
Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau

Abstract
:1. Introduction
2. Materials and Methods
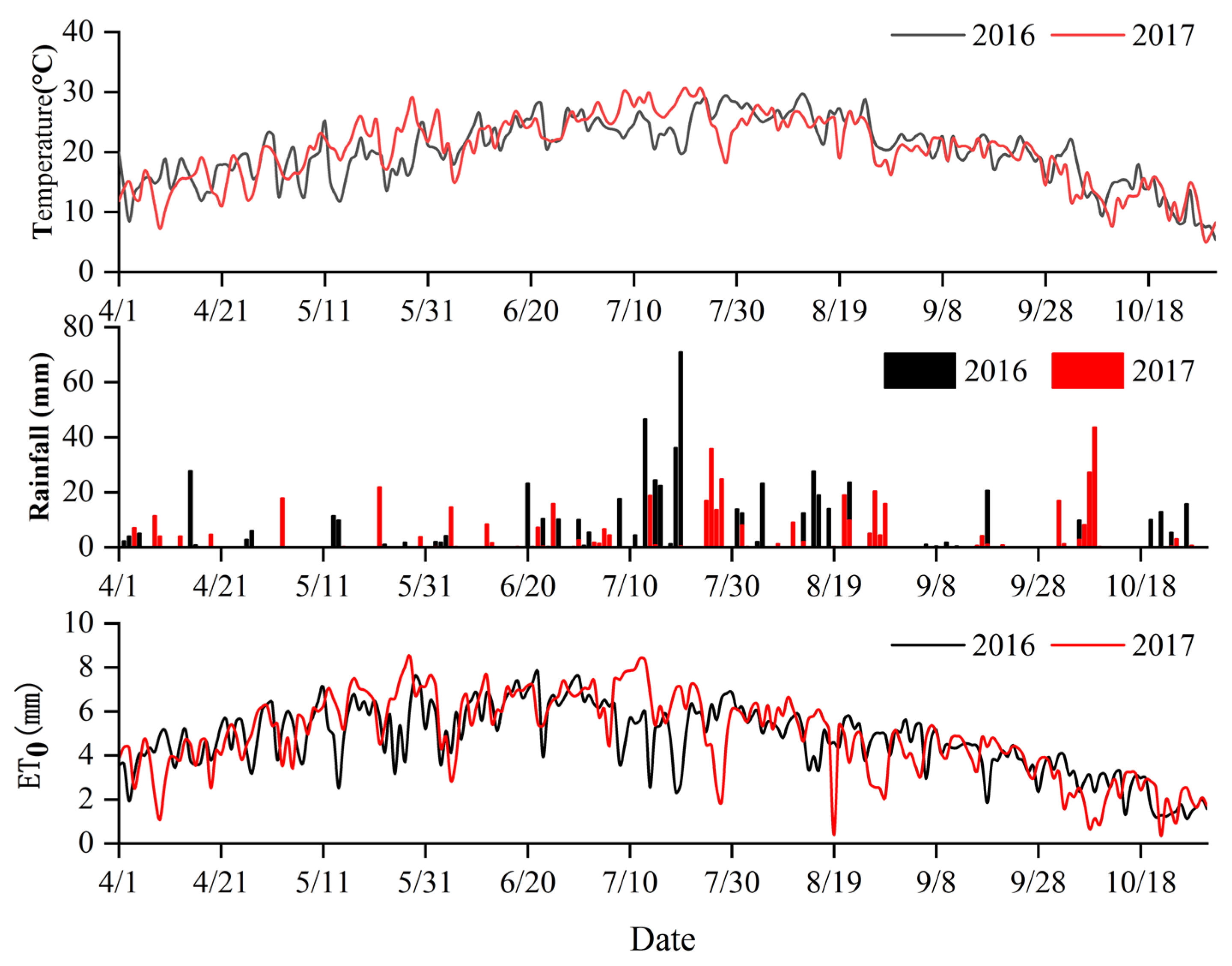
2.1. Study Site
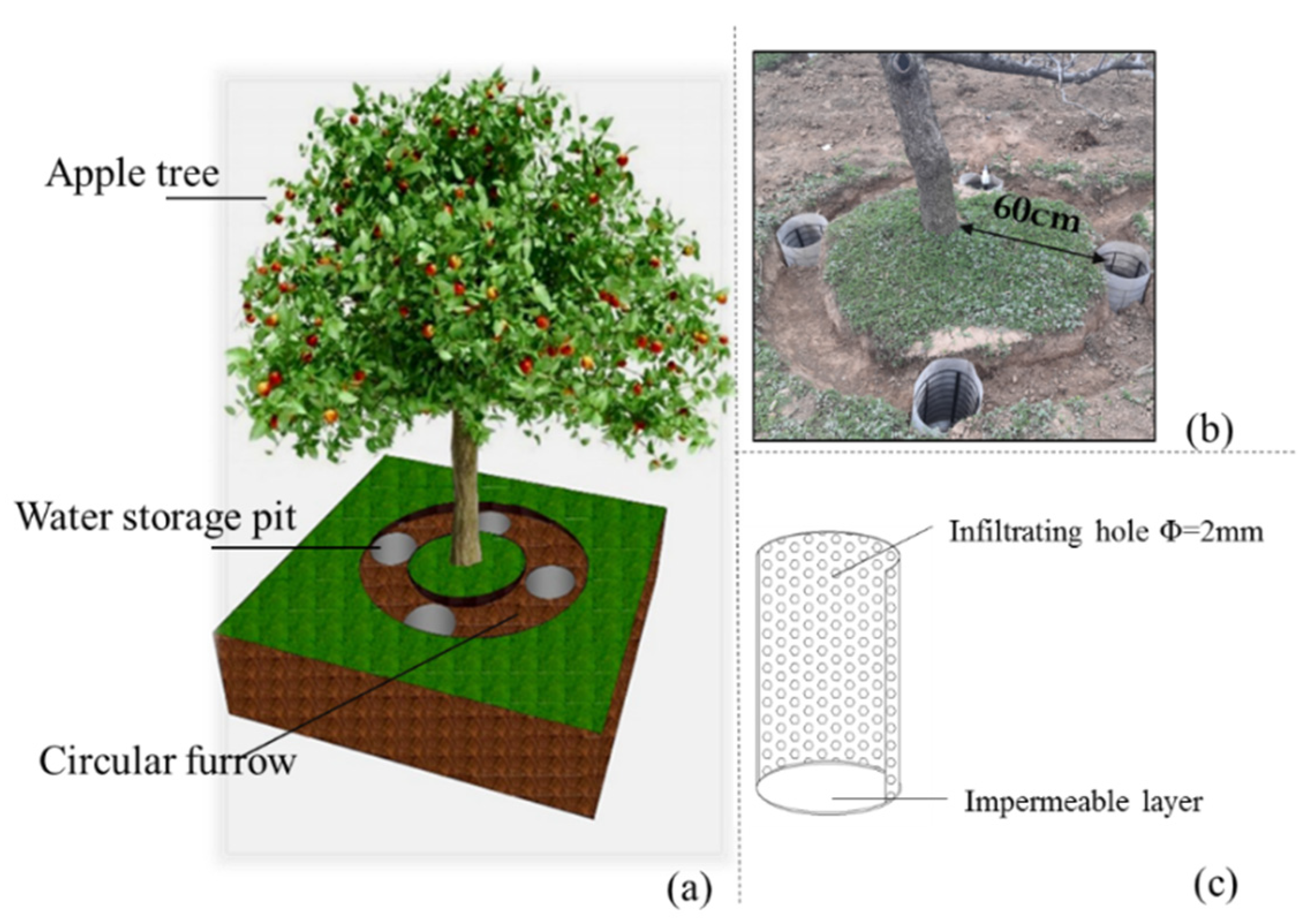
2.2. Experimental Design
2.3. Data Collection
2.3.1. Gas Exchange Parameters, Leaf Water Potential, and Soil Water Content
2.3.2. Leaf Chlorophyll Content and Chlorophyll Fluorescence Features
2.3.3. Antioxidant Enzyme and Soluble Sugar Content
2.4. Data Analysis
3. Results
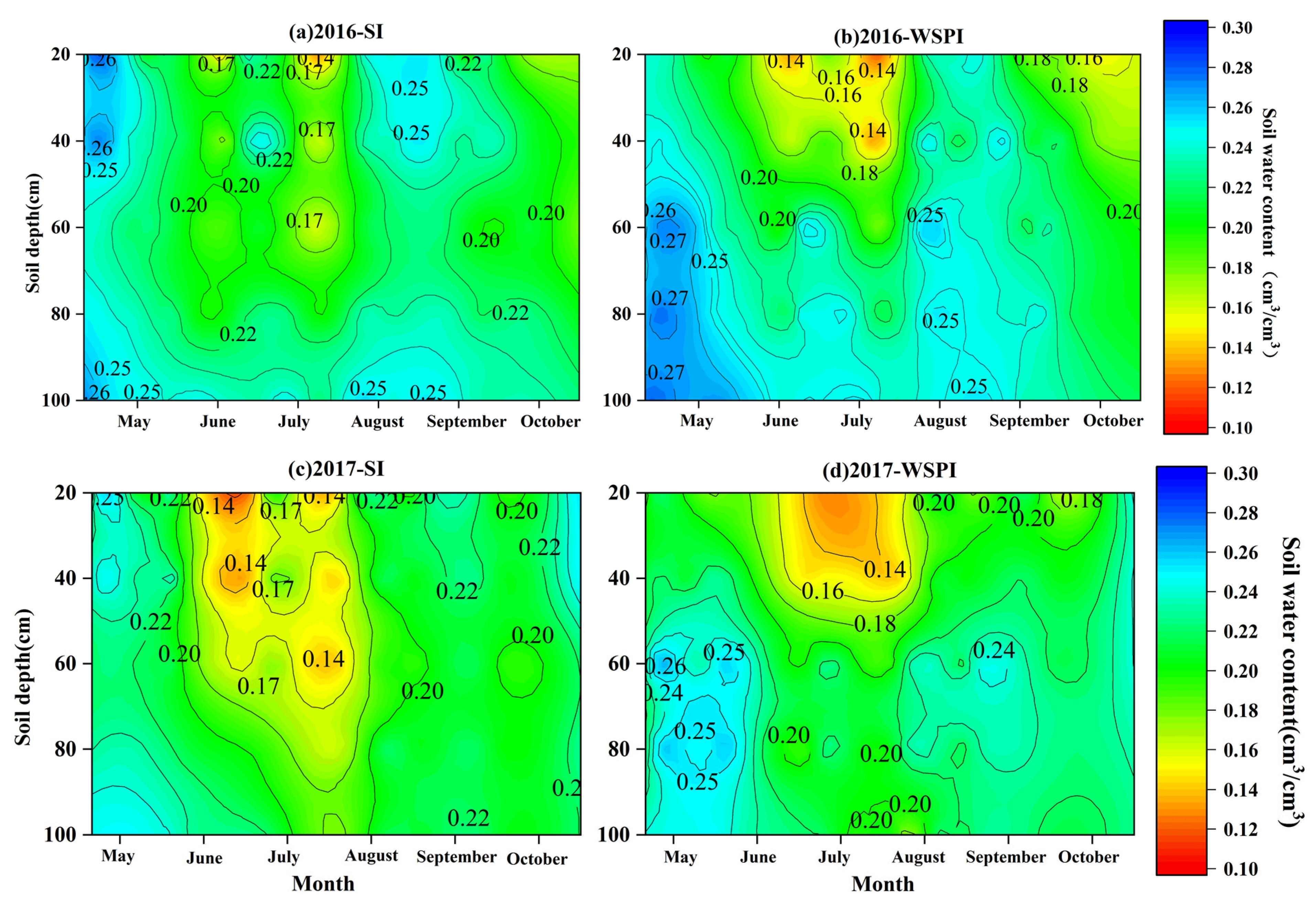
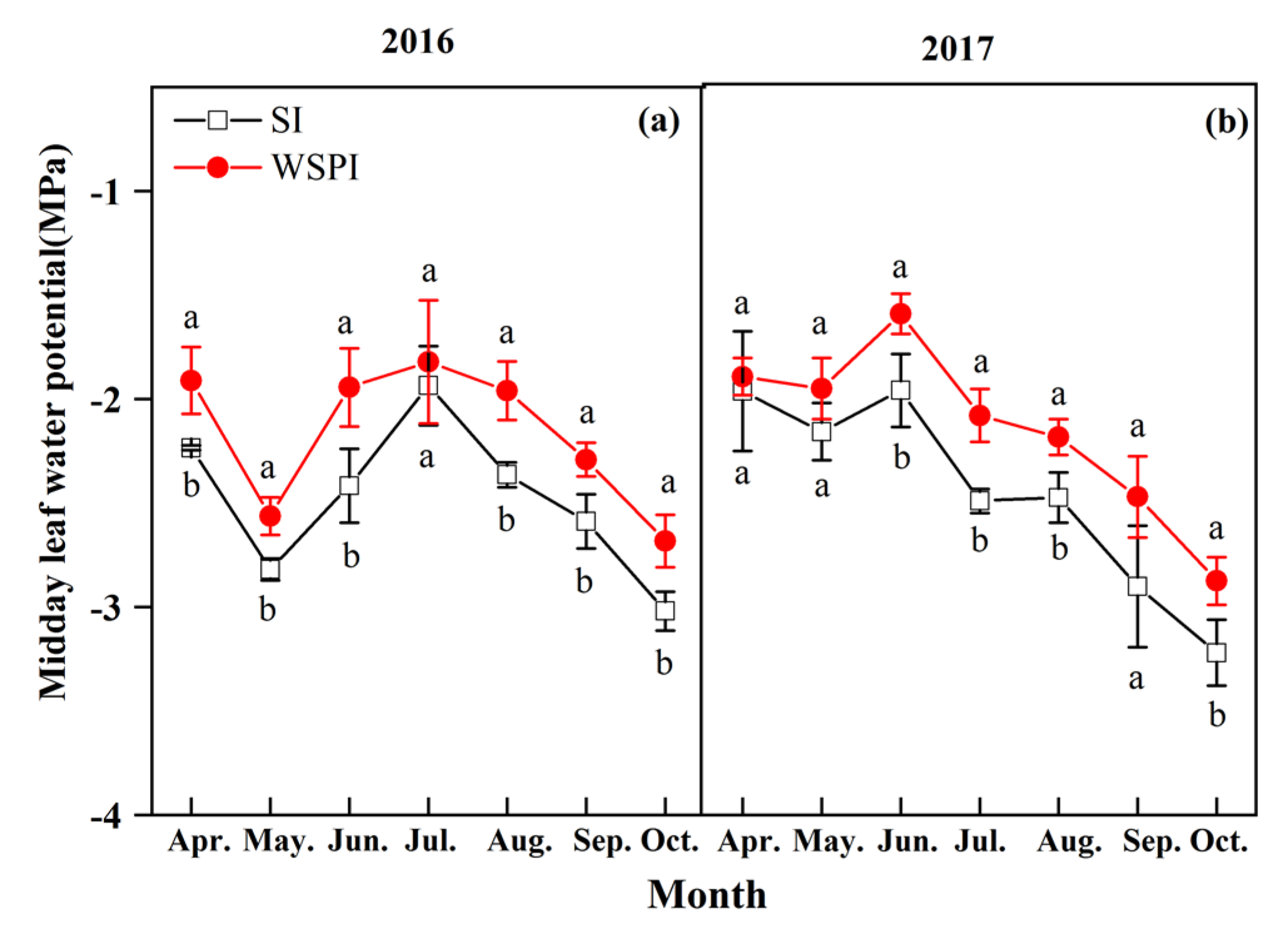
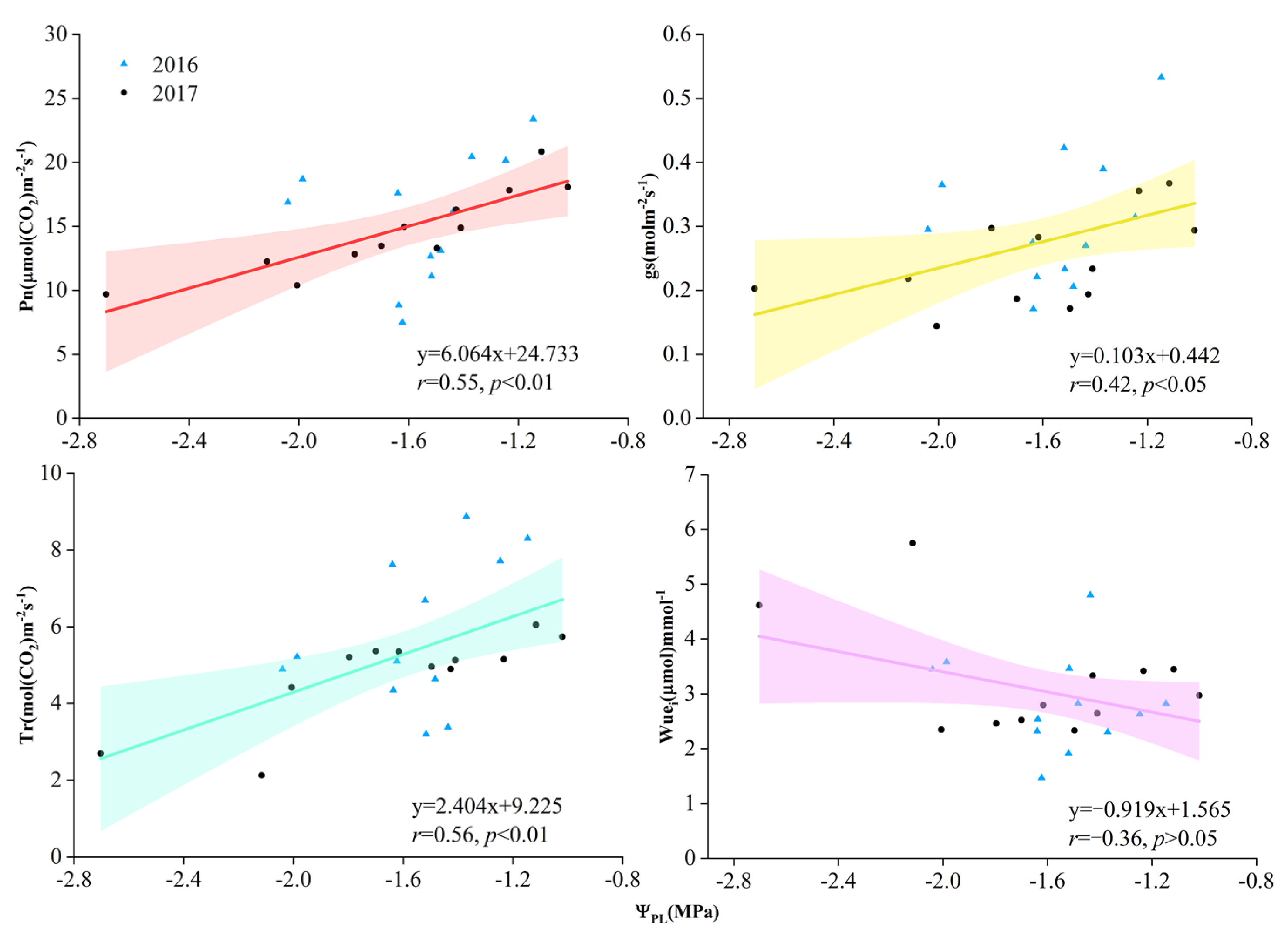
3.1. Soil Water Content and Leaf Water Potential
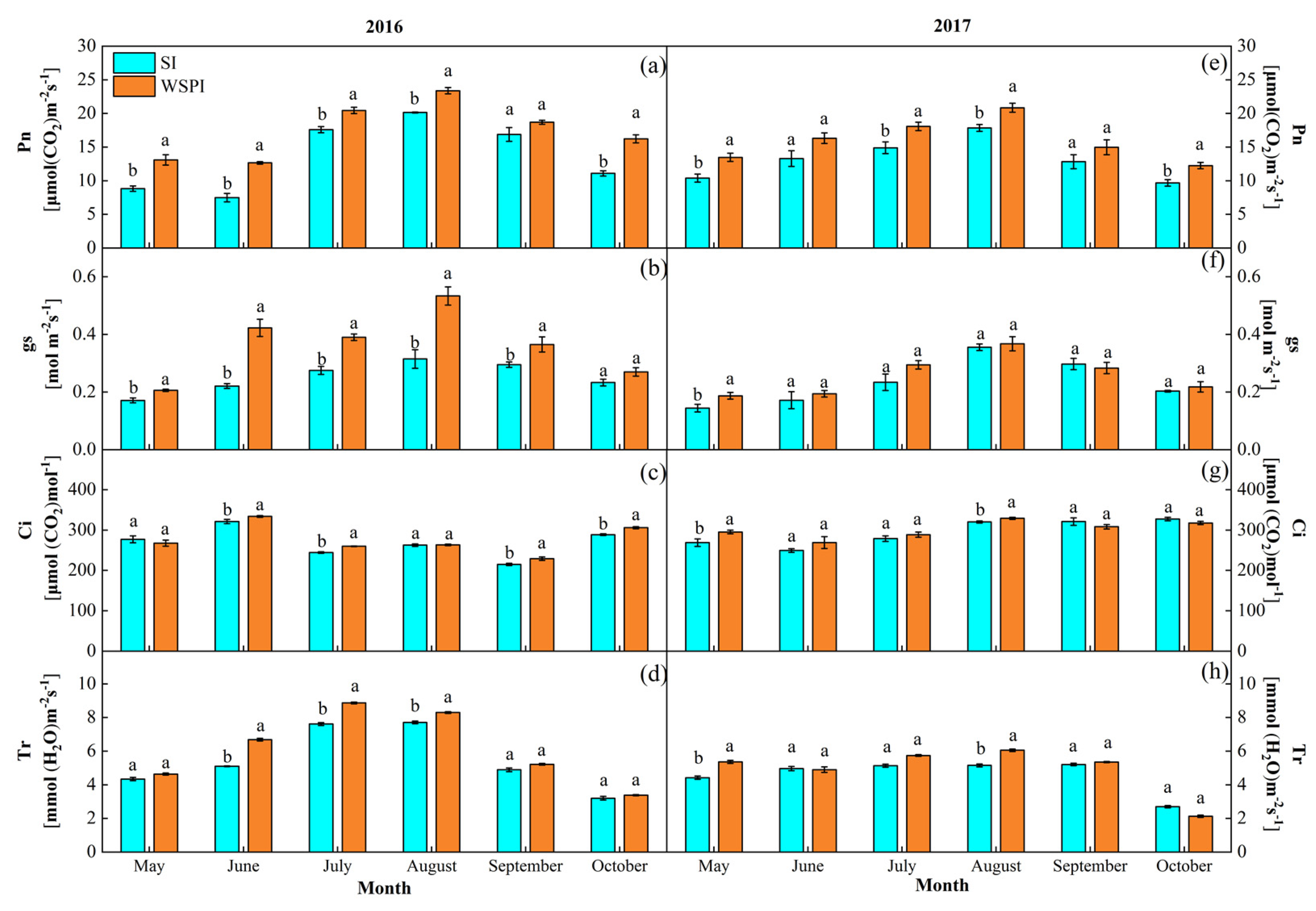
3.2. Gas Exchange Characteristics
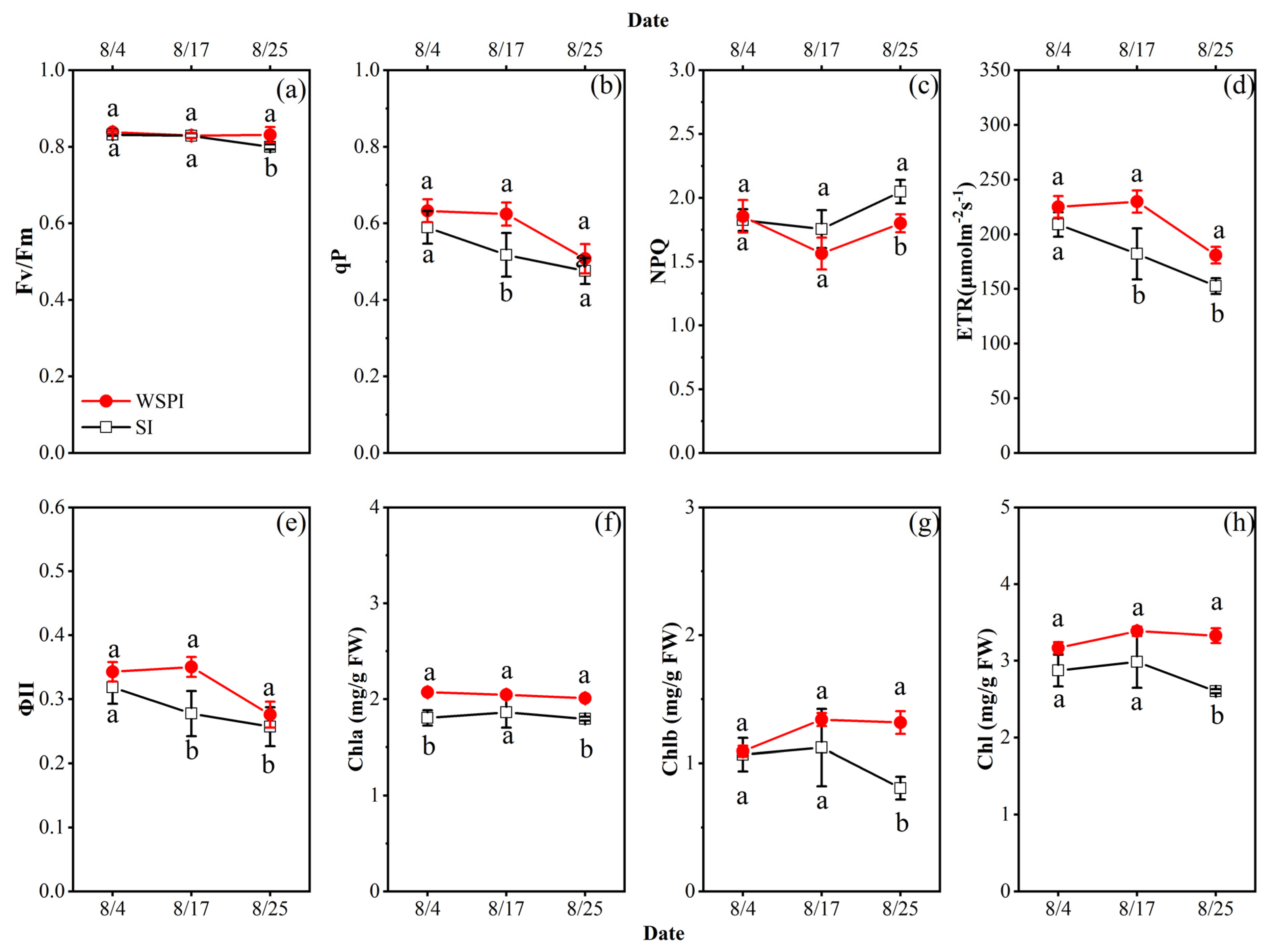
3.3. Chlorophyll Content and Fluorescence Features of Leaves
3.4. Antioxidant Enzyme, Soluble Sugar, and Yield
3.5. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kviklys, D.; Samupliene, G. Relationships among the rootstock, crop load, and sugar hormone signaling of apple tree, and their effects on biennial bearing. Front. Plant Sci. 2020, 11, 1213. [Google Scholar] [CrossRef]
- Zhao, D.; Yuan, J.; Xu, K.; Cheng, C.; Li, H. Selection of morphological, physiological and biochemical indices: Evaluating dwarfing apple interstocks in cold climate zones. N. Z. J. Crop Hortic. Sci. 2016, 44, 291–311. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Zhao, X.; Wu, P.; Dyck, M. Spatial distribution of soil moisture and fine roots in rain-fed apple orchards employing a rainwater collection and infiltration (rwci) system on the loess plateau of china. Agric. Water Manag. 2017, 184, 170–177. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, J.; Sun, X.; Guo, X.; Cheng, Q.; Shi, X. Estimating the root water uptake of surface-irrigated apples using water stable isotopes and the hydrus-1D model. Water 2018, 10, 1624. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, X.; Wu, P.; Yang, M.; Ye, M.; Tian, L.; Zou, Y.; Wu, Y.; Zhang, F.; Siddique, K. The economic-environmental trade-off of growing apple trees in the drylands of China: A conceptual framework for sustainable intensification. J. Clean. Prod. 2021, 296, 126497. [Google Scholar] [CrossRef]
- Guo, X.; Lei, T.; Sun, X.; Ma, J.; Zheng, L.; Zhang, S.; He, Q. Modelling soil water dynamics and root water uptake for apple trees under water storage pit irrigation. Int. J. Agric. Biol. Eng. 2019, 12, 126–134. [Google Scholar] [CrossRef]
- Meng, W.; Sun, X.; Ma, J.; Guo, X.; Lei, T.; Li, R. Measurement and simulation of the water storage pit irrigation trees evapotranspiration in the Loess Plateau. Agric. Water Manag. 2020, 226, 105804. [Google Scholar] [CrossRef]
- Sun, X.; Ma, J.; Guo, X. Water storage pit irrigation technique. In Proceedings of the International Conference on Effective Utilization of Agricultural Soil & Water Resources and Protection of Environment and the 4th Annual Academic Conference of Chinese Society of Agricultural Soil and Water Engineering, Nanjing, China, 16–18 August 2006; Hohai University Press: Nanjing, China, 2007; pp. 211–214. [Google Scholar]
- Sun, X. Effect of water storage pit irrigation on soil and water conservation. In Proceedings of the 12th ISCO Conference, Beijing, China, 26–31 May 2002; Tsinghua University Press: Beijing, China, 2002; Volume 2, pp. 367–369. [Google Scholar]
- Li, T.; Zhang, J. Effect of pit irrigation on soil water content, vigor, and water use efficiency within vineyards in extremely arid regions. Sci. Hortic. 2017, 218, 30–37. [Google Scholar] [CrossRef]
- Nazari, E.; Besharat, S.; Zeinalzadeh, K.; Mohammadi, A. Measurement and simulation of the water flow and root uptake in soil under subsurface drip irrigation of apple tree. Agric. Water Manag. 2021, 255, 106972. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, Y.; Hill, R.; Wu, S.; Song, X. Combined effects of biomaterial amendments and rainwater harvesting on soil moisture, structure and apple roots in a rainfed apple orchard on the Loess Plateau, China. Agric. Water Manag. 2021, 248, 106776. [Google Scholar] [CrossRef]
- Cheng, Q.; Ma, J.; Ren, R.; Zheng, L.; Guo, X.; Sun, X. Effects of fertilization management under WSPI on soil nitrogen distribution and nitrogen absorption in apple orchard in Loess Plateau. Agronomy 2020, 10, 1386. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, J.; Sun, X.; Guo, X. Spatial distribution of soil moisture and fine roots of apple trees under water storage pit irrigation. J. Irrig. Drain. Eng. 2014, 140, 333–340. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Chen, Z.; Tao, X.; Khan, A.; Tan, D.; Luo, H. Biomass accumulation, photosynthetic traits and root development of cotton as affected by irrigation and nitrogen-fertilization. Front. Plant Sci. 2018, 9, 173. [Google Scholar] [CrossRef]
- Kucukyumuk, C.; Kacal, E.; Ertek, A.; Ozturk, G.; Kurttas, Y.S.K. Pomological and vegetative changes during transition from flood irrigation to drip irrigation: Starkrimson Delicious apple variety. Sci. Hortic. 2012, 136, 17–23. [Google Scholar] [CrossRef]
- Ghafari, H.; Hassanpour, H.; Jafari, M.; Besharat, S. Effect of partial root zone irrigation on physiology, water use efficiency, fruit yield, phenolic compounds and antioxidant capacity of apple. Arch. Agron. Soil Sci. 2020, 67, 1521–1538. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, G.; Gu, S.; Xia, J.; Zhao, J. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica 2010, 48, 589–595. [Google Scholar] [CrossRef]
- Sircelj, H.; Tausz, M.; Grill, D.; Batič, F. Detecting different levels of drought stress in apple trees (Malus domestica, Borkh.) with selected biochemical and physiological parameters. Sci. Hortic. 2007, 113, 362–369. [Google Scholar] [CrossRef]
- Fujisawa, H.; Moriya, Y. Influence of rootstock and soil moisture on leaf photosynthesis of apple trees on JM1, JM7, JM8, and M.9 rootstocks. Acta Hortic. 2012, 932, 441–446. [Google Scholar] [CrossRef]
- Bolat, I.; Dikilitas, M.; Ercisli, S.; Ikinci, A.; Tonkaz, T. The effect of water stress on some morphological, physiological, and biochemical characteristics and bud success on apple and quince rootstocks. Sci. World J. 2014, 2014, 769732. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G.; Glenn, D. Apple rootstock resistance to drought. Sci. Hortic. 2016, 204, 70–78. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.; Yoon, T. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Zegbe, J.; Behboudian, M.; Clothier, B. Reduced irrigation maintains photosynthesis, growth, yield, and fruit quality in ‘pacific rose’ apple. J. Sustain. Agric. 2007, 30, 125–136. [Google Scholar] [CrossRef]
- Guo, X.; Sun, X.; Ma, J.; Lei, T.; Zheng, L. Effects of water storage pit irrigation on root distribution and root distribution fuction of apple trees. Fresen. Environ. Bull. 2019, 28, 326–338. [Google Scholar]
- Zheng, L.; Ma, J.; Sun, X.; Guo, X.; Li, Y.; Ren, R.; Cheng, Q. Effective root growth zone of apple tree under water storage pit irrigation using stable isotope methodology. Arch. Agron. Soil Sci. 2019, 65, 1521–1535. [Google Scholar] [CrossRef]
- Rouhi, V.; Samson, R.; Lemeur, R.; Van Damme, P. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ. Exp. Bot. 2007, 59, 117–129. [Google Scholar] [CrossRef]
- Glenn, D. Canopy gas exchange and water use efficiency of ’Empire’ apple in response to particle Film, irrigation, and microclimatic factors. J. Am. Soc. Hortic. Sci. 2010, 135, 25–32. [Google Scholar] [CrossRef]
- Jaouhari, N.; Abouabdillah, A.; Bouabid, R.; Bourioug, M.; Aleya, L.; Chaoui, M. Assessment of sustainable deficit irrigation in a moroccan apple orchard as a climate change adaptation strategy. Sci. Total Environ. 2018, 642, 574–581. [Google Scholar] [CrossRef]
- Porra, R.; Thompson, W.; Kriedmann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Tian, J.; Hu, Y.; Gan, X.; Zhang, Y.; Hu, X.; Gou, L.; Luo, H.; Zhang, W. Effects of increased night temperature on cellulose synthesis and the activity of sucrose metabolism enzymes in cotton fiber. J. Integr. Agric. 2013, 12, 979–988. [Google Scholar] [CrossRef]
- Dhindsa, R.; Plumb-Dhindsa, P.; Thorpe, T. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Williams, L.; Araujo, F. Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in vitis vinifera. J. Am. Soc. Hortic. 2002, 127, 448–454. [Google Scholar] [CrossRef]
- Zhou, B.; Sun, J.; Liu, S.; Jin, W.; Zhang, Q.; Wei, Q. Dwarfing apple rootstock responses to elevated temperatures: A study on plant physiological features and transcription level of related genes. J. Integr. Agric. 2016, 15, 1025–1033. [Google Scholar] [CrossRef]
- Atkinson, C.; Policarpo, M.; Webster, A.; Kingswell, G. Drought tolerance of clonal malus determined from measurements of stomatal conductance and leaf water potential. Tree Physiol. 2000, 20, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Naor, A.; Gal, Y.; Peres, M. The inherent variability of water stress indicators in apple, nectarine and pear orchards, and the validity of a leaf-selection procedure for water potential measurements. Irrig. Sci. 2006, 24, 129–135. [Google Scholar] [CrossRef]
- O’Connell, M.; Goodwin, I. Responses of ‘pink lady’ apple to deficit irrigation and partial rootzone drying: Physiology, growth, yield, and fruit quality. Aust. J. Agric. Res. 2007, 58, 1068–1076. [Google Scholar] [CrossRef]
- Eyles, A.; Bound, S.; Oliver, G.; Corkrey, R.; Hardie, M.; Green, S.; Close, D. Impact of biochar amendment on the growth, physiology and fruit of a young commercial apple orchard. Trees 2015, 29, 1817–1826. [Google Scholar] [CrossRef]
- Sircelj, H.; Tausz, M.; Grill, D.; Batic, F. Biochemical responses in leaves of two apple tree cultivars subjected to progressing drought. J. Plant Physiol. 2005, 162, 1308–1318. [Google Scholar] [CrossRef]
- Egea, G.; Dodd, I.; Gonzálezreal, M.; Domingo, R.; Baille, A. Partial rootzone drying improves almond tree leaf-level water use efficiency and afternoon water status compared with regulated deficit irrigation. Funct. Plant Biol. 2011, 38, 372–385. [Google Scholar] [CrossRef]
- Guo, Z.; Yu, Z.; Wang, D.; Shi, Y.; Zhang, Y. Photosynthesis and winter wheat yield responses to supplemental irrigation based on measurement of water content in various soil layers. Field Crop. Res. 2014, 166, 102–111. [Google Scholar] [CrossRef]
- Li, H.; Si, B.; Ma, X.; Wu, P. Deep soil water extraction by apple sequesters organic carbon via root biomass rather than altering soil organic carbon content. Sci. Total Environ. 2019, 670, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Black, M.; Patterson, K.; Minchin, P.; Gould, K.; Clearwater, M. Hydraulic responses of whole vines and individual roots of kiwifruit (actinidia chinensis) following root severance. Tree Physiol. 2011, 31, 508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, X.; Ma, J.; Guo, X.; An, J.; Li, R. Daily variation of stem flow rate of apple tree and influence factors under water storage pit irrigation. Water Saving Irri. 2017, 5, 11–15. [Google Scholar]
- Solari, L.; Johnson, S.; Dejong, T. Relationship of water status to vegetative growth and leaf gas exchange of peach (prunus persica) trees on different rootstocks. Tree Physiol. 2006, 26, 1333–1341. [Google Scholar] [CrossRef]
- Sun, X.; Yan, H.; Ma, P.; Liu, B.; Zou, Y.; Liang, D.; Ma, F.; Li, P. Responses of young ‘pink lady’ apple to alternate deficit irrigation following long-term drought: Growth, photosynthetic capacity, water-use efficiency, and sap flow. Photosynthetica 2012, 50, 501–507. [Google Scholar] [CrossRef]
- Ma, X.; Jacoby, P.; Sanguinet, K. Improving net photosynthetic rate and rooting depth of grapevines through a novel irrigation strategy in a semi-arid climate. Front. Plant Sci. 2020, 11, 575303. [Google Scholar] [CrossRef]
- Hamed, S.; Lefi, E.; Chaieb, M. Physiological responses of pistacia vera, l. versus pistacia atlantica desf.to water stress conditions under arid bioclimate in tunisia. Sci. Hortic. 2016, 203, 224–230. [Google Scholar] [CrossRef]
- Farquhar, G.; Ehleringer, J.; Hubick, K. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Phys. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Garcia-Sanchez, F.; Syvertsen, J.; Gimeno, V.; Botia, P.; Perez-Perez, J. Responses to flooding and drought stress by two citrus rootstock seedlings with different water-use efficiency. Physiol. Plant. 2007, 130, 532–542. [Google Scholar] [CrossRef]
- Yuan, J.; Xu, M.; Duan, W.; Fan, P.; Li, S. Effects of whole-root and half-root water stress on gas exchange and chlorophyll fluorescence parameters in apple trees. J. Am. Soc. Hortic. Sci. 2013, 138, 395–402. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [PubMed]
- Sayyad-Amin, P.; Jahansooz, M.; Borzouei, A.; Ajili, F. Changes in photosynthetic pigments and chlorophyll-a fluorescence attributes of sweet-forage and grain sorghum cultivars under salt stress. J. Biol. Phys. 2016, 42, 601–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, S.; Ribeiro, C.; Bacelar, E.; Ferreira, H.; Oliveira, I.; Silva, A.; Goncalves, B. Influence of training system on physiological performance, biochemical composition and antioxidant parameters in apple tree (Malus domestica Borkh.). Sci. Hortic. 2017, 225, 394–398. [Google Scholar] [CrossRef]
- Li, J.; Chen, P.; Sun, X.; Liu, P. Effects of fertigation strategies on water and nitrogen distribution under water storage pit irrigation for orchards. Int. J. Agric. Biol. Eng. 2018, 11, 165–171. [Google Scholar] [CrossRef]
- Baker, N. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef]
- Ballester, C.; Zarco-Tejada, P.; Nicolas, E.; Alarcon, J.; Fereres, E.; Intrigliolo, D.; Gonzalez-Dugo, V. Evaluating the performance of xanthophyll, chlorophyll and structure-sensitive spectral indices to detect water stress in five fruit tree species. Precis. Agric. 2018, 19, 178–193. [Google Scholar] [CrossRef]
- Petridis, A.; Therios, I.; Samouris, G.; Koundouras, S.; Giannakoula, A. Effect of water deficit on leaf phenolic composition, gas exchange, oxidative damage and antioxidant activity of four greek olive (olea europaea L.) cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef]
- Abid, G.; M’hamdi, M.; Mingeot, D.; Aouida, M.; Aroua, I.; Muhovski, Y.; Khaled, S.; Souissi, F.; Mannai, K.; Jebara, M. Effect of drought stress on chlorophyll fluorescence, antioxidant enzyme activities and gene expression patterns in faba bean (Vicia faba L.). Arch. Agron. Soil Sci. 2017, 63, 536–552. [Google Scholar] [CrossRef]
- Silva, E.; Ferreira-Silva, S.; Fontenele, A.; Ribeiro, R.; Viégas, R.; Silveira, J. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef]
- Pina, J.; Moraes, R. Gas exchange, antioxidants and foliar injuries in saplings of a tropical woody species exposed to ozone. Ecotoxicol. Environ. Saf. 2010, 73, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef] [Green Version]
- Samuoliene, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between apple tree rootstock, crop-load, plant nutritional status and yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Hernandez-Santana, V.; Fernández, J.; Cuevas, M.; Perez-Martin, A.; Diaz-Espejo, A. Photosynthetic limitations by water deficit: Effect on fruit and olive oil yield, leaf area and trunk diameter and its potential use to control vegetative growth of super-highdensity olive orchards. Agric. Water Manag. 2017, 184, 9–18. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, F.; Li, F. Effect of different drip irrigation methods and fertilization on growth, physiology and water use of young apple tree. Sci. Hortic. 2011, 129, 119–126. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Month | Rs (W m−1) | RH (%) | Ta (℃) | P (mm) | VPD (kPa) | ET0 (mm) | I (mm) |
|---|---|---|---|---|---|---|---|---|
| April | 239.55 | 55.52 | 16.26 | 48.6 | 0.79 | 130.78 | 45 | |
| May | 234.67 | 59.85 | 18.17 | 24.0 | 0.81 | 168.79 | ||
| June | 265.69 | 73.35 | 23.19 | 76.0 | 0.71 | 187.17 | 45 | |
| 2016 | July | 233.92 | 87.91 | 25.05 | 262.8 | 0.35 | 170.89 | |
| August | 240.54 | 87.64 | 25.24 | 134.2 | 0.35 | 155.29 | ||
| September | 224.33 | 82.37 | 20.18 | 24.2 | 0.39 | 123.81 | 45 | |
| October | 155.75 | 83.39 | 12.95 | 58.5 | 0.21 | 74.19 | ||
| April | 237.13 | 52.63 | 14.69 | 31 | 0.80 | 124.57 | 45 | |
| May | 258.58 | 49.91 | 21.25 | 43.6 | 1.18 | 191.65 | 45 | |
| June | 241.54 | 72.87 | 23.07 | 51.6 | 0.72 | 187.47 | 45 | |
| 2017 | July | 228.24 | 79.65 | 26.75 | 125.4 | 0.67 | 194.26 | 45 |
| August | 214.65 | 88.24 | 23.91 | 95 | 0.33 | 151.73 | 45 | |
| September | 199.51 | 81.01 | 20.29 | 6.6 | 0.42 | 125.15 | ||
| October | 106.32 | 91.23 | 12.08 | 104.2 | 0.12 | 65.83 |
| SOD (U/g FW) | Sucrose (mg/g DW) | Fructose (mg/g DW) | Yield (kg/tree) | Single Fruit Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| August | October | August | October | August | October | 2016 | 2017 | 2016 | 2017 | |
| SI | 150.24 a | 175.67 a | 11.34 a | 11.41 a | 0.78 a | 0.23 a | 10.28 b | 11.47 b | 183.72 b | 170.64 b |
| WSPI | 95.39 b | 123.56 b | 6.98 b | 11.71 a | 0.77 a | 0.27 a | 12.89 a | 14.53 a | 234.39 a | 213.03 a |
| Traits | PC1 | PC2 | PC3 |
|---|---|---|---|
| Ψm | 0.969 | 0.055 | 0.029 |
| Pn | 0.405 | 0.885 | −0.080 |
| gs | −0.354 | 0.869 | 0.212 |
| Ci | −0.708 | 0.476 | 0.313 |
| Tr | 0.574 | 0.629 | 0.415 |
| WUE | −0.849 | 0.038 | 0.291 |
| Fv/Fm | 0.714 | 0.559 | −0.123 |
| qP | 0.966 | 0.065 | 0.159 |
| NPQ | −0.131 | −0.295 | 0.882 |
| ETR | 0.942 | 0.225 | −0.109 |
| ΦII | 0.941 | 0.226 | −0.108 |
| Chla | −0.597 | 0.722 | −0.303 |
| Chlb | −0.498 | 0.782 | −0.148 |
| Chl | −0.554 | 0.803 | −0.223 |
| SOD | 0.053 | −0.706 | −0.298 |
| Sucrose | −0.770 | −0.169 | 0.178 |
| Fructose | 0.685 | 0.220 | 0.452 |
| Eigen value | 7.991 | 5.023 | 1.731 |
| Cumulative% | 47.01 | 76.56 | 86.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Ma, J.; Sun, X.; Guo, X. Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau. Agriculture 2022, 12, 1362. https://doi.org/10.3390/agriculture12091362
Zheng L, Ma J, Sun X, Guo X. Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau. Agriculture. 2022; 12(9):1362. https://doi.org/10.3390/agriculture12091362
Chicago/Turabian StyleZheng, Lijian, Juanjuan Ma, Xihuan Sun, and Xianghong Guo. 2022. "Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau" Agriculture 12, no. 9: 1362. https://doi.org/10.3390/agriculture12091362
APA StyleZheng, L., Ma, J., Sun, X., & Guo, X. (2022). Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau. Agriculture, 12(9), 1362. https://doi.org/10.3390/agriculture12091362