
A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia)

 ,
, 

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
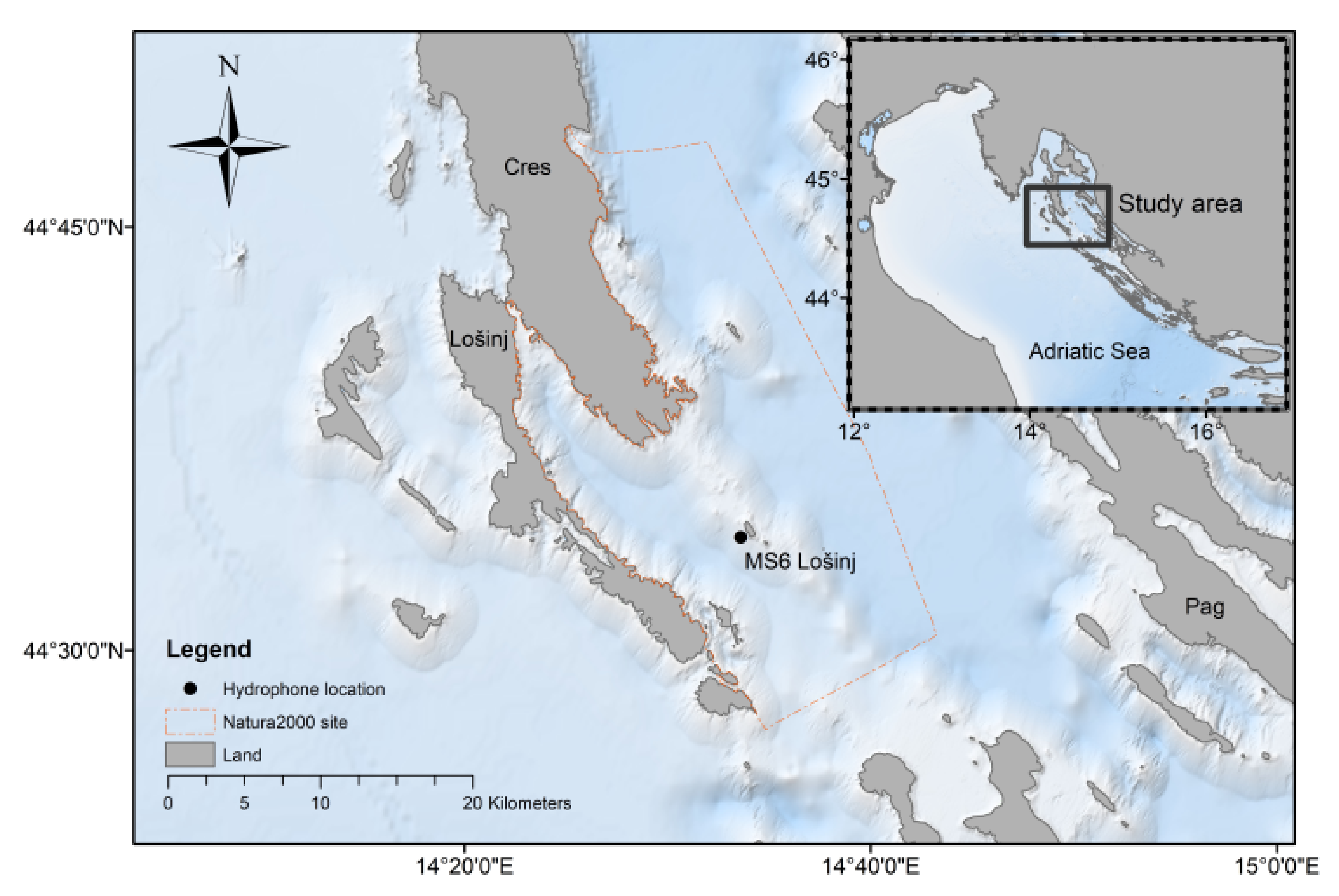
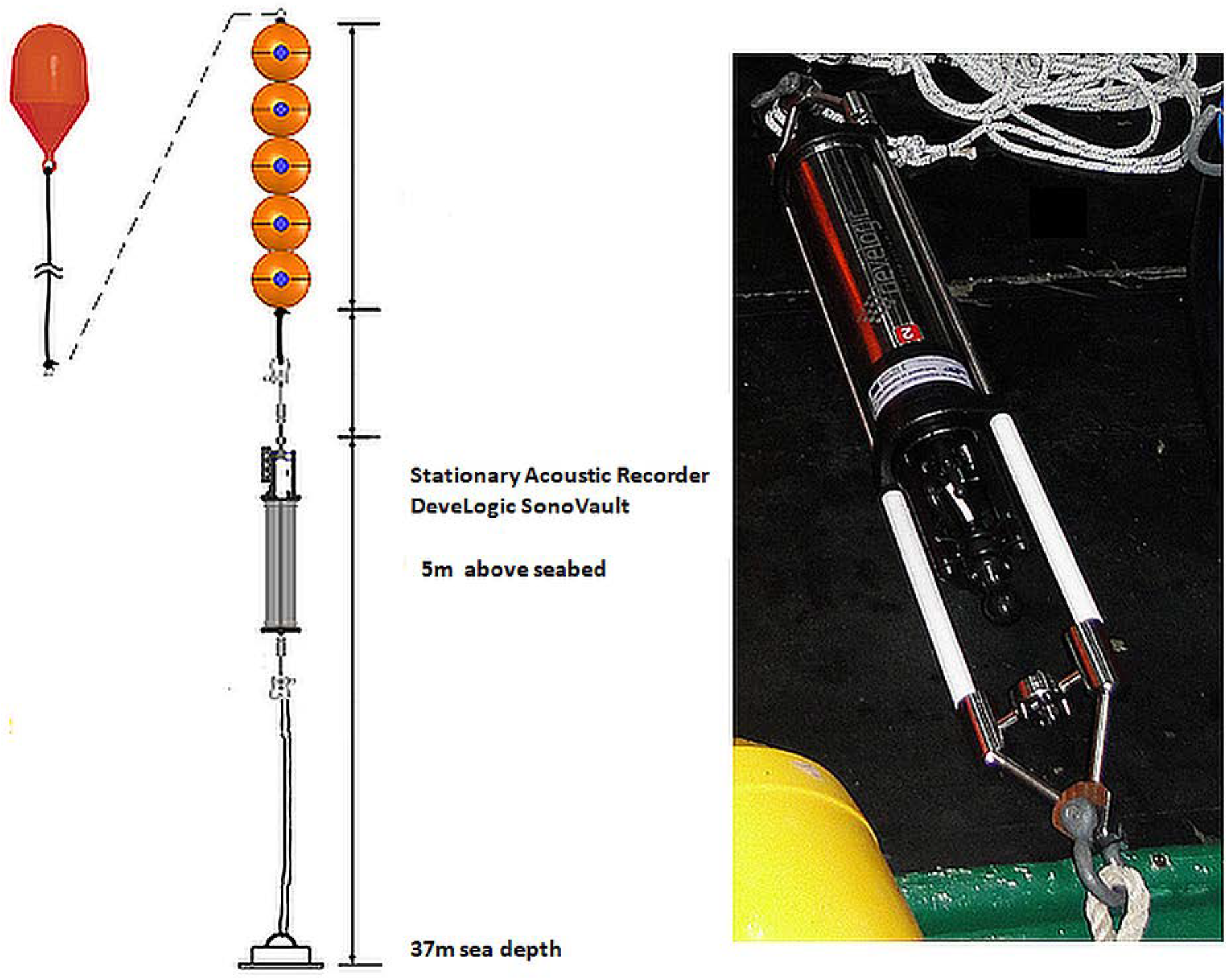
2.1. The Study Site and Data Collection
2.2. Data Analysis
2.2.1. Manual Scrolling
2.2.2. Fish Sound Analysis
2.2.3. Sound Pressure Levels Analysis
- p is the pressure recorded by hydrophone,
- p0 is the reference pressure.
2.2.4. Statistical Analysis
3. Results
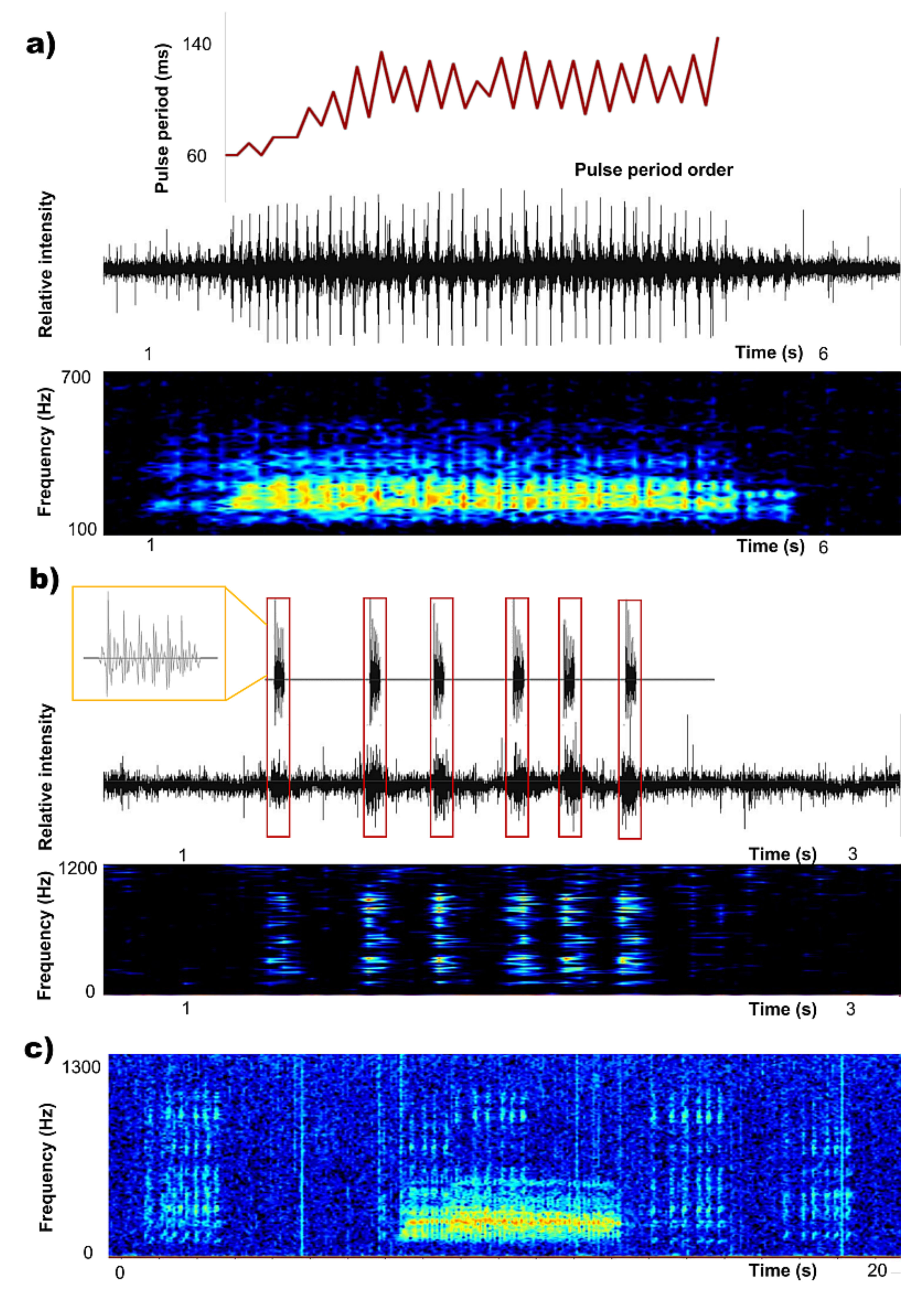
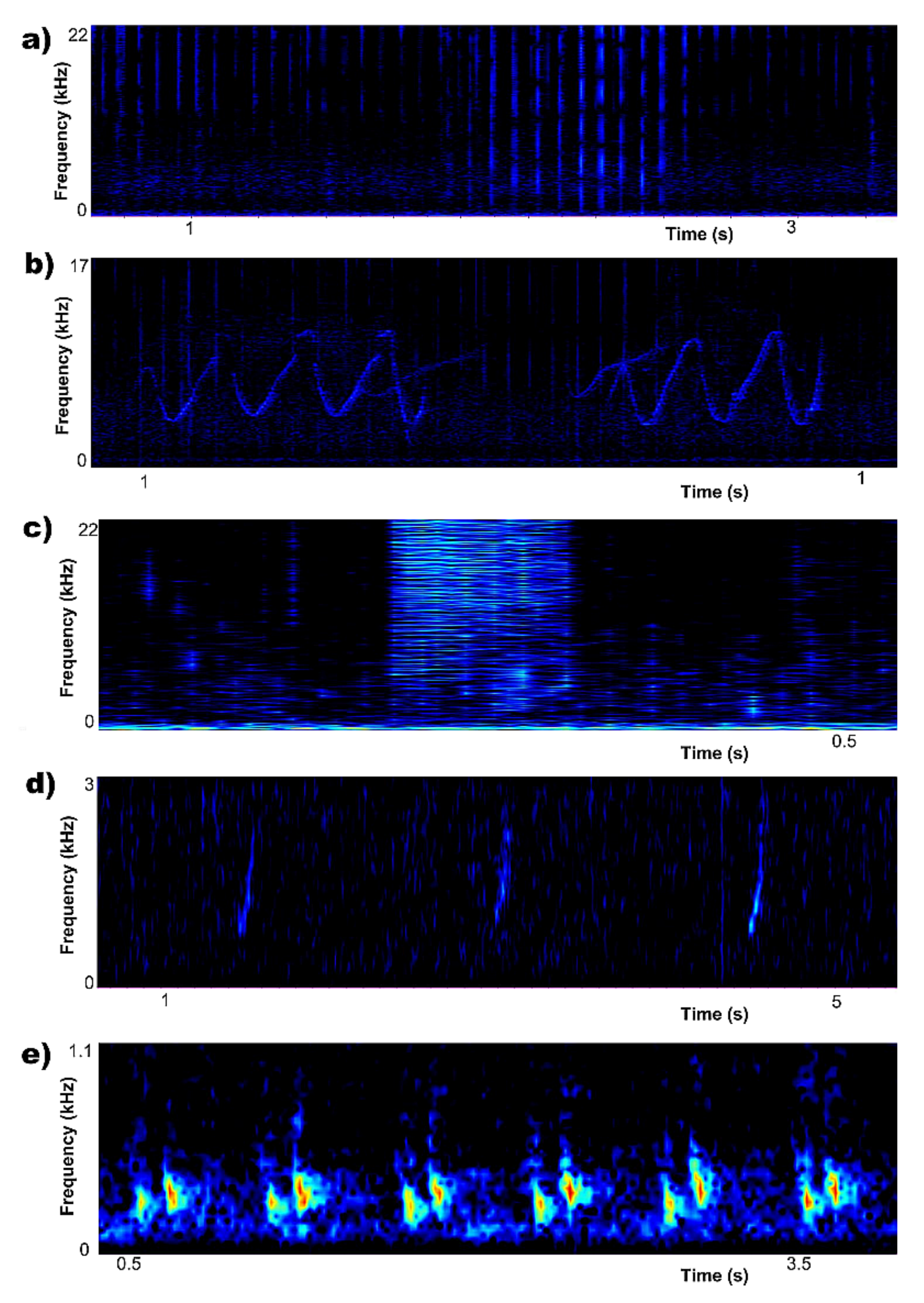
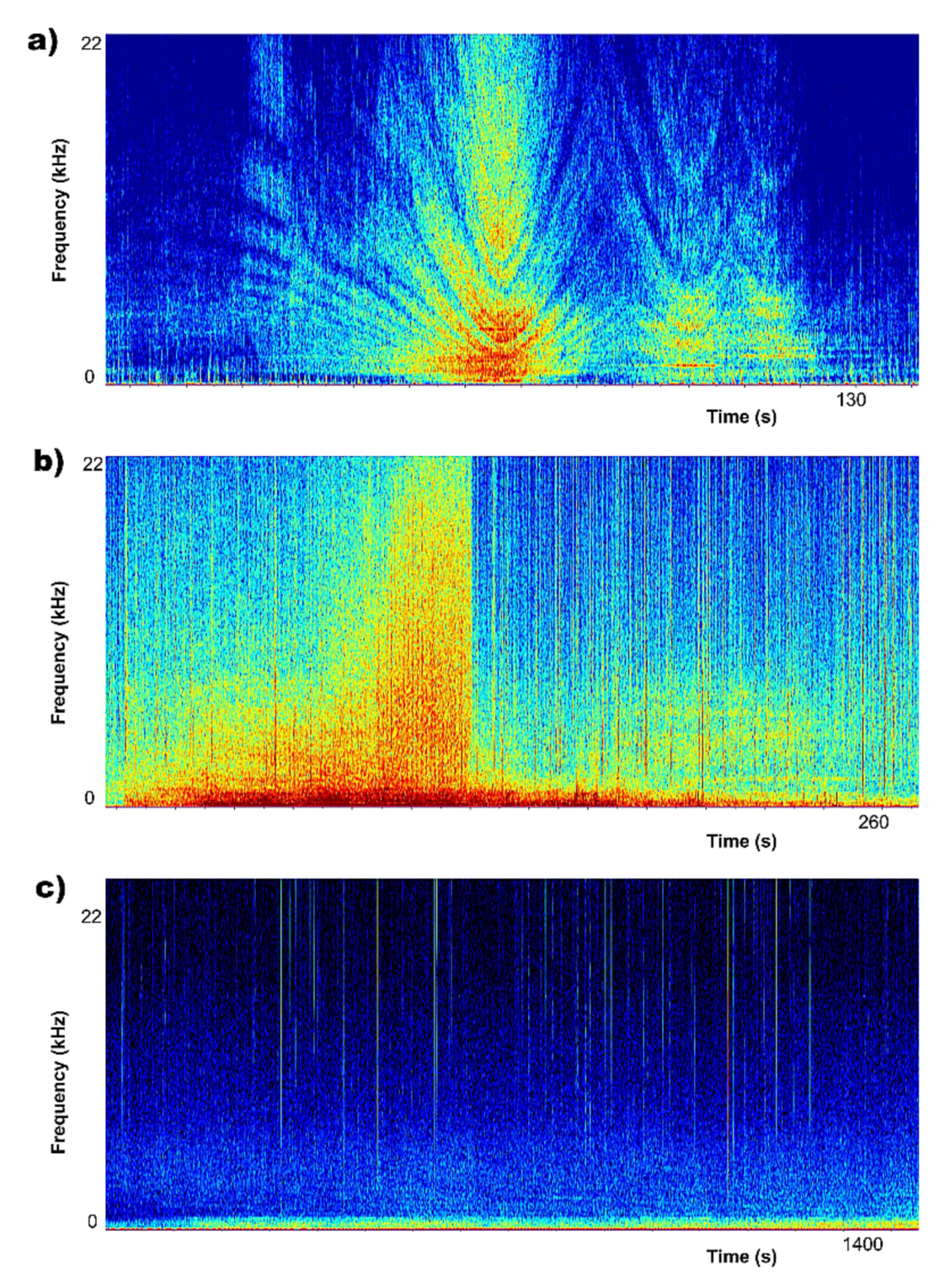
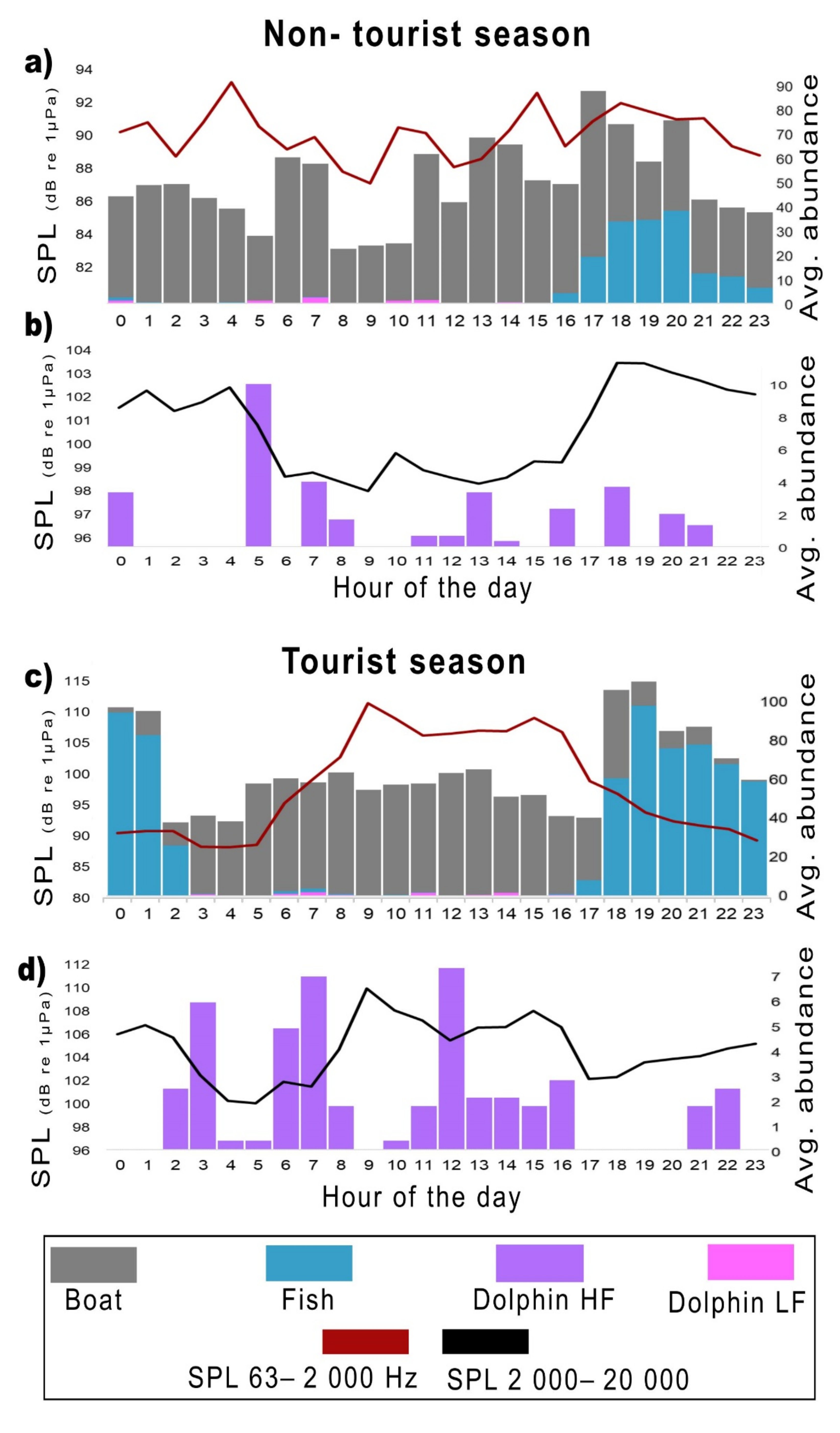
3.1. Biophony and Anthrophony Characterizing the Local Soundscape
- (a)
- (b)
- (c)
- (d)

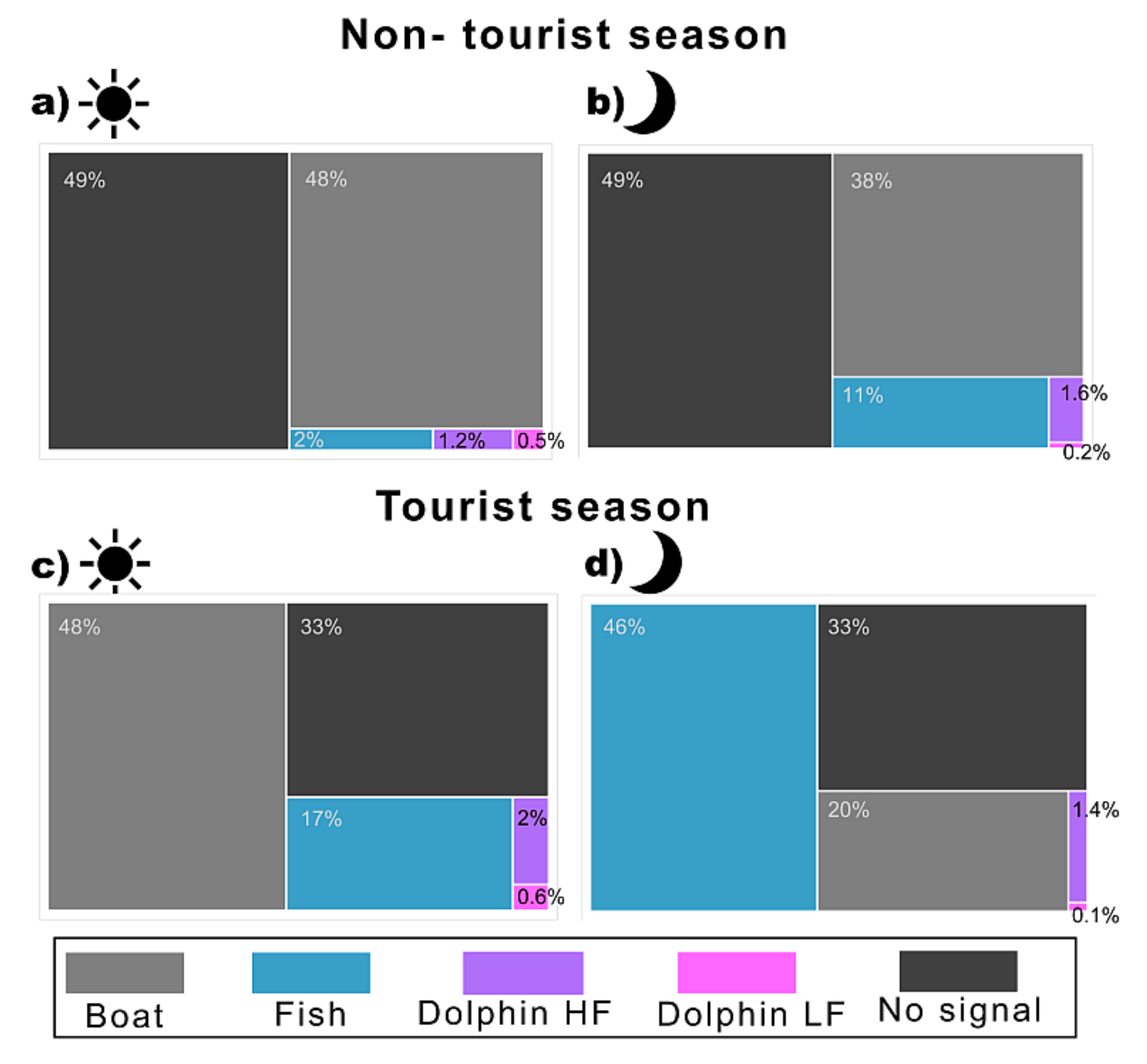
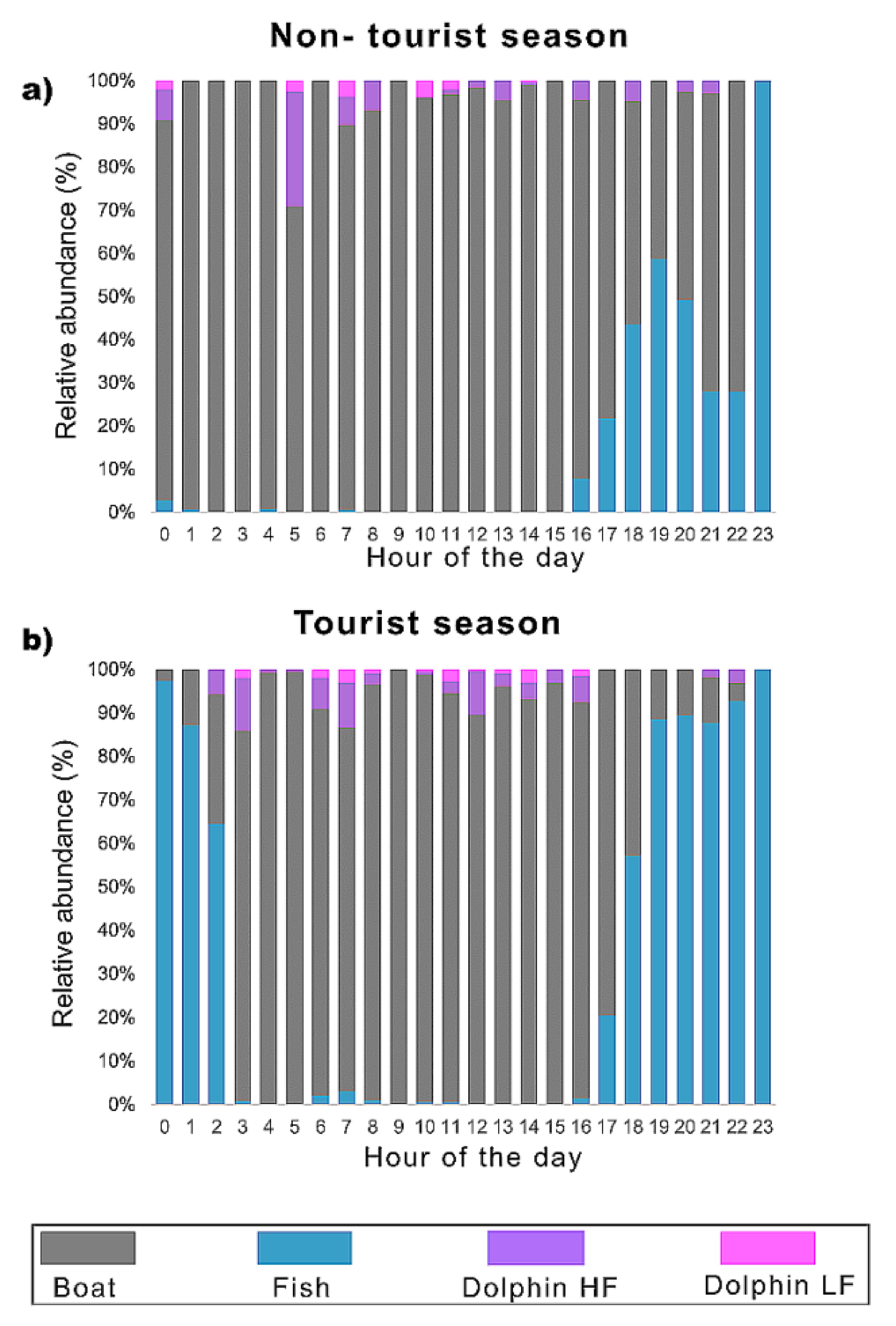
3.2. Temporal Distribution of the Biophony and Anthrophony of the Local Soundscape
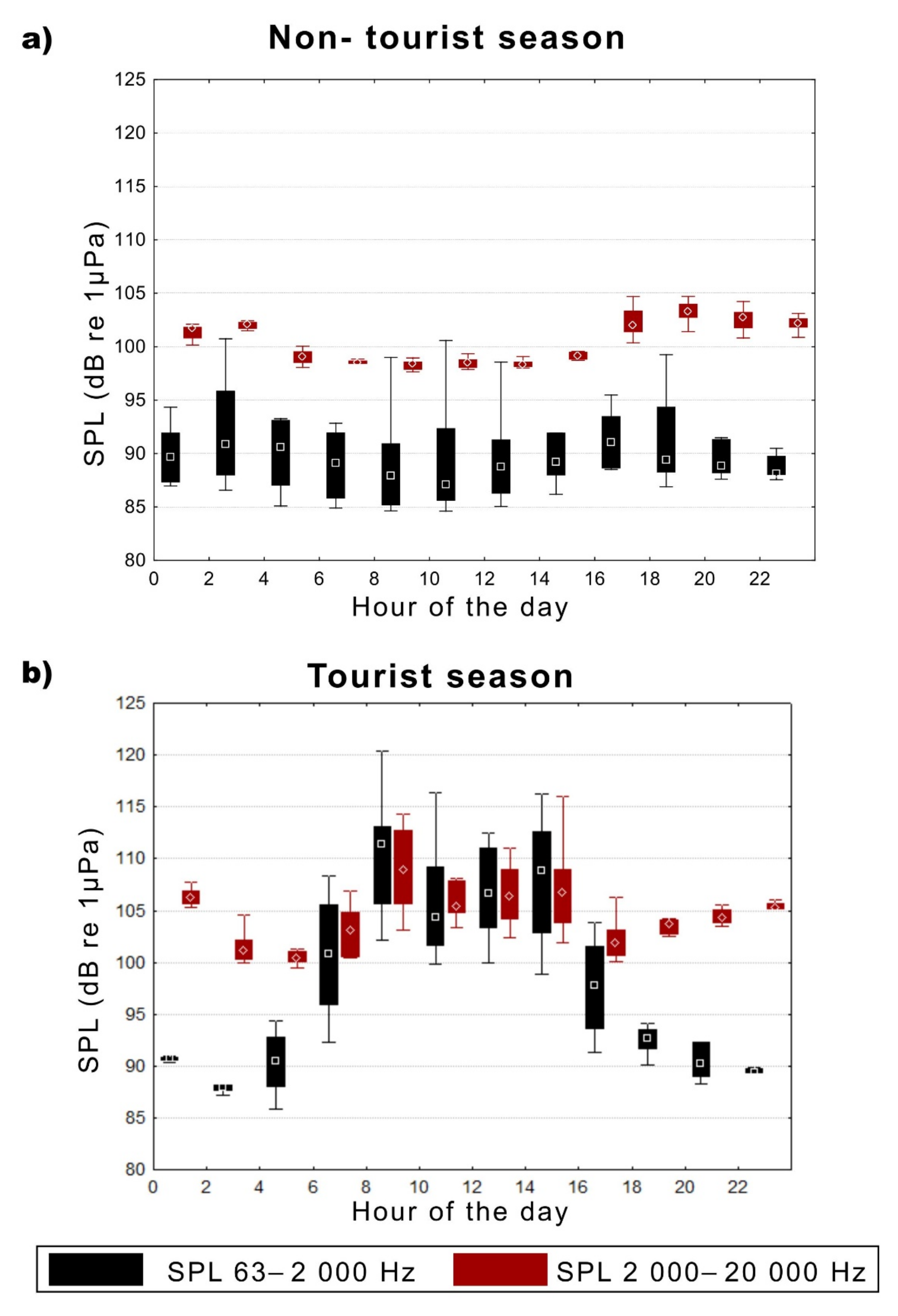
3.3. Sound Pressure Levels (SPLs) in the Study Area
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pijanowski, B.C.; Villanueva-Rivera, L.J.; Dumyahn, S.L.; Farina, A.; Krause, B.L.; Napoletano, B.M.; Gage, S.H.; Pieretti, N. Soundscape ecology: The science of sound in the landscape. BioScience 2011, 61, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Lillis, A.; Eggleston, D.B.; Bohnenstiehl, D.R. Estuarine soundscapes: Distinct acoustic characteristics of oyster reefs compared to soft-bottom habitats. Mar. Ecol. Prog. Ser. 2014, 505, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Farina, A.; Gage, S.H. Ecoacoustics: The Ecological Role of Sounds; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 95–108. [Google Scholar]
- Farina, A. Ecoacoustics: A quantitative approach to investigate the ecological role of environmental sounds. Mathematics 2019, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Desiderà, E.; Guidetti, P.; Panzalis, P.; Navone, A.; Valentini-Poirrier, C.A.; Boissery, P.; Gervaise, C.; Di Iorio, L. Acoustic fish communities: Sound diversity of rocky habitats reflects fish species diversity. Mar. Ecol. Prog. Ser. 2019, 608, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Di Iorio, L.; Audax, M.; Deter, J.; Holon, F.; Lossent, J.; Gervaise, C.; Boissery, P. Biogeography of acoustic biodiversity of NW Mediterranean coralligenous reefs. Sci. Rep. 2021, 11, 16991. [Google Scholar] [CrossRef]
- McWilliam, J.N.; Hawkins, A.D. A comparison of inshore marine soundscapes. J. Exp. Mar. Biol. Ecol. 2013, 446, 166–176. [Google Scholar] [CrossRef]
- Buscaino, G.; Ceraulo, M.; Pieretti, N.; Corrias, V.; Farina, A.; Filiciotto, F.; Maccarrone, V.; Grammauta, R.; Caruso, F.; Giuseppe, A.; et al. Temporal patterns in the soundscape of the shallow waters of a Mediterranean marine protected area. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pieretti, N.; Martire, M.L.; Farina, A.; Danovaro, R. Marine soundscape as an additional biodiversity monitoring tool: A case study from the Adriatic Sea (Mediterranean Sea). Ecol. Indic. 2017, 83, 13–20. [Google Scholar] [CrossRef]
- Ceraulo, M.; Papale, E.; Caruso, F.; Filiciotto, F.; Grammauta, R.; Parisi, I.; Mazzola, S.; Farina, A.; Buscaino, G. Acoustic comparison of a patchy Mediterranean shallow water seascape: Posidonia oceanica meadow and sandy bottom habitats. Ecol. Indic. 2018, 85, 1030–1043. [Google Scholar] [CrossRef]
- Picciulin, M.; Kéver, L.; Parmentier, E.; Bolgan, M. Listening to the unseen: Passive acoustic monitoring reveals the presence of a cryptic fish species. Aquat. Conserv. 2019, 29, 202–210. [Google Scholar] [CrossRef]
- Vieira, M.; Fonseca, P.J.; Amorim, M.C.P. Fish sounds and boat noise are prominent soundscape contributors in an urban European estuary. Mar. Pollut. Bull. 2021, 172, 112845. [Google Scholar] [CrossRef] [PubMed]
- Warren, V.E.; McPherson, C.; Giorli, G.; Goetz, K.T.; Radford, C.A. Marine soundscape variation reveals insights into baleen whales and their environment: A case study in central New Zealand. R. Soc. Open Sci. 2021, 8, 201503. [Google Scholar] [CrossRef] [PubMed]
- Pieretti, N.; Martire, M.L.; Corinaldesi, C.; Musco, L.; Dell’Anno, A.; Danovaro, R. Anthropogenic noise and biological sounds in a heavily industrialized coastal area (Gulf of Naples, Mediterranean Sea). Mar. Environ. Res. 2020, 159, 105002. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.N.; Kerridge, E.; Simpson, S.D. Acoustic communication in a noisy world: Can fish compete with anthropogenic noise? Behav. Ecol. 2014, 25, 1022–1030. [Google Scholar] [CrossRef] [Green Version]
- Kunc, H.P.; McLaughlin, K.E.; Schmidt, R. Aquatic noise pollution: Implications for individuals, populations, and ecosystems. R. Soc. Lond. B. Biol. Sci. 2016, 283, 20160839. [Google Scholar] [CrossRef] [Green Version]
- Cox, K.; Brennan, L.P.; Gerwing, T.G.; Dudas, S.E.; Juanes, F. Sound the alarm: A meta-analysis on the effect of aquatic noise on fish behavior and physiology. Glob. Chang. Biol. 2018, 24, 3105–3116. [Google Scholar] [CrossRef]
- Seitz, R.D.; Wennhage, H.; Bergström, U.; Lipcius, R.N.; Ysebaert, T. Ecological value of coastal habitats for commercially and ecologically important species. ICES J. Mar. Sci. 2014, 71, 648–665. [Google Scholar] [CrossRef] [Green Version]
- Airoldi, L.; Beck, M.W. Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. Annu. Rev. 2007, 45, 345–405. [Google Scholar]
- Claudet, J.; Fraschetti, S. Human-driven impacts on marine habitats: A regional meta-analysis in the Mediterranean Sea. Biol. Conserv. 2010, 143, 2195–2206. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: Assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [Green Version]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben RaisLasram, F.; Cheung, W.W.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Bearzi, G.; di Sciara, G.; Politi, E. Social ecology of bottlenose dolphins in the Kvarnerić (northern Adriatic Sea). Mar. Mamm. Sci. 1997, 13, 650–668. [Google Scholar] [CrossRef]
- Bearzi, G.; Politi, E.; di Sciara, G.N. Diurnal behavior of free-ranging bottlenose dolphins in the Kvarnerić (Northern Adriatic Sea). Mar. Mamm. Sci. 1999, 15, 1065–1097. [Google Scholar] [CrossRef]
- Pleslić, G.; RakoGospić, N.; Mackelworth, P.; Wiemann, A.; Holcer, D.; Fortuna, C. The abundance of common bottlenose dolphins (Tursiops truncatus) in the former special marine reserve of the Cres-Lošinj Archipelago, Croatia. Aquat. Conserv. 2015, 25, 125–137. [Google Scholar] [CrossRef]
- Bearzi, G.; Fortuna, C.; Reeves, R. Tursiops truncatus (Mediterranean subpopulation). In The IUCN Red List of Threatened Species; IUCN Global Species Programme Red List Unit: Cambridge, UK, 2012; p. e.T16369383A16369386. [Google Scholar]
- Mackelworth, P.; Holcer, D.; Fortuna, C.M. Unbalanced governance: The Cres-Lošinj special marine reserve, a missed conservation opportunity. Mar. Policy 2013, 41, 126–133. [Google Scholar] [CrossRef]
- Rako, N.; Fortuna, C.M.; Holcer, D.; Mackelworth, P.; Nimak-Wood, M.; Pleslić, G.; Sebastianutto, L.; Vilibić, I.; Wiemann, A.; Picciulin, M. Leisure boating noise as a trigger for the displacement of the bottlenose dolphins of the Cres–Lošinj archipelago (northern Adriatic Sea, Croatia). Mar. Pollut. Bull. 2013, 68, 77–84. [Google Scholar] [CrossRef]
- RakoGospić, N.R.; Picciulin, M. Changes in whistle structure of resident bottlenose dolphins in relation to underwater noise and boat traffic. Mar. Pollut. Bull. 2016, 105, 193–198. [Google Scholar] [CrossRef]
- Jézéquel, Y.; Chauvaud, L.; Bonnel, J. Spiny lobster sounds can be detectable over kilometres underwater. Sci. Rep. 2020, 10, 7943. [Google Scholar] [CrossRef]
- Schmitz, B. Sound Production in Crustacea with Special Reference to the Alpheidae. In The Crustacean Nervous System; Springer: Berlin/Heidelberg, Germany, 2001; pp. 521–533. [Google Scholar]
- Radford, C.; Jeffs, A.; Tindle, C.; Montgomery, J.C. Resonating sea urchin skeletons create coastal choruses. Mar. Ecol. Prog. Ser. 2008, 362, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Di Iorio, L.; Gervaise, C.; Jaud, V.; Robson, A.A.; Chauvaud, L. Hydrophone detects cracking sounds: Non-intrusive monitoring of bivalve movement. J. Exp. Mar. Biol. Ecol. 2012, 432, 9–16. [Google Scholar] [CrossRef]
- Rice, A.N.; Farina, S.C.; Makowski, A.J.; Kaatz, I.M.; Lobel, P.S.; Bemis, W.E.; Bass, A.H. Evolutionary Patterns in Sound Production across Fishes. Ichthyol. Herpetol. 2022, 110, 1–12. [Google Scholar] [CrossRef]
- Parmentier, E.; Diogo, R.; Fine, M.L. Multiple exaptations leading to fish sound production. Fish Fish. 2017, 18, 958–966. [Google Scholar] [CrossRef]
- Amorim, M.C.P. Diversity of Sound Production in Fish; Communication in fishes; Science Publishers: Enfield, UK, 2006; pp. 71–104. [Google Scholar]
- Rako, N.; Picciulin, M.; Vilibić, I.; Fortuna, C.M. Spatial and temporal variability of sea ambient noise as an anthropogenic pressure index: The case of the Cres-Lošinj archipelago, Croatia. J. Mar. Biol. Assoc. U. K. 2013, 93, 27–36. [Google Scholar] [CrossRef]
- Putland, R.L.; Constantine, R.; Radford, C.A. Exploring spatial and temporal trends in the soundscape of an ecologically significant embayment. Sci. Rep. 2017, 7, 5713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolgan, M.; Gervaise, C.; Di Iorio, L.; Lossent, J.; Lejeune, P.; Raick, X.; Parmentier, E. Fish biophony in a Mediterranean submarine canyon. J. Acoust. Soc. Am. 2020, 147, 2466–2477. [Google Scholar] [CrossRef] [PubMed]
- Carriço, R.; Silva, M.A.; Menezes, G.M.; Fonseca, P.J.; Amorim, M.C.P. Characterization of the acoustic community of vocal fishes in the Azores. PeerJ 2019, 7, e7772. [Google Scholar] [CrossRef] [Green Version]
- Au, W.W.L. The Sonar of Dolphins; Springer: New York, NY, USA, 1993. [Google Scholar]
- Janik, V.M. Food–related bray calls in wild bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. Lond. B. Biol. Sci. 2000, 267, 923–927. [Google Scholar] [CrossRef] [Green Version]
- Lammers, M.O.; Au, W.W.L.; Aubauer, R.; Nachtigall, P.E. A comparative analysis of the pulsed emissions of free-ranging hawaiian spinner dolphins (Stenella longirostris). In Echolocation in Bats and Dolphins; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 414–419. [Google Scholar]
- Simard, P.; Lace, N.; Gowans, S.; Quintana-Rizzo, E.; Kuczaj, S.A.; Wells, R.S.; Mann, D.A. Low frequency narrow-band calls in bottlenose dolphins (Tursiops truncatus): Signal properties, function, and conservation implications. J. Acoust. Soc. Am. 2011, 130, 3068–3076. [Google Scholar] [CrossRef]
- Picciulin, M.; Calcagno, G.; Sebastianutto, L.; Bonacito, C.; Codarin, A.; Costantini, M.; Ferrero, E.A. Diagnostics of nocturnal calls of Sciaena umbra (L., fam., Sciaenidae) in a nearshore Mediterranean marine riserve. Bioacoustics 2012, 22, 109–120. [Google Scholar] [CrossRef]
- Janik, V.M.; King, S.L.; Sayigh, L.S.; Wells, R.S. Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mamm. Sci. 2013, 29, 109–122. [Google Scholar] [CrossRef]
- Gridley, T.; Nastasi, A.; Kriesell, H.J.; Elwen, S.H. The acoustic repertoire of wild common bottlenosedolphins (Tursiops truncatus) in Walvis Bay, Namibia. Bioacoustics 2015, 24, 153–174. [Google Scholar] [CrossRef] [Green Version]
- Bohnenstiehl, D.R.; Lillis, A.; Eggleston, D.B. The Curious Acoustic Behavior of Estuarine Snapping Shrimp: Temporal Patterns of Snapping Shrimp Sound in Sub-Tidal Oyster Reef Habitat. PLoS ONE 2016, 11, e0143691. [Google Scholar] [CrossRef] [PubMed]
- Smott, S.; Monczak, A.; Miller, M.; Montie, E.W. Boat noise in an estuarine soundscape–A potential risk on the acoustic communication and reproduction of soniferous fish in the May River, South Carolina. Mar. Pollut. Bull. 2018, 133, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Underwater Acoustics—Terminology, ISO 18405:2017. 2017. Available online: https://www.iso.org/standard/62406.html (accessed on 15 November 2021).
- Verfuss, U.K.; Andersson, M.; Folegot, T.; Laanearu, J.; Matuschek, R.; Pajala, J.; Sigray, P.; Tegowski, J.; Tougaard, J. BIAS Standards for Noise Measurements. Background Information, Guidelines and Quality Assurance. Amended Version. 2015. Available online: https://biasproject.files.wordpress.com/2016/04/bias_standards_v5_final.pdf (accessed on 15 November 2021).
- Betke, K.; Folegot, T.; Matuschek, R.; Pajala, J.; Persson, L.; Tegowski, J.; Tougaard, J.; Wahlberg, M. BIAS Standards for Signal Processing. Aims, Processes and Recommendations; Amended version; Verfus, U.K., Sigray, P., Eds.; BIAS: 2015. Available online: https://biasproject.files.wordpress.com/2016/01/bias_sigproc_standards_v5_final.pdf (accessed on 15 November 2021).
- Kéver, L.; Boyle, K.S.; Dragičević, B.; Dulčić, J.; Casadevall, M.; Parmentier, E. Sexual dimorphism of sonic apparatus and extreme intersexual variation of sounds in Ophidion rochei (Ophidiidae): First evidence of a tight relationship between morphology and sound characteristics in Ophidiidae. Front. Zool. 2012, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Parmentier, E.; Bouillac, G.; Dragicevic, B.; Dulcic, J.; Fine, M.L. Call properties and morphology of the sound- producing organ in Ophidion rochei (Ophidiidae). J. Exp. Biol. 2010, 213, 3230–3236. [Google Scholar] [CrossRef] [Green Version]
- Schultz, K.W.; Cato, D.H.; Corkeron, P.J.; Bryden, M.M. Low frequency narrow-band sounds produced by bottlenose dolphins. Mar. Mamm. Sci. 1995, 11, 503–509. [Google Scholar] [CrossRef]
- Lilly, J.C.; Miller, A.M. Sounds emitted by the bottlenose dolphin: The audible emissions of captive dolphins under water or in air are remarkably complex and varied. Science 1961, 133, 1689–1693. [Google Scholar] [CrossRef]
- Gridley, T.; Berggren, P.; Cockcroft, V.G.; Janik, V.M. Whistle vocalizations of Indo-Pacific bottlenosedolphins (Tursiops aduncus) inhabiting the south-west Indian Ocean. J. Acoust. Soc. Am. 2012, 132, 4032–4040. [Google Scholar] [CrossRef] [Green Version]
- Watkins, W.A. The harmonic interval: Fact or artifact in spectral analysis of pulse trains. In Marine Bio-Acoustics; Tavolga, W.N., Ed.; Pergamon Press: Oxford, UK, 1967; pp. 15–42. [Google Scholar]
- Dos Santos, M.E.; Ferreira, A.J.; Harzen, S. Rhythmic sound sequences emitted by aroused bottlenosedolphins in the Sado Estuary, Portugal. In Sensory Systems of Aquatic Mammals; de Spil: Woerden, NL, USA, 1995; pp. 325–334. [Google Scholar] [CrossRef]
- Radford, C.A.; Jeffs, A.G.; Tindle, C.T.; Montgomery, J.C. Temporal patterns in ambient noise of biological origin from a shallow water temperate reef. Oecologia 2008, 156, 921–929. [Google Scholar] [CrossRef]
- Staaterman, E.; Paris, C.B.; DeFerrari, H.A.; Mann, D.A.; Rice, A.N.; D’Alessandro, E.K. Celestial patterns in marine soundscapes. Mar. Ecol. Progr. Ser. 2014, 508, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Ramcharitar, J.; Gannon, D.; Popper, A. Bioacoustics of fishes of the family Sciaenidae (croackers and drums). Trans. Am. Fish Soc. 2006, 135, 1409–1431. [Google Scholar] [CrossRef]
- Gottesman, B.L.; Francomano, D.; Zhao, Z.; Bellisario, K.; Ghadiri, M.; Broadhead, T.; Gasc, A.; Pijanowski, B.C. Acoustic monitoring reveals diversity and surprising dynamics in tropical freshwater soundscapes. Freshw. Biol. 2018, 65, 117–132. [Google Scholar] [CrossRef]
- Dulcic, J.; Matic, S.; Kraljevic, M.; Franicevic, M.; Lipej, L. New data on the cuskeel, Ophidion rochei, from the eastern Adriatic. J. Mar. Biol. Assoc. U. K. 2002, 82, 1045–1046. [Google Scholar] [CrossRef]
- Pallaoro, A.; Jardas, I. Ichthyological collection of the Institute of Oceanography and Fisheries in split (Croatia). Natura Croatica 1996, 3, 177–219. [Google Scholar]
- Nielsen, J.G.; Cohen, D.M.; Markle, D.F.; Robins, C.R. Ophidiiform Fishes of the World (Order Ophidiiformes). An Annotated and Illustrated Catalogue of Pearlfishes, Cusk-eels, Brotulas and Other Ophidiiform Fishes Known to Date; FAO species catalogue. FAO Fisheries Synopsis; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999; Volume 18, ISBN 92-5-104375-2. [Google Scholar]
- Kéver, L.; Boyle, K.S.; Parmentier, E. Effects of seawater temperature on sound characteristics in Ophidion rochei (Ophidiidae). J. Fish Biol. 2015, 87, 502–509. [Google Scholar] [CrossRef]
- Kéver, L.; Lejeune, P.; Michel, L.N.; Parmentier, E. Passive acoustic recording of Ophidion rochei calling activity in Calvi Bay (France). Mar. Ecol. 2016, 37, 1315–1324. [Google Scholar] [CrossRef]
- Kovačic, M. Ichthyological collection (Cyclostomata, Selachii, Osteichthyes) of the Natural History Muesum Rijeka. In Natural History Researches of the Rijeka Region; Arko-Pijevac, M., Kovačić, M., Crnković, D., Eds.; Prirodoslovni Muzej u Rijeci: Rijeka, Croatia, 1998; pp. 685–698. [Google Scholar]
- Finneran, J.J. Dolphin “packet” use during long-range echolocation tasks. J. Acoust. Soc. Am. 2013, 133, 1796–1810. [Google Scholar] [CrossRef]
- Sprčić, A. Prehrana Dobrog Dupina (Tursiops truncatus, Montagu 1821) u Sjeveroistočnom Jadranu. Doctoral Dissertation, University of Zagreb, Faculty of Science, Department of Biology, Zagreb, Croatia, 2011. [Google Scholar]
- Janik, V.M.; Sayigh, L.S.; Wells, R.S. Signature whistle shape conveys identity information to bottlenose dolphins. Proc. Natl. Acad. Sci. USA 2006, 103, 8293–8297. [Google Scholar] [CrossRef] [Green Version]
- Janik, V.M.; Slater, P.J.B. Context-specific use suggests that bottlenose dolphin signature whistles are cohesion calls. Anim. Behav. 1998, 56, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Rako-Gospić, N.; Radulović, M.; Vučur, T.; Pleslić, G.; Holcer, D.; Mackelworth, P. Factor associated variations in the home range of a resident Adriatic common bottlenose dolphin population. Mar. Pollut. Bull. 2017, 124, 234–244. [Google Scholar] [CrossRef]
- Pleslić, G.; Rako-Gospić, N.; Miočić-Stošić, J.; BlazinićVučur, T.; Radulović, M.; Mackelworth, P.; Frleta-Valić, M.; Holcer, D. Social structure and spatial distribution of bottlenose dolphins (Tursiops truncatus) along the Croatian Adriatic coast. Aquat. Conserv. 2019, 29, 2116–2132. [Google Scholar] [CrossRef]
- Pirotta, E.; Laesser, B.E.; Hardaker, A.; Riddoch, N.; Marcoux, M.; Lusseau, D. Dredging displaces bottlenose dolphins from an urbanised foraging patch. Mar. Pollut. Bull. 2013, 74, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Marley, S.A.; Salgado-Kent, C.P.; Erbe, C.; Parnum, I. Effects of vessel traffic and underwater noise on the movement, behaviour and vocalisations of bottlenose dolphins in an urbanised estuary. Sci. Rep. 2017, 7, 13437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malfante, M.; Dalla Mura, M.; Mars, J.I.; Gervaise, C. Automatic fish sounds classification. J. Acoust. Soc. Am. 2018, 139, 2115–2116. [Google Scholar] [CrossRef] [Green Version]
- Monczak, A.; Ji, Y.; Soueidan, J.; Montie, E.W. Automatic detection, classification, and quantification of sciaenid fish calls in an estuarine soundscape in the Southeast United States. PLoS ONE 2019, 14, e0209914. [Google Scholar] [CrossRef]
- Vieira, M.; Pereira, B.P.; Pousão-Ferreira, P.; Fonseca, P.J.; Amorim, M. Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach. Fishes 2019, 4, 28. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. Pulses | 1 Peak Freq (Hz) | 2 Peak Freq (Hz) | Sound Duration (ms) | Alternation Start | Long Pulse Period (ms) | Short Pulse Period (ms) | ||
|---|---|---|---|---|---|---|---|---|
| Lošinj data (N = 20) | mean | 36.7 | 241.8 | 384.1 | 4144.4 | 7.9 | 129.4 | 103.1 |
| SD | 5.8 | 26.7 | 45.1 | 727.7 | 2.6 | 13.7 | 10.9 | |
| c.v. (%) | 15.8 | 11.0 | 11.7 | 17.5 | 33.7 | 10.6 | 10.6 |
| 5% Freq (Hz) | Center Freq (Hz) | Peak Freq (Hz) | 95% Freq (Hz) | Sound Duration (ms) | Number of Trains | Train Duration (ms) | Train Period (ms) | ||
|---|---|---|---|---|---|---|---|---|---|
| Lošinj data (N = 15) | mean | 161.4 | 679.6 | 610.4 | 1406 | 1698 | 5.9 | 88.3 | 236.1 |
| SD | 16.8 | 116.1 | 207 | 16.2 | 72.8 | 0.2 | 23.3 | 73.2 | |
| c.v. (%) | 10.4 | 17.8 | 33.9 | 1.1 | 4.3 | 3.4 | 26.4 | 31.0 |
| Whistle | Chirp | Burst Pulse | Low Frequency Narrow-Band Calls | Bray | ||
|---|---|---|---|---|---|---|
| NTS | Day | 16 | 7 | 0 | 41 | 0 |
| NTS | Night | 12 | 1 | 0 | 19 | 0 |
| Total | 28 | 8 | 0 | 60 | 0 | |
| TS | Day | 41 | 22 | 9 | 196 | 0 |
| TS | Night | 16 | 0 | 1 | 0 | 0 |
| Total | 57 | 22 | 10 | 196 | 0 | |
| Fish Sounds * | Boat Noise | Dolphin Sounds (Whistles, Chirp, Burst Pulse, Echolocation Clicks) | Dolphin Sounds (Low Frequency Narrow-Band Calls) | |
|---|---|---|---|---|
| NTS | 6.7% ± 1.4 | 42.5% ± 2 | 1.4% ± 0.3 | 0.3% ± 1.4 |
| TS | 27.3% ± 3.5 | 37% ± 1.4 | 1.8% ± 0.5 | 0.4% ± 1.5 |
| Mann-Whitney U | 5623.5 | 6586.5 | 7156 | 6907 |
| P | 0.001 | 0.252 | 0.903 | 0.221 |
| Fish Sounds | Boat Noises | LFN Dolphin Sounds | |
|---|---|---|---|
| SPLsLF—TS | rs = −0.40, p-value = 0.00 | rs = 0.43, p-value = 0.00 | rs = 0.18, p-value = 0.04 |
| SPLsLF—NTS | rs = 0.023, p-value = 0.79 | rs = 0.039, p-value = 0.67 | rs = −0.06, p-value = 0.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picciulin, M.; Bolgan, M.; Rako-Gospić, N.; Petrizzo, A.; Radulović, M.; Falkner, R. A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia). J. Mar. Sci. Eng. 2022, 10, 300. https://doi.org/10.3390/jmse10020300
Picciulin M, Bolgan M, Rako-Gospić N, Petrizzo A, Radulović M, Falkner R. A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia). Journal of Marine Science and Engineering. 2022; 10(2):300. https://doi.org/10.3390/jmse10020300
Chicago/Turabian StylePicciulin, Marta, Marta Bolgan, Nikolina Rako-Gospić, Antonio Petrizzo, Marko Radulović, and Raffaela Falkner. 2022. "A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia)" Journal of Marine Science and Engineering 10, no. 2: 300. https://doi.org/10.3390/jmse10020300
APA StylePicciulin, M., Bolgan, M., Rako-Gospić, N., Petrizzo, A., Radulović, M., & Falkner, R. (2022). A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia). Journal of Marine Science and Engineering, 10(2), 300. https://doi.org/10.3390/jmse10020300