
Nitrogen Nutrition Differentially Affects Concentrations of Photosynthetic Pigments and Antioxidant Compounds in Mexican Marigold (Tagetes erecta L.)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Plant Material
2.2. Experiment Management
2.3. Evaluated Treatments
2.4. Experimental Design
2.5. Study Variables
2.5.1. Chlorophylls a, b, Total Chlorophylls, and Chlorophyll a/Chlorophyll b Ratio
2.5.2. Total Carotenoids
2.5.3. Total Phenolic Compounds
2.5.4. Total Flavonoids
2.5.5. Antioxidant Activity
2.6. Statistical Analysis of the Results
3. Results
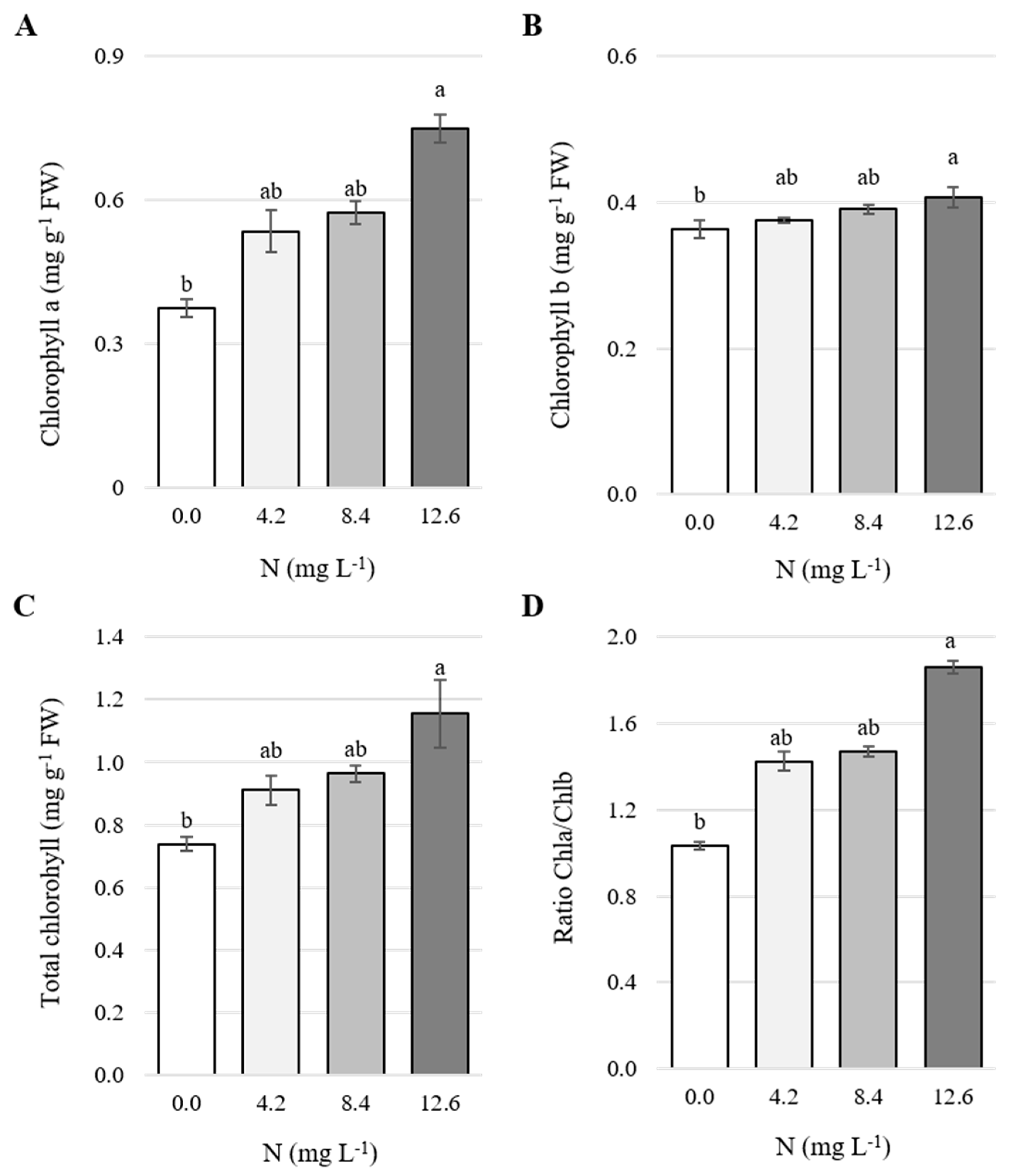
3.1. Concentration of Chlorophylls in Leaves
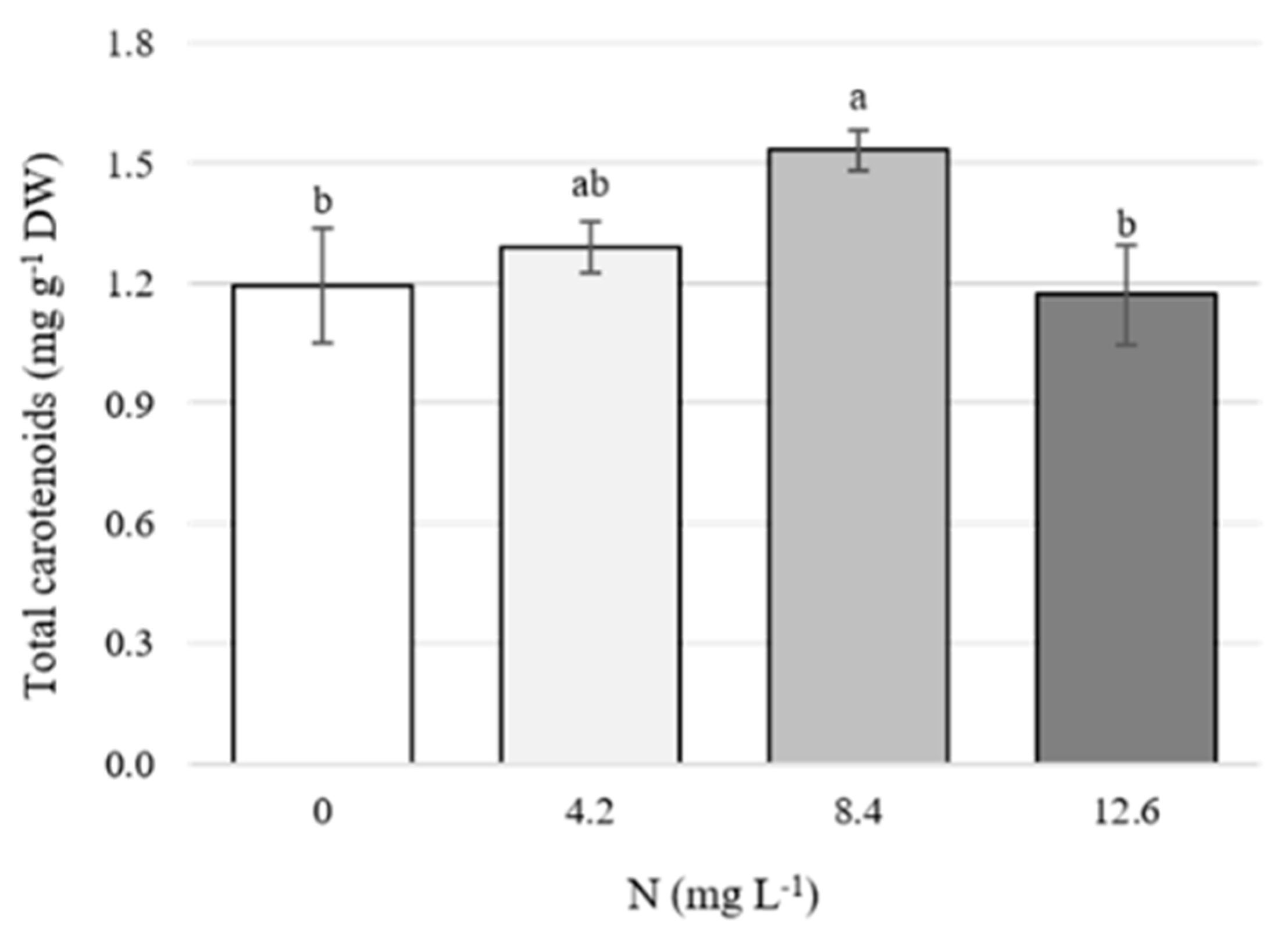
3.2. Concentrations of Total Carotenoids in Flowers
3.3. Concentration of Total Phenols in Leaves and Flowers
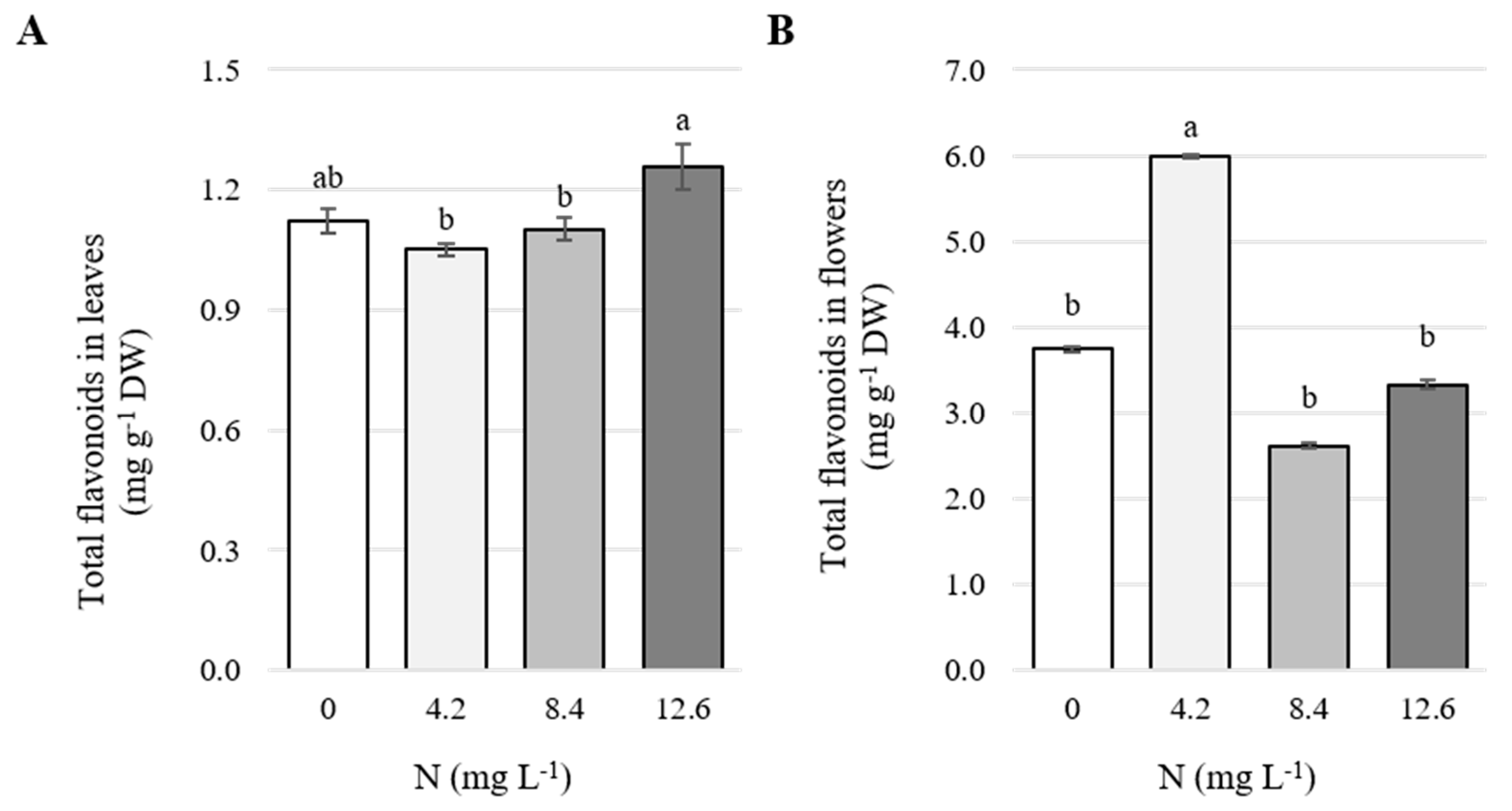
3.4. Concentration of Total Flavonoids in Leaves and Flowers
3.5. Antioxidant Activity in Flowers
4. Discussion
4.1. Concentration of Chlorophylls
4.2. Concentration of Total Carotenoids in Flowers
4.3. Concentration of Total Phenolic Compounds in Leaves and Flowers
4.4. Concentrations of Total Flavonoids in Leaves and Flowers
4.5. Antioxidant Activity in Flowers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Casassola, A.; Patussi, B.S. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. In Abiotic and Biotic Stress in Plants; Shanker, A.K., Shanker, C., Eds.; Intech Open: Rijeka, Croatia, 2016; pp. 463–480. [Google Scholar] [CrossRef] [Green Version]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minatel, I.O.; Vanz, B.C.; Ferreria, M.I.; Gomez, G.H.A.; Chen-Yen, O.C.; Pereira, L.G.P. Phenolic compounds: Functional properties, impact of processing and bioavailability. In Phenolic Compounds. Biological Activity; Soto-Hernández, M., Palma-Tenango, M., García-Mateos, M.R., Eds.; Intech Open: Rijeka, Croatia, 2017; pp. 1–24. [Google Scholar]
- Bessada, S.M.F.; Barreira, J.C.M.; Oliveira, M.B.P.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crops Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Burlec, A.F.; Arsene, C.; Gille, E.; Hăncianu, M.; Cioancă, O. Ornamental Asteraceae species as new sources of secondary metabolites. Indian J. Pharm. Educ. Res. 2017, 51, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Mir, R.A.; Ahanger, M.A.; Agarwal, R.M. Marigold: From mandap to medicine and from ornamentation to remediation. Am. J. Plant Sci. 2019, 10, 309–338. [Google Scholar] [CrossRef] [Green Version]
- Siddhu, N.; Saxena, J. Evaluation of in vitro antioxidant activity of flowers of Tagetes erecta. Int. J. Pharmacogn. Phytochem. Res. 2017, 9, 975–979. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Rahman, Z.A. Involvement of nitrogen of flavonoids, glutathione, anthocyanin, ascorbic acid and antioxidant activities of Malaysian medicinal plant Labisia pumila Blume (Kacip Fatimah). Int. J. Mol. Sci. 2012, 13, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Rahman, Z.A. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of Kacip Fatimah (Labisia pumila Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Carotenoid pigments in kale are influenced by nitrogen concentration and form. J. Sci. Food Agric. 2007, 87, 900–907. [Google Scholar] [CrossRef]
- Chenard, C.H.; Kopsell, D.A.; Kopsell, D.E. Nitrogen concentration affects nutrient and carotenoid accumulation in parsley. J. Plant Nutr. 2005, 28, 285–297. [Google Scholar] [CrossRef]
- Deng, B.; Li, Y.; Xu, D.; Ye, Q.; Liu, G. Nitrogen availability alters flavonoid accumulation in Cyclocarya paliurus via the effects on the internal carbon/nitrogen balance. Sci. Rep. 2019, 9, 2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biesiada, A.; Sokół-Łętowska, A.; Kucharska, A. The effect of nitrogen fertilization on yielding and antioxidant activity of lavender (Lavandula angustifolia Mill.). Acta Sci. Pol. Hortorum Cultus 2008, 7, 33–40. [Google Scholar]
- Li, J.; Zhu, Z.; Gerendás, J. Effects of nitrogen and sulfur on total phenolics and antioxidant activity in two genotypes of leaf mustard. J. Plant Nutr. 2008, 31, 1642–1655. [Google Scholar] [CrossRef]
- Król, B. The effect of different nitrogen fertilization rates on yield and quality of marigold (Calendula officinalis L. Tokaj) raw material. Acta Agrobot. 2011, 64, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Shafiullah, M.I.; Shah, F.A.; Khan, T.; Khan, T.; Ahmad, I.; Shahid, M.A.; Khan, S. Response of Marigold (Tagetes erecta L.) to different levels of nitrogen at Bagh E Naran Park Peshawar. Int. J. Environ. Sci. Nat. Res. 2018, 14, 555873. [Google Scholar] [CrossRef]
- Souri, M.K.; Rahmani, R.; Ahmadi, M.; Rezainejad, A. Morphophysiological and nutrient uptake characteristics of marigold (Tagetes spp.) under different levels of nitrogen fertilization. Horticult. Int. J. 2018, 2, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A. The universal nutrient solution. In Proceedings of the 6th International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984; pp. 633–649. [Google Scholar]
- Trejo-Téllez, L.I.; Peralta-Sánchez, M.G.; Gómez-Merino, F.C.; Rodríguez-Mendoza, M.N.; Serrato-Cruz, M.A.; Arévalo-Becerril, A.E. Sodium chloride on dry biomass and macronutrient cations absorption in cempasúchil (Tagetes erecta Linn.). Rev. Mex. Cienc. Agríc. 2013, 4, 979–990. [Google Scholar]
- Harbone, J.B. Clorophyll extraction. In Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Harbone, J.B., Ed.; Chapman and Hall: London, UK, 1973; pp. 205–207. [Google Scholar]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaish 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Waterman, P.G.; Mole, S. Analysis of Phenolic Plant Metabolites, 1st ed.; Blackwell Scientific Publications: Oxford, UK, 1994; 238p. [Google Scholar]
- Gong, Y.; Liu, X.; He, W.-H.; Xu, H.-G.; Yuan, F.; Gao, Y.-X. Investigation into the antioxidant activity and chemical composition of alcoholic extracts from defatted marigold (Tagetes erecta L.) residue. Fitoterapia 2012, 83, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, K.; Ghasemi, Y.; Ebrahimzadeh, M.A. Antioxidant activity, phenol and flavonoid contents of 13 citrus species peels and tissues. Pak. J. Pharm. Sci. 2009, 22, 227–281. [Google Scholar]
- Ebrahimzadeh, M.A.; Pourmorad, F.; Bekhradnia, A.R. Iron chelating activity, phenol and flavonoid content of some medicinal plants from Iran. Afr. J. Biotechnol. 2008, 7, 3188–3192. [Google Scholar]
- Nanyonga, S.K.; Opoku, A.R.; Lewu, F.B.; Oyedeji, O.O.; Singh, M.; Oyedeji, A.O. Antioxidant activity and cytotoxicity of the leaf and bark extracts of Tarchonanthus camphorates. Trop. J. Pharm. Res. 2013, 12, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, C.B.; Ramos, L.E.; Ibáñez, V.L. Evaluación de la capacidad antioxidante de siete plantas medicinales peruanas. Rev. Horiz. Méd. 2008, 8, 56–72. [Google Scholar]
- Chizzola, R.; Michistschs, H.; Franz, C. Antioxidative properties of Thymus vulgaris leaves: Comparison of different extracts and essential oil chemotypes. J. Agri. Food Chem. 2008, 56, 6897–6904. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, E.E.; Pacheco, S.M.; García, M.R.; San Miguel, C.R.; Ramírez, V.G.; Soto, H.R.M. Actividad antioxidante de alcaloides de Erythrina americana Miller. Rev. Fitotec. Mex. 2011, 34, 241–246. [Google Scholar]
- Kuskoski, E.M.; Asuero, A.G.; Troncoso, A.M.; Mancini-Filho, J.; Fett, R. Aplicación de diversos métodos químicos para determinar actividad antioxidante en pulpa de frutos. Ciênc. Tecnol. Aliment. 2005, 25, 726–732. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute Inc. SAS/STAT Users Guide, Version 9.3; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Hassanain, M.A.; Osman, A.R.; Sewedan, E.; El-Dein, M.S. Influence of nitrogen and yeast on production of marigold plants Tagetes erecta. World Rural Observ. 2018, 10, 37–46. [Google Scholar] [CrossRef]
- Polara, N.D.; Gajipara, N.N.; Barad, A.V. Effect of nitrogen and phosphorus nutrition on growth, flowering, flower yield and chlorophyll content of different varieties of African marigold (Tagetes erecta L.). J. Appl. Hortic. 2015, 17, 44–47. [Google Scholar] [CrossRef]
- Porto, S.R.M.; Deitos, F.D.; Vieira, P.A.J.; Bonomo, P.; Silva, S.I.; Nascimento, C.C.; Rodriguez, B.P.H.; Santos, B.M. Chlorophyll and carbohydrates in Arachis pintoi plants under influence of water regimes and nitrogen fertilization. Rev. Bras. Zootec. 2013, 42, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Abubakar, A.W.; Manga, A.A.; Kamara, A.Y.; Tofa, A.I. Physiological evaluations of maize hybrids under low nitrogen. Adv. Agric. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bojović, B.; Marković, A. Correlation between nitrogen and chlorophyll content in wheat (Triticum aestivum L.). Kragujev. J. Sci. 2009, 31, 69–74. [Google Scholar]
- Gárate, A.; Bonilla, I. Nutrición mineral y producción vegetal. In Fundamento de Fisiología Vegetal; Azcón-Bieto, J., Talón, M., Eds.; McGraw-Hill Interamericana: Barcelona, España, 2009; pp. 113–129. [Google Scholar]
- Fornari, Z.E.; Gaviraghi, L.; Basso, C.J.; Pinheiro, M.M.V.; Vian, A.L.; Santi, A.L. Relationship between photosynthetic pigments and corn production under nitrogen sources. Pesqui. Agropecu. Trop. 2020, 50, e63661. [Google Scholar] [CrossRef]
- Wang, S.; Guan, K.; Wang, Z.; Ainsworth, E.A.; Zheng, T.; Townsend, P.A.; Li, K.; Moller, C.; Wu, G.; Jiang, C. Unique contributions of chlorophyll and nitrogen to predict crop photosynthetic capacity from leaf spectroscopy. J. Exp. Bot. 2021, 72, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2016, 23, 3513–3524. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Porto, J.S.; Reboucas, T.N.H.; Moraes, M.O.B.; Bomfim, M.P.; Lemos, O.L.; Luz, J.M.Q. Quality and antioxidant activity of tomato cultivated under different sources and doses of nitrogen. Rev. Caatinga Mossoró 2016, 29, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Duan, Y.; Tan, F.; Zhang, S. Effects of long-term nitrogen and organic fertilization on antioxidants content of tomato fruits. J. Hortic. 2016, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P. Carotenoids of therapeutic significance from marigold. Nat. Prod. Chem. Res. 2014, 2, e110. [Google Scholar] [CrossRef] [Green Version]
- Legha, M.R.; Prasad, K.V.; Singh, S.K.; Kaur, C.; Arora, A.; Kumar, S. Induction of carotenoid pigments in callus cultures of Calendula officinalis L. in response to nitrogen and sucrose levels. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 99–106. [Google Scholar] [CrossRef]
- Razaq, M.; Zhang, P.; Hai-long, S.; Salahuddin. Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinsi, B.; Negrini, N.; Morgutti, S.; Espen, L. Nitrogen starvation and nitrate or ammonium availability differently affect phenolic composition in green and purple basil. Agronomy 2020, 10, 498. [Google Scholar] [CrossRef] [Green Version]
- Elhanafi, L.; Houhou, M.; Rais, C.; Mansouri, I.; Elghadraoui, L.; Greche, H. Impact of excessive nitrogen fertilization on the biochemical quality, phenolic compounds, and antioxidant power of Sesamum indicum L. seeds. J. Food Qual. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Allahdadi, M.; Farzane, P. Influence of different levels of nitrogen fertilizer on some phytochemical characteristics of artichoke (Cynara scolymus L.) leaves. J. Med. Plants Stud. 2018, 6, 109–115. [Google Scholar]
- Onyango, C.M.; Harbinson, J.; Imungi, J.K.; Onwonga, R.N.; van Kooten, O. Effect of nitrogen source, crop maturity stage and storage conditions on phenolics and oxalate contents in vegetable amaranth (Amaranthus hypochondriacus). J. Agric. Sci. 2012, 4, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.; Urlić, B.; Špika, M.J.; Kläring, H.P.; Krumbein, A.; Baldermann, S.; Ban, S.G.; Perica, S.; Schwarz, D. Nitrogen limited red and green leaf lettuce accumulate flavonoid glycosides, caffeic acid derivatives, and sucrose while losing chlorophylls, β-carotene and xanthophylls. PLoS ONE 2015, 10, e0142867. [Google Scholar] [CrossRef]
- Stumpf, B.; Yan, F.; Honermeier, B. Influence of nitrogen fertilization on yield phenolic compounds in wheat grains (Triticum aestivum L. ssp. aestivum). J. Plant Nutr. Soil Sci. 2019, 182, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Yañez-Mansilla, E.; Cartes, P.; Reyes-Díaz, M.; Ribera-Fonseca, A.; Rengel, Z.; Lobos, W.; Alberdi, M. Leaf nitrogen thresholds ensuring high antioxidant features of Vaccinium corymbosum cultivars. J. Soil Sci. Plant Nutr. 2015, 15, 574–586. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Amarowicz, R.; Cwalina-Ambroziak, B.; Janiak, M.A.; Bogucka, B. Effect of N fertilization on the content of phenolic compounds in Jerusalem Artichoke (Helianthus tuberosus L.) tubers and their antioxidant capacity. Agronomy 2020, 10, 1215. [Google Scholar] [CrossRef]
- Peled-Zehavi, H.; Oliva, M.; Xie, Q.; Tzin, V.; Oren-Shamir, M.; Aharoni, A.; Galili, G. Metabolic engineering of the phenylpropanoid and its primary, precursor pathway to enhance the flavor of fruit and the aroma of flowers. Bioengineering 2015, 2, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Munene, R.; Changamu, E.; Koir, N.; Gweyi-Onyango, J. Effects of different nitrogen forms on growth, phenolics, flavonoids and antioxidant activity in amaranth species. Trop. Plant Res. 2017, 4, 81–89. [Google Scholar] [CrossRef]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Xie, Y.; Hu, L.; Si, J.; Wang, Z. Excessive nitrogen application dampens antioxidant capacity and grain filling in wheat as revealed by metabolic and physiological analyses. Sci. Rep. 2017, 7, 43363. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent Grade Salts | Nitrogen in the Nutrient Solution (g L−1) | |||
|---|---|---|---|---|
| 0 | 4.2 | 8.4 | 12.6 | |
| (mg L−1) | ||||
| KH2PO4 | 6.8045 | 6.8045 | 6.8045 | 6.8045 |
| Ca(OH)2 | 16.6709 | – | – | – |
| Ca(NO3)2 4H2O | – | 35.4225 | 53.1338 | 53.1338 |
| MgSO4 7H2O | 24.6470 | 24.6470 | 24.6470 | 24.6470 |
| K2SO4 | 13.0688 | 13.0688 | 13.0688 | 13.0688 |
| KOH | 8.4158 | 8.4158 | – | – |
| KNO3 | – | – | 15.1650 | 15.1650 |
| CH4N2O | – | – | – | 7.7220 |
| Nitrogen in the Nutrient Solution (mg L−1) | Antioxidant Activity (μM eq. Trolox g−1 Fresh Matter) |
|---|---|
| 0 | 1125.62 ± 79.4 b |
| 4.2 | 1722.60 ± 2.8 a |
| 8.4 | 1690.21 ± 4.2 a |
| 12.6 | 1495.45 ± 180.8 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peralta-Sánchez, M.G.; Gómez-Merino, F.C.; Tejeda-Sartorius, O.; Trejo-Téllez, L.I. Nitrogen Nutrition Differentially Affects Concentrations of Photosynthetic Pigments and Antioxidant Compounds in Mexican Marigold (Tagetes erecta L.). Agriculture 2023, 13, 517. https://doi.org/10.3390/agriculture13030517
Peralta-Sánchez MG, Gómez-Merino FC, Tejeda-Sartorius O, Trejo-Téllez LI. Nitrogen Nutrition Differentially Affects Concentrations of Photosynthetic Pigments and Antioxidant Compounds in Mexican Marigold (Tagetes erecta L.). Agriculture. 2023; 13(3):517. https://doi.org/10.3390/agriculture13030517
Chicago/Turabian StylePeralta-Sánchez, María Guadalupe, Fernando C. Gómez-Merino, Olga Tejeda-Sartorius, and Libia I. Trejo-Téllez. 2023. "Nitrogen Nutrition Differentially Affects Concentrations of Photosynthetic Pigments and Antioxidant Compounds in Mexican Marigold (Tagetes erecta L.)" Agriculture 13, no. 3: 517. https://doi.org/10.3390/agriculture13030517
APA StylePeralta-Sánchez, M. G., Gómez-Merino, F. C., Tejeda-Sartorius, O., & Trejo-Téllez, L. I. (2023). Nitrogen Nutrition Differentially Affects Concentrations of Photosynthetic Pigments and Antioxidant Compounds in Mexican Marigold (Tagetes erecta L.). Agriculture, 13(3), 517. https://doi.org/10.3390/agriculture13030517