477 ** | 0.870 ** | 0.424 * | 0.589 ** | 0.708 ** | 1 | |
| NO3−-N/AP | −0.362 * | −0.376 ** | 0.412 ** | 0.900 ** | 0.822 ** | 0.080 | 0.850 ** | 0.788 ** | 0.507 * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, J.; Arai, Y.; Shen, J.; Wang, R.; Ai, S. Effects of Phosphorus on Nitrification Process in a Fertile Soil Amended with Urea. Agriculture 2021, 11, 523. https://doi.org/10.3390/agriculture11060523
Ning J, Arai Y, Shen J, Wang R, Ai S. Effects of Phosphorus on Nitrification Process in a Fertile Soil Amended with Urea. Agriculture. 2021; 11(6):523. https://doi.org/10.3390/agriculture11060523
Chicago/Turabian StyleNing, Jianfeng, Yuji Arai, Jian Shen, Ronghui Wang, and Shaoying Ai. 2021. "Effects of Phosphorus on Nitrification Process in a Fertile Soil Amended with Urea" Agriculture 11, no. 6: 523. https://doi.org/10.3390/agriculture11060523
APA StyleNing, J., Arai, Y., Shen, J., Wang, R., & Ai, S. (2021). Effects of Phosphorus on Nitrification Process in a Fertile Soil Amended with Urea. Agriculture, 11(6), 523. https://doi.org/10.3390/agriculture11060523