
Spatial Distribution of Colored Dissolved Organic Matter in the Western Arctic Ocean

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
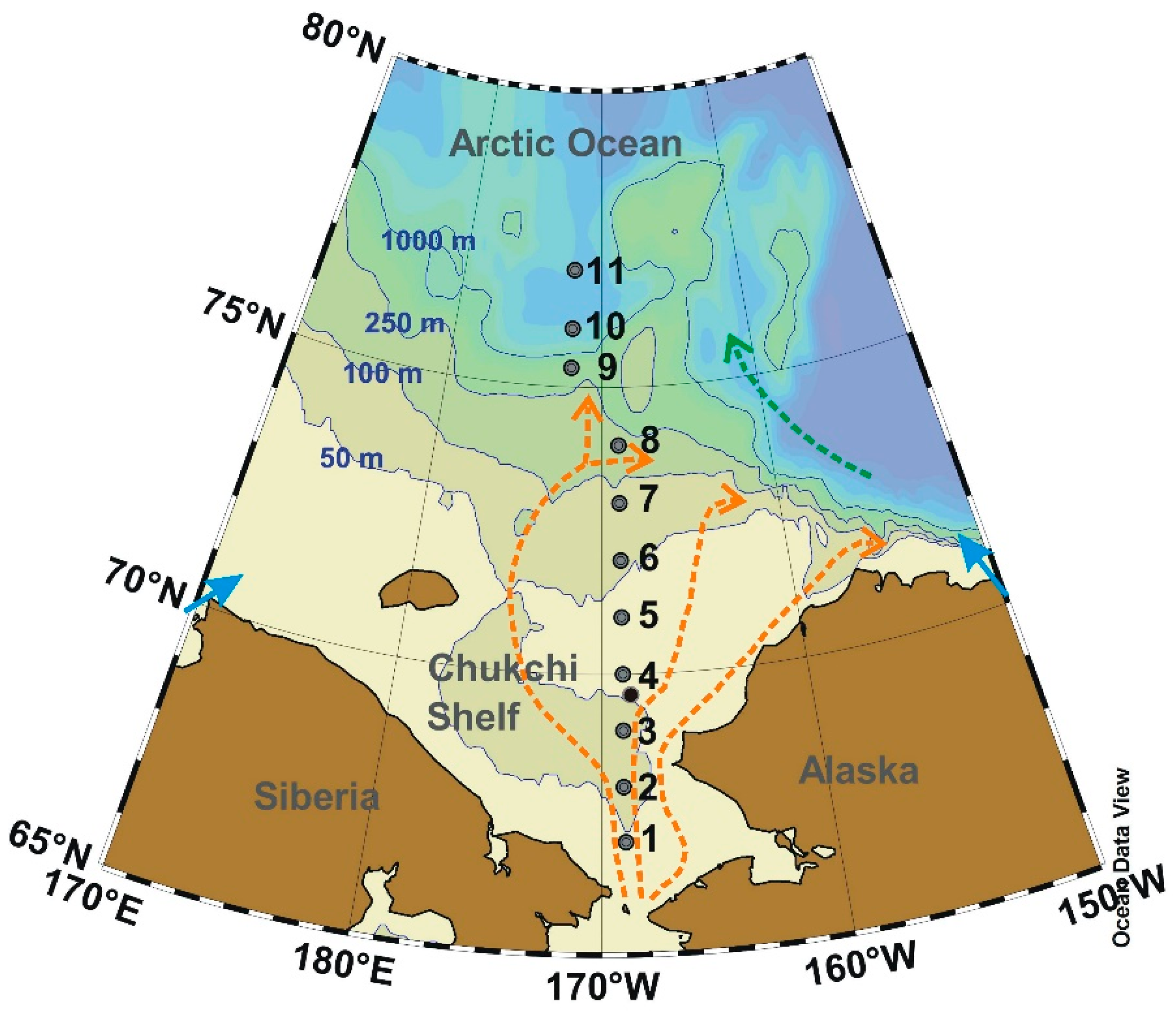
2.1. Study Sites and Sampling
2.2. Absorption Spectroscopy Analysis
2.3. EEM and PARAFAC Modeling
3. Results
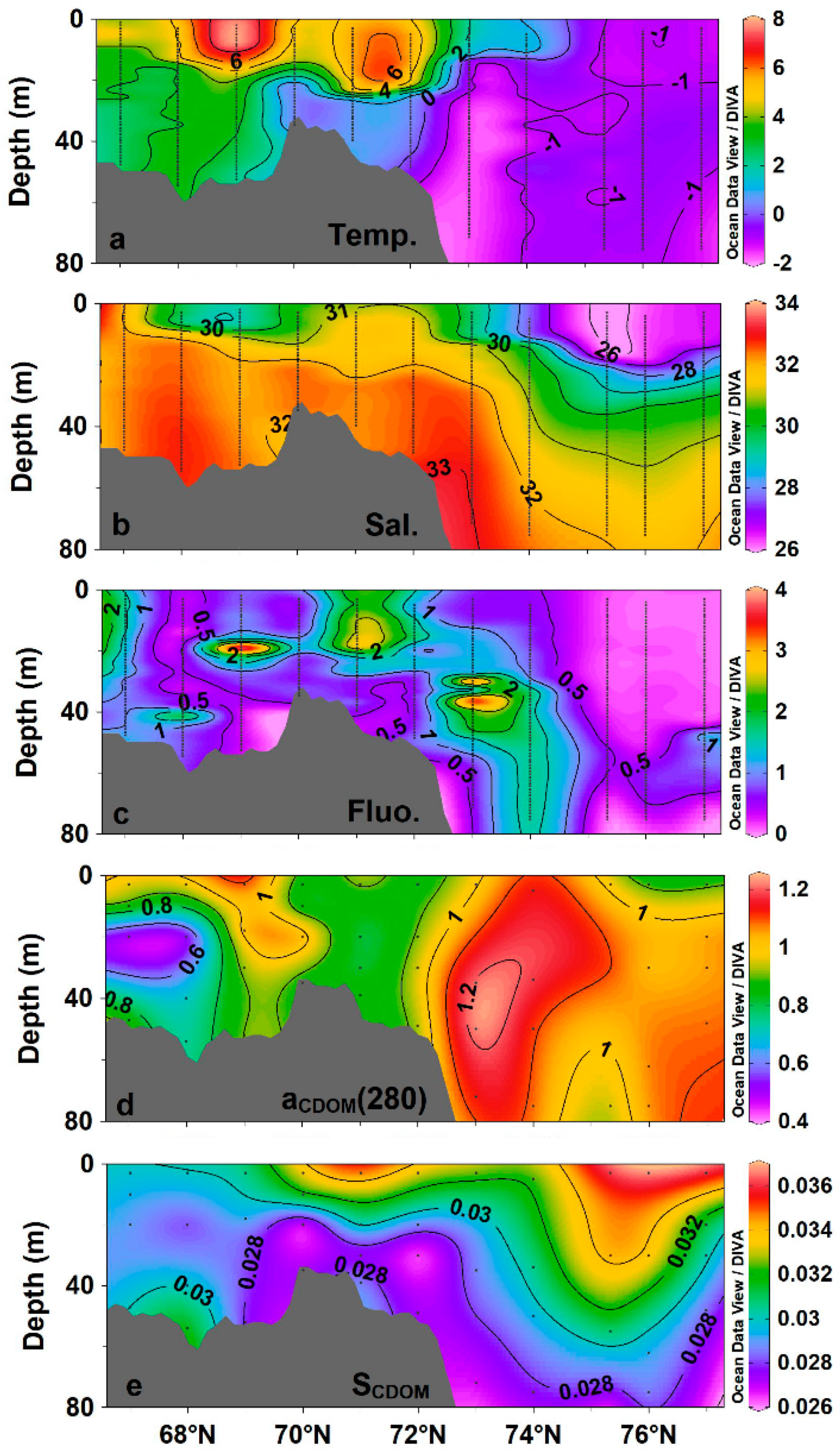
3.1. Variability of Environmental Factors and CDOM Absorption Properties
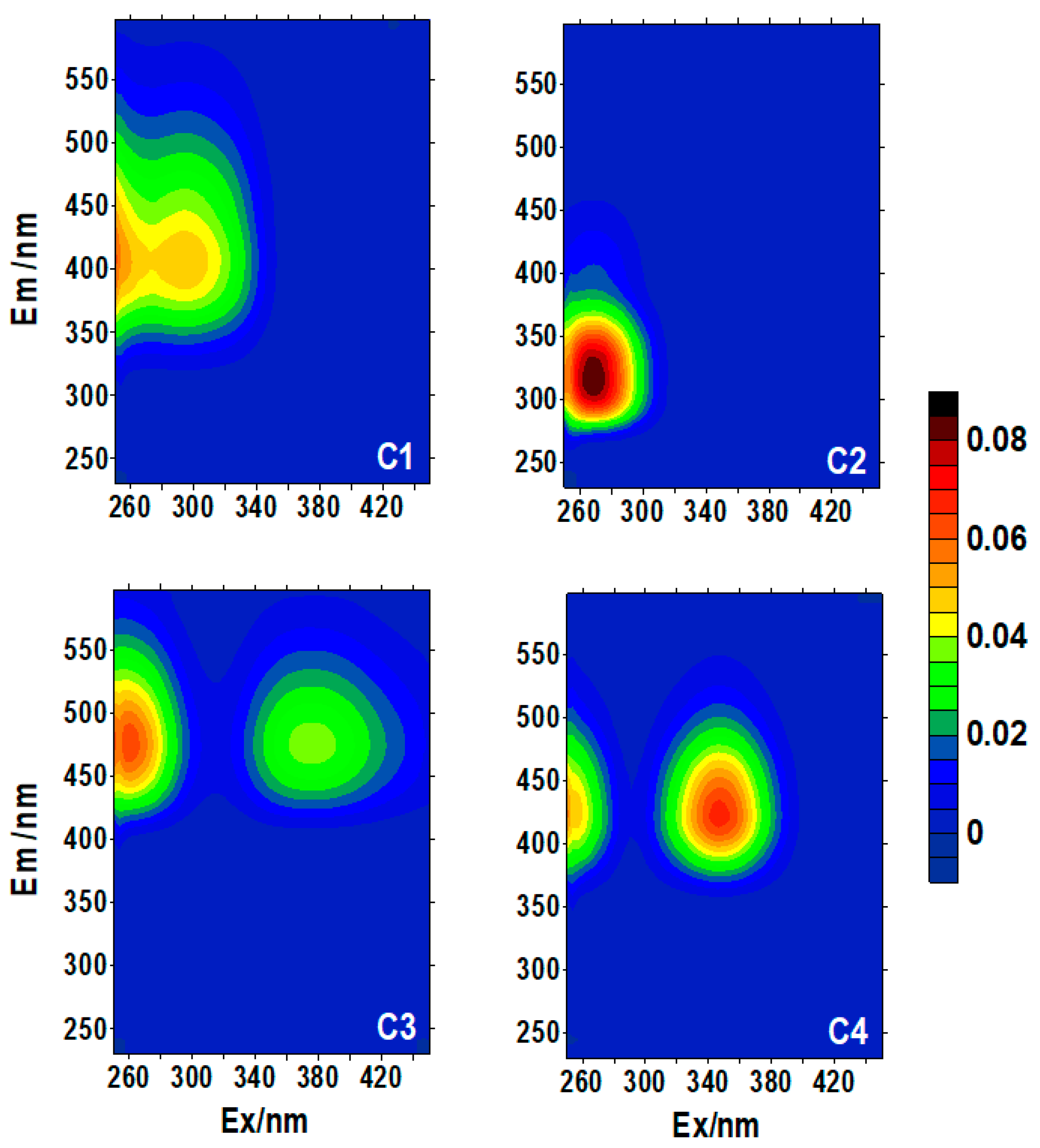
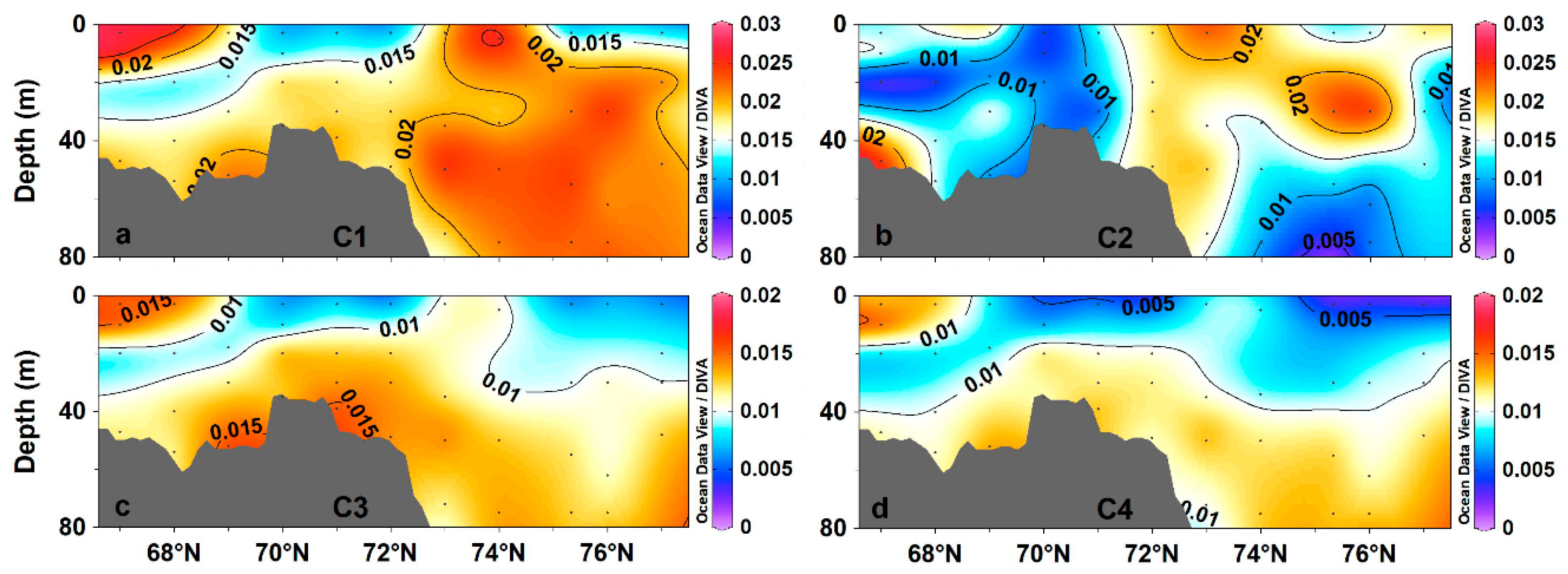
3.2. Fluorescent Component Characteristics
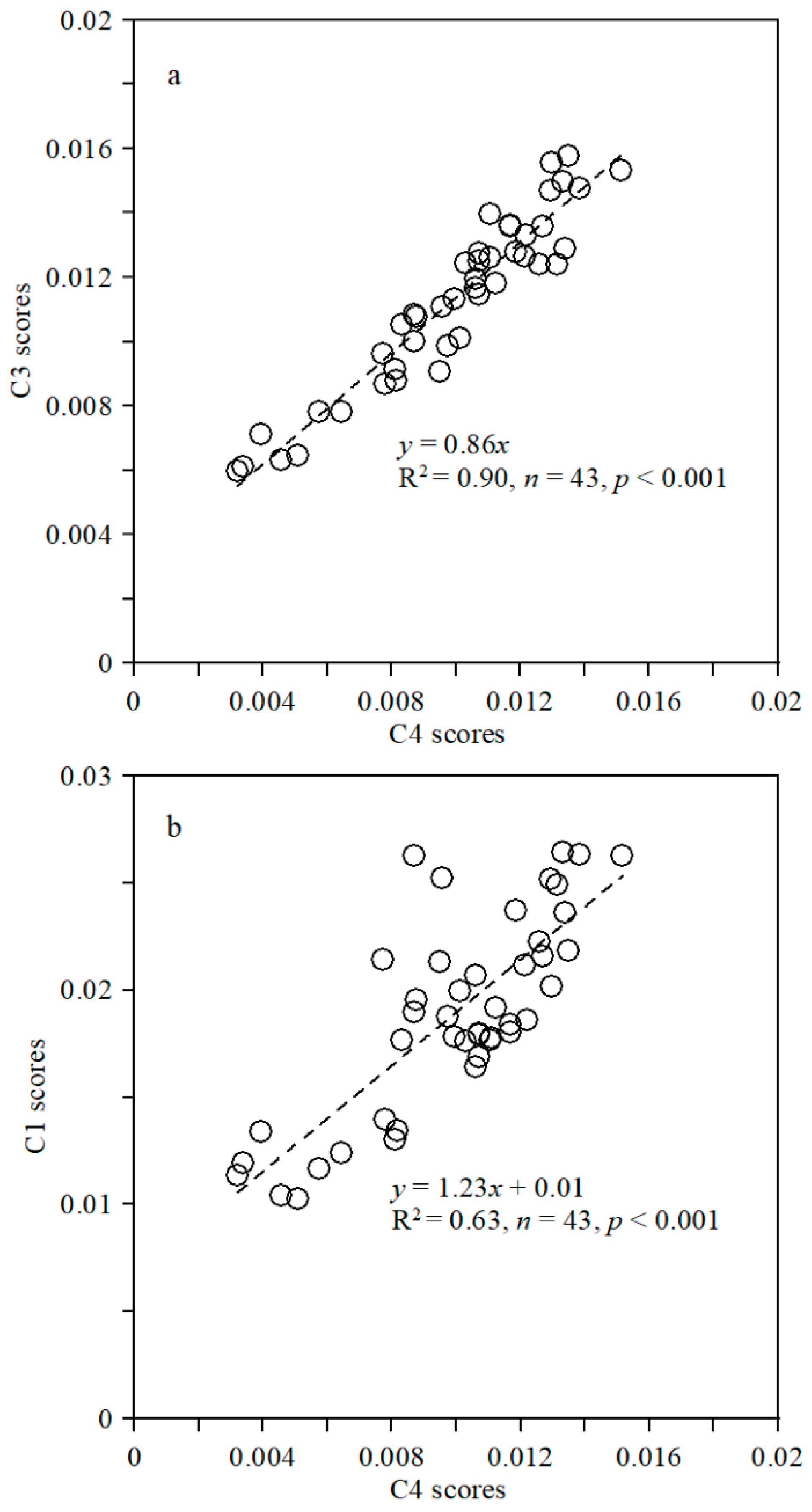
3.3. Variability in CDOM Fluorescent Components
4. Discussion
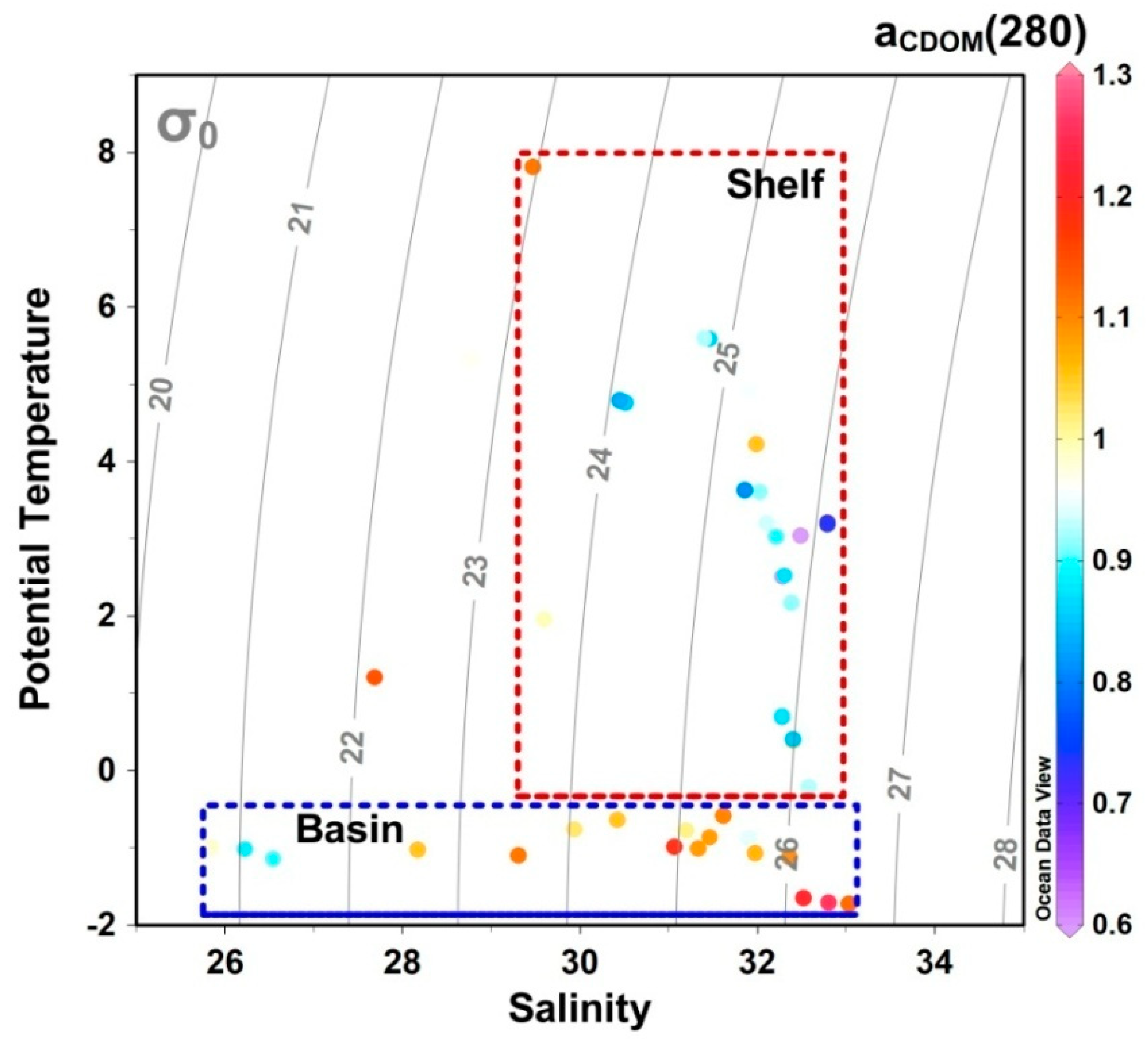
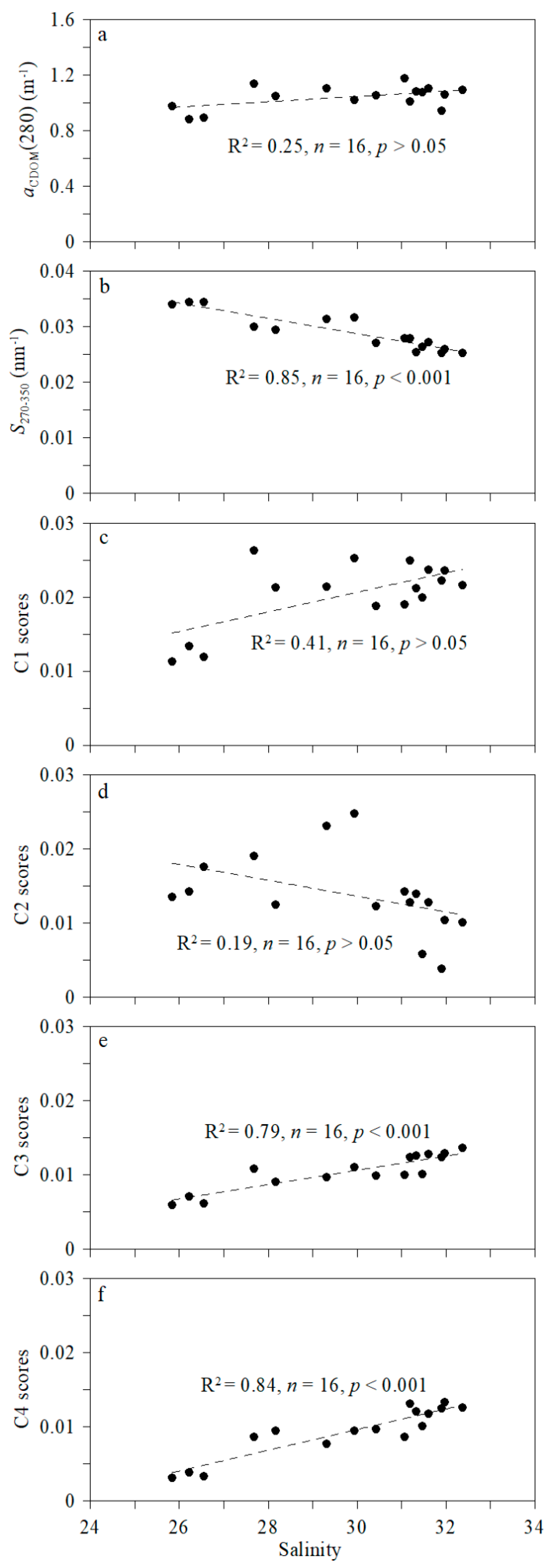
4.1. Shelf–Basin Difference in CDOM Concentration
4.2. Effects of Ice Melting and Pacific Inflow on CDOM Fluorescence
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jiao, N.; Herndl, G.J.; Hansell, D.A.; Benner, R.; Kattner, G.; Wilhelm, S.W.; Kirchman, D.L.; Weinbauer, M.G.; Luo, T.; Chen, F.; et al. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Genet. 2010, 8, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Benner, R. Chemical Composition and Reactivity. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: New York, NY, USA, 2002; pp. 59–90. [Google Scholar]
- Stedmon, C.A.; Nelson, N.B. The Optical Properties of DOM in the Ocean. In Biogeochemistry of Marine Dissolved Organic Matter; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 481–508. [Google Scholar]
- Chen, M.; Kim, J.-H.; Nam, S.-I.; Niessen, F.; Hong, W.-L.; Kang, M.-H.; Hur, J. Production of fluorescent dissolved organic matter in Arctic Ocean sediments. Sci. Rep. 2016, 6, 39213. [Google Scholar] [CrossRef] [Green Version]
- Brogi, S.R.; Charrière, B.; Gonnelli, M.; Vaultier, F.; Sempéré, R.; Vestri, S.; Santinelli, C. Effect of UV and Visible Radiation on Optical Properties of Chromophoric Dissolved Organic Matter Released by Emiliania huxleyi. J. Mar. Sci. Eng. 2020, 8, 888. [Google Scholar] [CrossRef]
- Rabe, B.; Karcher, M.; Kauker, F.; Schauer, U.; Toole, J.; Krishfield, R.A.; Pisarev, S.; Kikuchi, T.; Su, J. Arctic Ocean basin liquid freshwater storage trend 1992–2012. Geophys. Res. Lett. 2014, 41, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Giles, K.A.; Laxon, S.W.; Ridout, A.L.; Wingham, D.J.; Bacon, S. Western Arctic Ocean freshwater storage increased by wind-driven spin-up of the Beaufort Gyre. Nat. Geosci. 2012, 5, 194–197. [Google Scholar] [CrossRef]
- Woodgate, R.; Peralta-Ferriz, C. Warming and Freshening of the Pacific Inflow to the Arctic from 1990–2019 Implying Dramatic Shoaling in Pacific Winter Water Ventilation of the Arctic Water Column. Geophys. Res. Lett. 2021, 48, e2021GL092528. [Google Scholar] [CrossRef]
- Timmermans, M.-L.; Marshall, J.; Proshutinsky, A.; Scott, J. Seasonally derived components of the Canada Basin halocline. Geophys. Res. Lett. 2017, 44, 5008–5015. [Google Scholar] [CrossRef]
- Nicolaus, M.; Katlein, C.; Maslanik, J.; Hendricks, S. Changes in Arctic sea ice result in increasing light transmittance and absorption. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, Y.; Jin, H.; Cai, W.-J.; Li, H.; Jin, M.; Qi, D.; Chen, J. Freshening leads to a three-decade trend of declining nutrients in the western Arctic Ocean. Environ. Res. Lett. 2021, 16, 054047. [Google Scholar] [CrossRef]
- Zhang, M.; Marandino, C.A.; Yan, J.; Wu, Y.; Park, K.; Sun, H.; Gao, Z.; Xu, S. Unravelling Surface Seawater DMS Concentration and Sea-To-Air Flux Changes After Sea Ice Retreat in the Western Arctic Ocean. Glob. Biogeochem. Cycles 2021, 35. [Google Scholar] [CrossRef]
- Terhaar, J.; Orr, J.C.; Ethé, C.; Regnier, P.; Bopp, L. Simulated Arctic Ocean Response to Doubling of Riverine Carbon and Nutrient Delivery. Glob. Biogeochem. Cycles 2019, 33, 1048–1070. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K.M.; van Dijken, G.L.; Arrigo, K.R. Changes in phytoplankton concentration now drive increased Arctic Ocean primary production. Science 2020, 369, 198–202. [Google Scholar] [CrossRef]
- Zhuang, Y.; Jin, H.; Chen, J.; Li, H.; Ji, Z.; Bai, Y.; Zhang, T. Nutrient and phytoplankton dynamics driven by the Beaufort Gyre in the western Arctic Ocean during the period 2008–2014. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2018, 137, 30–37. [Google Scholar] [CrossRef]
- Bates, N.R.; Moran, B.; Hansell, D.; Mathis, J.T. An increasing CO2sink in the Arctic Ocean due to sea-ice loss. Geophys. Res. Lett. 2006, 33, 23609. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.-J.; Chen, L.; Chen, B.; Gao, Z.; Lee, S.H.; Chen, J.; Pierrot, D.; Sullivan, K.; Wang, Y.; Hu, X.; et al. Decrease in the CO 2 Uptake Capacity in an Ice-Free Arctic Ocean Basin. Science 2010, 329, 556–559. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Nam, S.-I.; Kim, J.-H.; Kwon, Y.-J.; Hong, S.; Jung, J.; Shin, K.-H.; Hur, J. High abundance of protein-like fluorescence in the Amerasian Basin of Arctic Ocean: Potential implication of a fall phytoplankton bloom. Sci. Total. Environ. 2017, 599–600, 355–363. [Google Scholar] [CrossRef]
- DeFrancesco, C.; Guéguen, C. Long-term Trends in Dissolved Organic Matter Composition and Its Relation to Sea Ice in the Canada Basin, Arctic Ocean (2007–2017). J. Geophys. Res. Oceans 2021, 126, e2020JC016578. [Google Scholar] [CrossRef]
- Shen, Y.; Benner, R.; Kaiser, K.; Fichot, C.; Whitledge, T.E. Pan-Arctic Distribution of Bioavailable Dissolved Organic Matter and Linkages with Productivity in Ocean Margins. Geophys. Res. Lett. 2018, 45, 1490–1498. [Google Scholar] [CrossRef]
- Stedmon, C.; Markager, S.; Kaas, H. Optical Properties and Signatures of Chromophoric Dissolved Organic Matter (CDOM) in Danish Coastal Waters. Estuar. Coast. Shelf Sci. 2000, 51, 267–278. [Google Scholar] [CrossRef]
- Guo, W.; Stedmon, C.; Han, Y.; Wu, F.; Yu, X.; Hu, M. The conservative and non-conservative behavior of chromophoric dissolved organic matter in Chinese estuarine waters. Mar. Chem. 2007, 107, 357–366. [Google Scholar] [CrossRef]
- Yushmanova, A.; Kopelevich, O.; Vazyulya, S.; Sahling, I. Inter-Annual Variability of the Seawater Light Absorption in Surface Layer of the Northeastern Black Sea in Connection with Hydrometeorological Factors. J. Mar. Sci. Eng. 2019, 7, 326. [Google Scholar] [CrossRef] [Green Version]
- Stedmon, C.A.; Markager, S. Tracing the production and degradation of autochthonous fractions of dissolved organic matter by fluorescence analysis. Limnol. Oceanogr. 2005, 50, 1415–1426. [Google Scholar] [CrossRef]
- Guo, W.; Yang, L.; Hong, H.; Stedmon, C.; Wang, F.; Xu, J.; Xie, Y. Assessing the dynamics of chromophoric dissolved organic matter in a subtropical estuary using parallel factor analysis. Mar. Chem. 2011, 124, 125–133. [Google Scholar] [CrossRef]
- Osburn, C.L.; Handsel, L.T.; Mikan, M.P.; Paerl, H.W.; Montgomery, M. Fluorescence Tracking of Dissolved and Particulate Organic Matter Quality in a River-Dominated Estuary. Environ. Sci. Technol. 2012, 46, 8628–8636. [Google Scholar] [CrossRef]
- Markager, S.; Vincent, W.F. Spectral light attenuation and the absorption of UV and blue light in natural waters. Limnol. Oceanogr. 2000, 45, 642–650. [Google Scholar] [CrossRef]
- Stedmon, C.A.; Bro, R. Characterizing dissolved organic matter fluorescence with parallel factor analysis: A tutorial. Limnol. Oceanogr. Meth. 2008, 6, 572–579. [Google Scholar] [CrossRef]
- Coble, P.G.; Del Castillo, C.E.; Avril, B. Distribution and optical properties of CDOM in the Arabian Sea during the 1995 Southwest Monsoon. Deep Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 2195–2223. [Google Scholar] [CrossRef]
- Jia, R.; Mu, X.; Chen, M.; Zhu, J.; Wang, B.; Li, X.; Astakhov, A.S.; Zheng, M.; Qiu, Y. Sources of particulate organic matter in the Chukchi and Siberian shelves: Clues from carbon and nitrogen isotopes. Acta Oceanol. Sin. 2020, 39, 96–108. [Google Scholar] [CrossRef]
- Gueguen, C.; Guo, L.; Yamamoto-Kawai, M.; Tanaka, N. Colored dissolved organic matter dynamics across the shelf-basin interface in the western Arctic Ocean. J. Geophys. Res. Space Phys. 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Fichot, C.G.; Benner, R. Dissolved organic matter composition and bioavailability reflect ecosystem productivity in the Western Arctic Ocean. Biogeosciences 2012, 9, 4993–5005. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Yang, L.; Zhai, W.; Chen, W.; Osburn, C.L.; Huang, X.; Li, Y. Runoff-mediated seasonal oscillation in the dynamics of dissolved organic matter in different branches of a large bifurcated estuary-The Changjiang Estuary. J. Geophys. Res. Biogeosci. 2014, 119, 776–793. [Google Scholar] [CrossRef]
- Chen, M.; Huang, Y.; Guo, L.; Cai, P.; Yang, W.; Liu, G.; Qiu, Y. Biological productivity and carbon cycling in the Arctic Ocean. Chin. Sci. Bull. 2002, 47, 1037–1040. [Google Scholar] [CrossRef]
- Zhuang, Y.; Jin, H.; Li, H.; Chen, J.; Lin, L.; Bai, Y.; Ji, Z.; Zhang, Y.; Gu, F. Pacific inflow control on phytoplankton community in the Eastern Chukchi Shelf during summer. Cont. Shelf Res. 2016, 129, 23–32. [Google Scholar] [CrossRef]
- Anderson, L.G.; Amon, R.M. DOM in the Arctic Ocean. In Biogeochemistry of Marine Dissolved Organic Matter; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 609–633. [Google Scholar]
- Zhuang, Y.; Jin, H.; Chen, J.; Ren, J.; Zhang, Y.; Lan, M.; Zhang, T.; He, J.; Tian, J. Phytoplankton Community Structure at Subsurface Chlorophyll Maxima on the Western Arctic Shelf: Patterns, Causes, and Ecological Importance. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005570. [Google Scholar] [CrossRef]
- Massi, L.; Frittitta, L.; Melillo, C.; Polonelli, F.; Bianchi, V.; De Biasi, A.M.; Nuccio, C. Seasonal Dynamic of CDOM in a Shelf Site of the South-Eastern Ligurian Sea (Western Mediterranean). J. Mar. Sci. Eng. 2020, 8, 703. [Google Scholar] [CrossRef]
- Li, W.K.W.; McLaughlin, F.A.; Lovejoy, C.; Carmack, E.C. Smallest Algae Thrive as the Arctic Ocean Freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Y.; Jin, H.; Zhang, Y.; Li, H.; Zhang, T.; Li, Y.; Bai, Y.; Ren, J.; Chen, J. Incursion of Alaska Coastal Water as a mechanism promoting small phytoplankton in the western Arctic Ocean. Prog. Oceanogr. 2021, 197, 102639. [Google Scholar] [CrossRef]
- Mathis, J.T.; Hansell, D.; Bates, N.R. Strong hydrographic controls on spatial and seasonal variability of dissolved organic carbon in the Chukchi Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 3245–3258. [Google Scholar] [CrossRef]
- Chen, M.; Jung, J.; Lee, Y.K.; Hur, J. Surface accumulation of low molecular weight dissolved organic matter in surface waters and horizontal off-shelf spreading of nutrients and humic-like fluorescence in the Chukchi Sea of the Arctic Ocean. Sci. Total. Environ. 2018, 639, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Brogi, S.R.; Ha, S.-Y.; Kim, K.; Derrien, M.; Lee, Y.K.; Hur, J. Optical and molecular characterization of dissolved organic matter (DOM) in the Arctic ice core and the underlying seawater (Cambridge Bay, Canada): Implication for increased autochthonous DOM during ice melting. Sci. Total. Environ. 2018, 627, 802–811. [Google Scholar] [CrossRef]
- Stedmon, C.; Amon, R.; Rinehart, A.; Walker, S. The supply and characteristics of colored dissolved organic matter (CDOM) in the Arctic Ocean: Pan Arctic trends and differences. Mar. Chem. 2011, 124, 108–118. [Google Scholar] [CrossRef]
- Zhuang, Y.; Li, H.; Jin, H.; Gao, S.; Chen, J.; Li, Y.; Bai, Y.; Ji, Z. Vertical distribution of nutrient tracers in the western Arctic Ocean and its relationship to water structure and biogeochemical processes. Acta Oceanol. Sin. 2020, 39, 109–114. [Google Scholar] [CrossRef]
- Nishino, S.; Shimada, K.; Itoh, M. Use of ammonium and other nitrogen tracers to investigate the spreading of shelf waters in the western Arctic halocline. J. Geophys. Res. Oceans 2005, 110, e2019JC015428. [Google Scholar] [CrossRef]
- Stedmon, C.A.; Amon, R.M.W.; Bauch, D.; Bracher, A.; Gonçalves-Araujo, R.; Hoppmann, M.; Krishfield, R.; Laney, S.; Rabe, B.; Reader, H.; et al. Insights into Water Mass Origins in the Central Arctic Ocean from In-Situ Dissolved Organic Matter Fluorescence. J. Geophys. Res. Oceans 2021, 126, e2021JC017407. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, X.; Zhuang, Y.; Li, H.; Xu, J.; Jin, H.; Chen, J. Spatial Distribution of Colored Dissolved Organic Matter in the Western Arctic Ocean. J. Mar. Sci. Eng. 2022, 10, 352. https://doi.org/10.3390/jmse10030352
Cai X, Zhuang Y, Li H, Xu J, Jin H, Chen J. Spatial Distribution of Colored Dissolved Organic Matter in the Western Arctic Ocean. Journal of Marine Science and Engineering. 2022; 10(3):352. https://doi.org/10.3390/jmse10030352
Chicago/Turabian StyleCai, Xiaoxia, Yanpei Zhuang, Hongliang Li, Jing Xu, Haiyan Jin, and Jianfang Chen. 2022. "Spatial Distribution of Colored Dissolved Organic Matter in the Western Arctic Ocean" Journal of Marine Science and Engineering 10, no. 3: 352. https://doi.org/10.3390/jmse10030352
APA StyleCai, X., Zhuang, Y., Li, H., Xu, J., Jin, H., & Chen, J. (2022). Spatial Distribution of Colored Dissolved Organic Matter in the Western Arctic Ocean. Journal of Marine Science and Engineering, 10(3), 352. https://doi.org/10.3390/jmse10030352