
Benthic Foraminifera and Productivity Regimes in the Kveithola Trough (Barents Sea)—Ecological Implications in a Changing Arctic and Actuopaleontological Meaning


 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
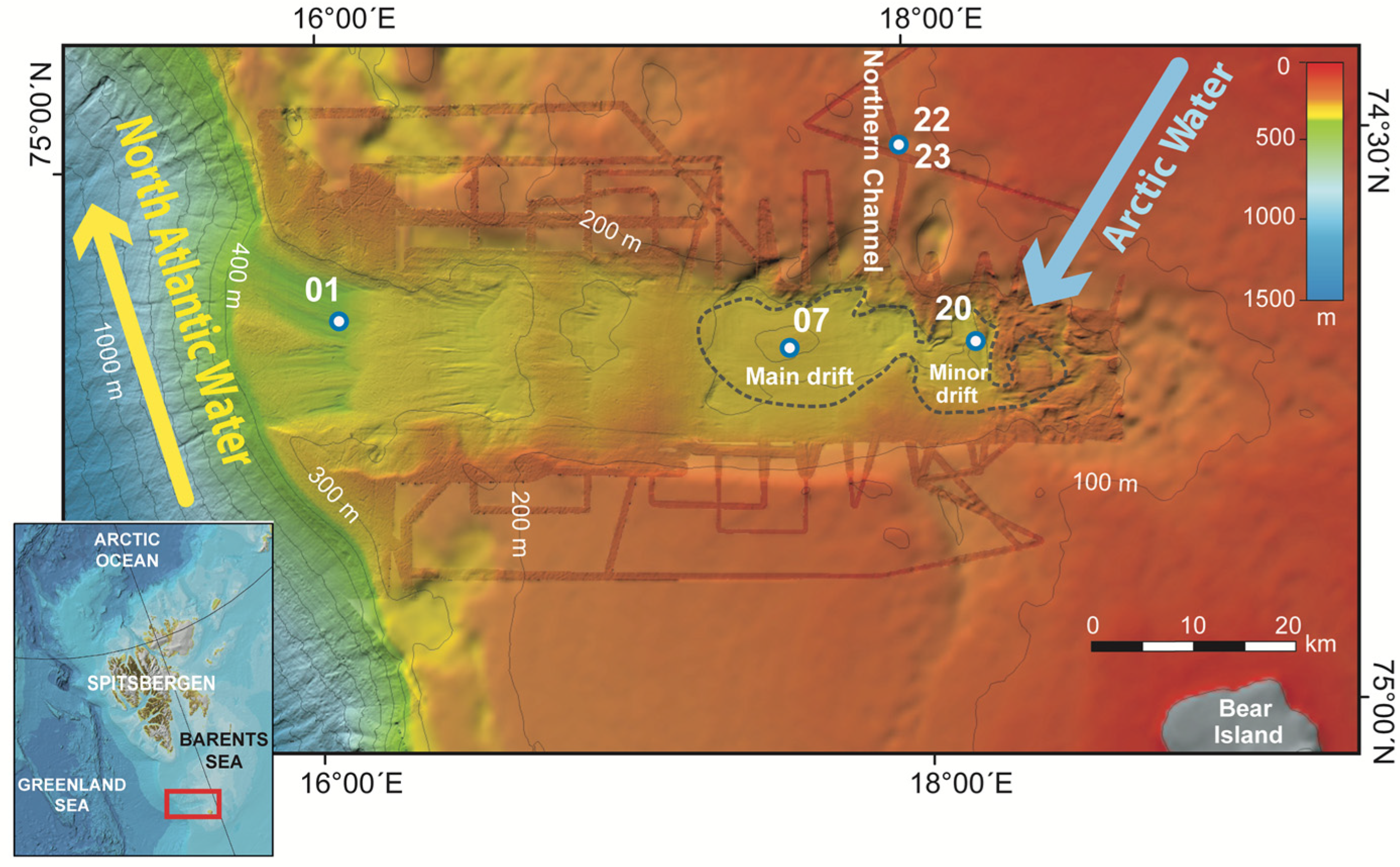
2.1. Study Area: The Kveithola Trough
2.2. Sampling Strategy
2.3. Sedimentological Analyses
2.4. Total Organic Carbon (TOC), Total Nitrogen (TN), and Biopolymeric Carbon (BPC)
2.5. Foraminiferal Analyses
2.6. Statistical Analyses
3. Results
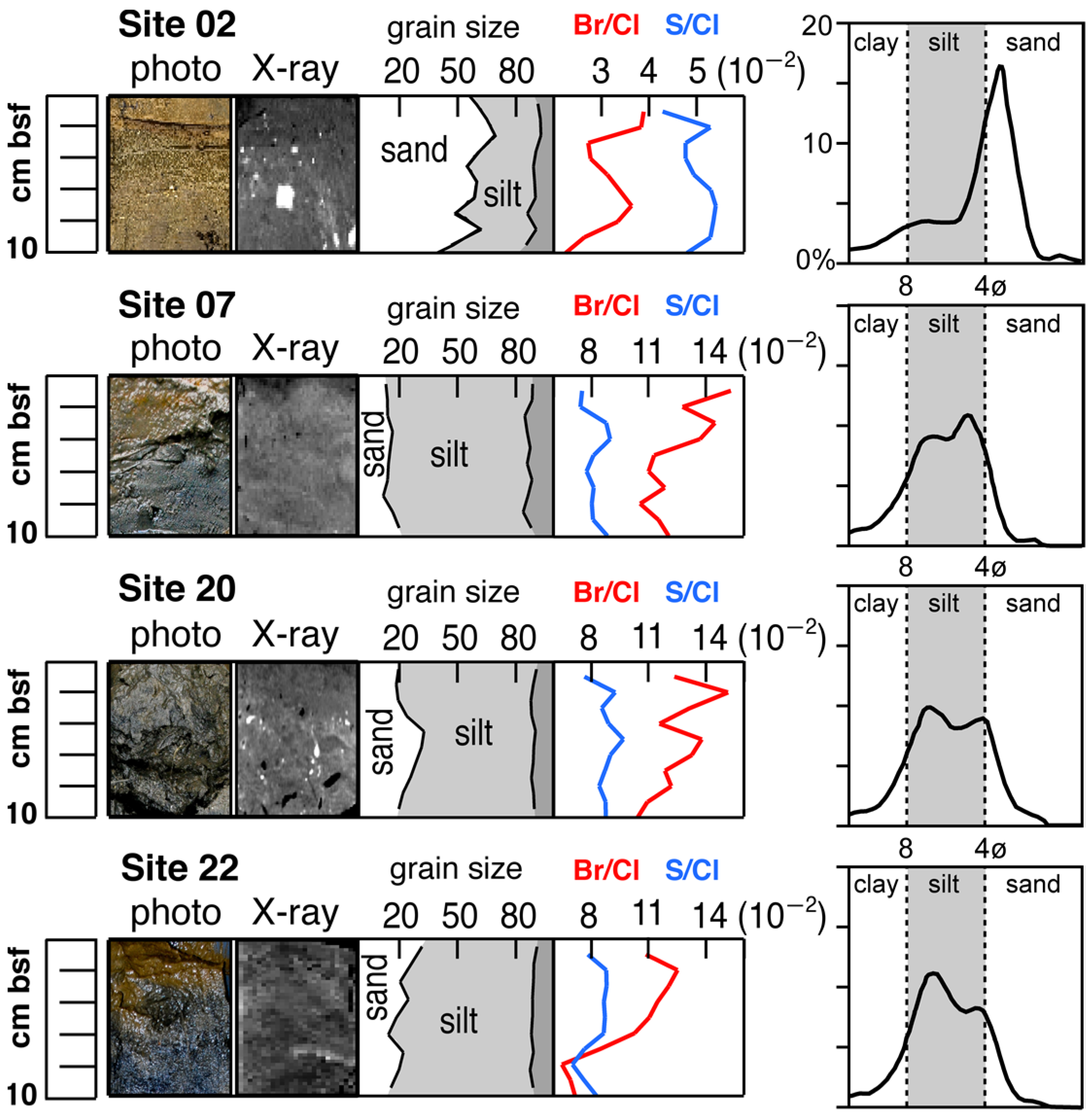
3.1. Sediment Description and Geochemistry
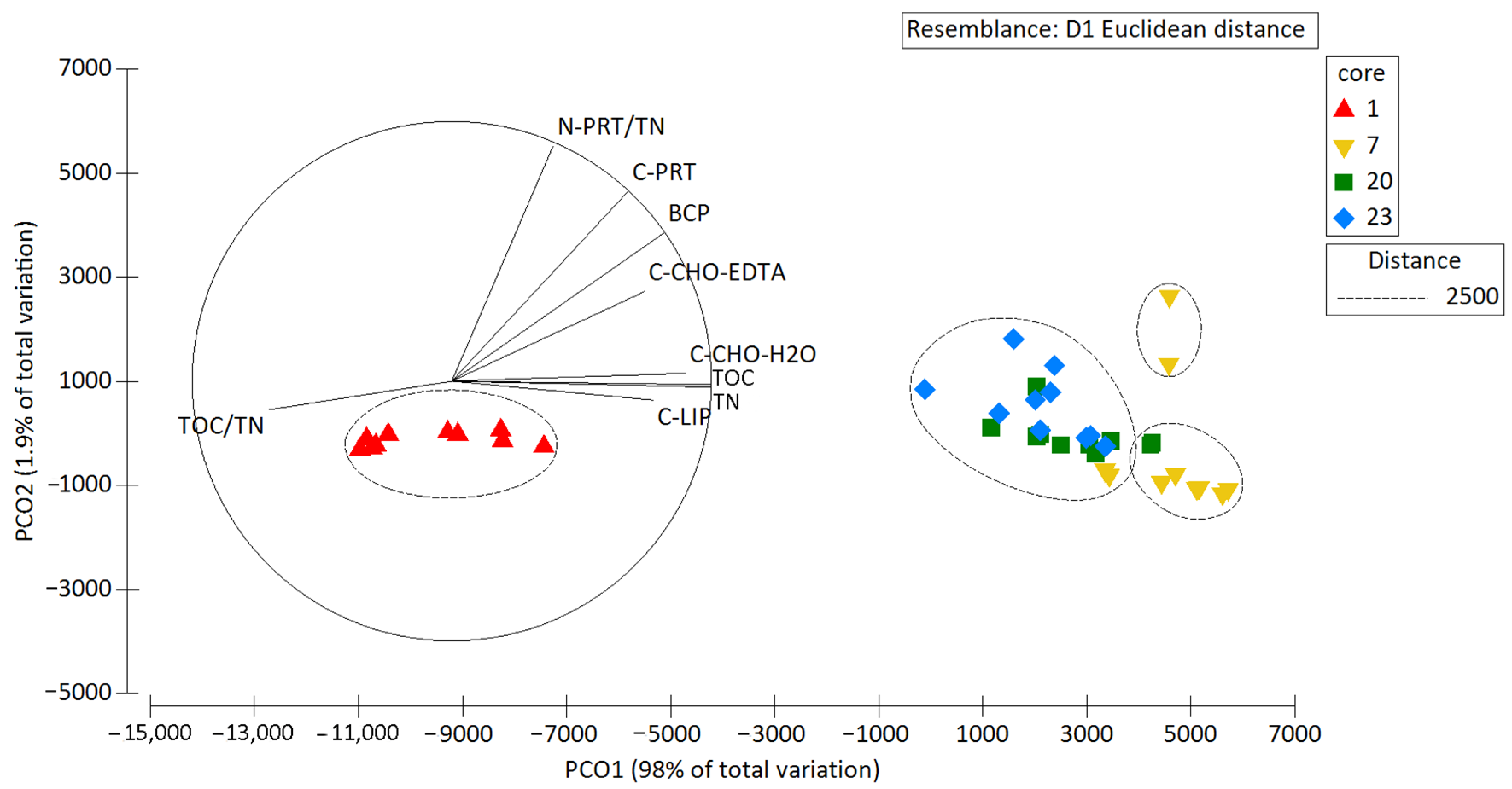
3.2. Chemical Characterization of Sediments
3.3. Foraminiferal Densities and Diversity
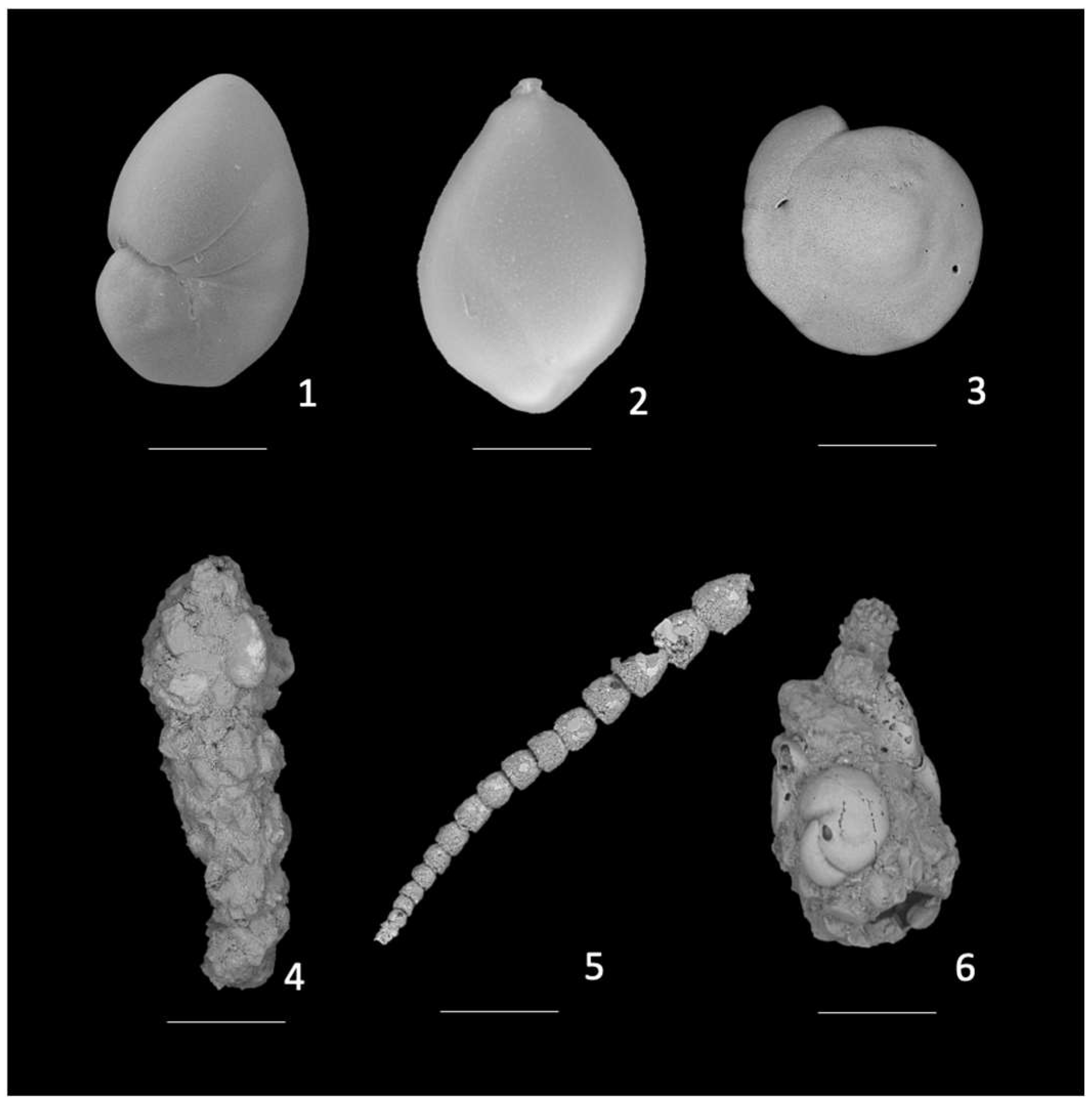
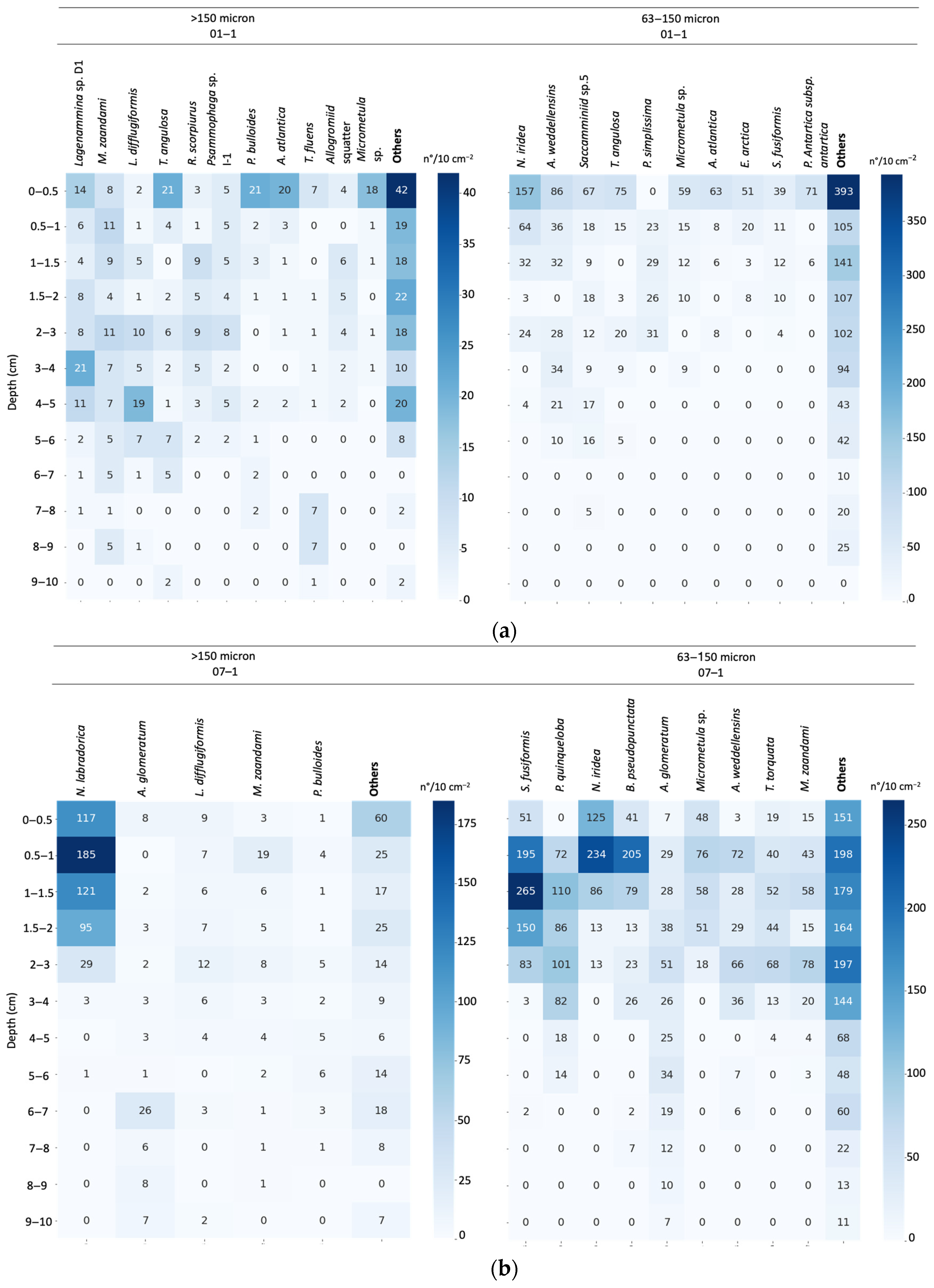
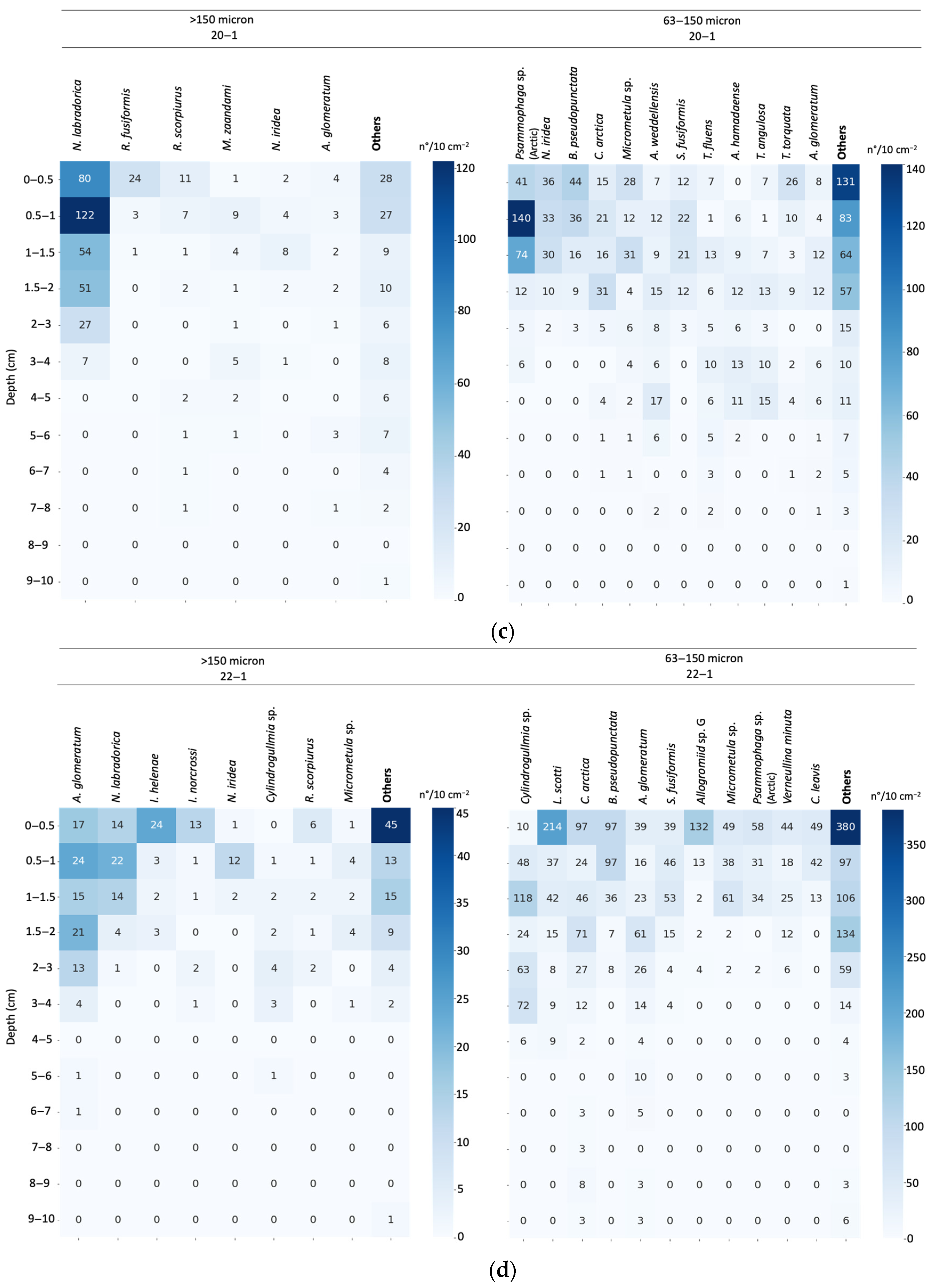
3.4. Assemblage Composition
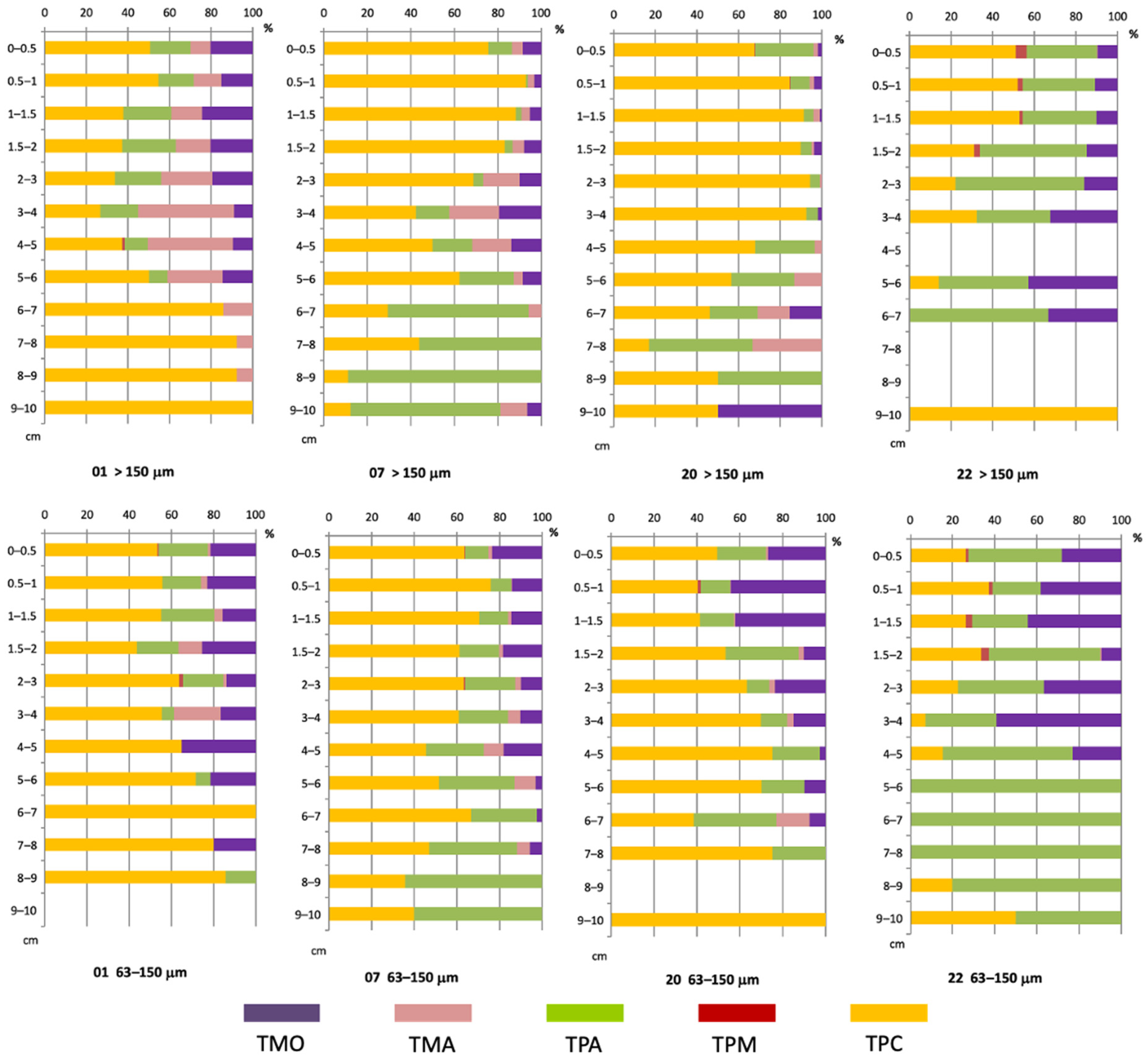
3.5. Vertical Distribution of Living Foraminiferal Assemblages
3.6. Relationship between Environmental Parameters and Foraminiferal Abundance and Biodiversity
4. Discussion
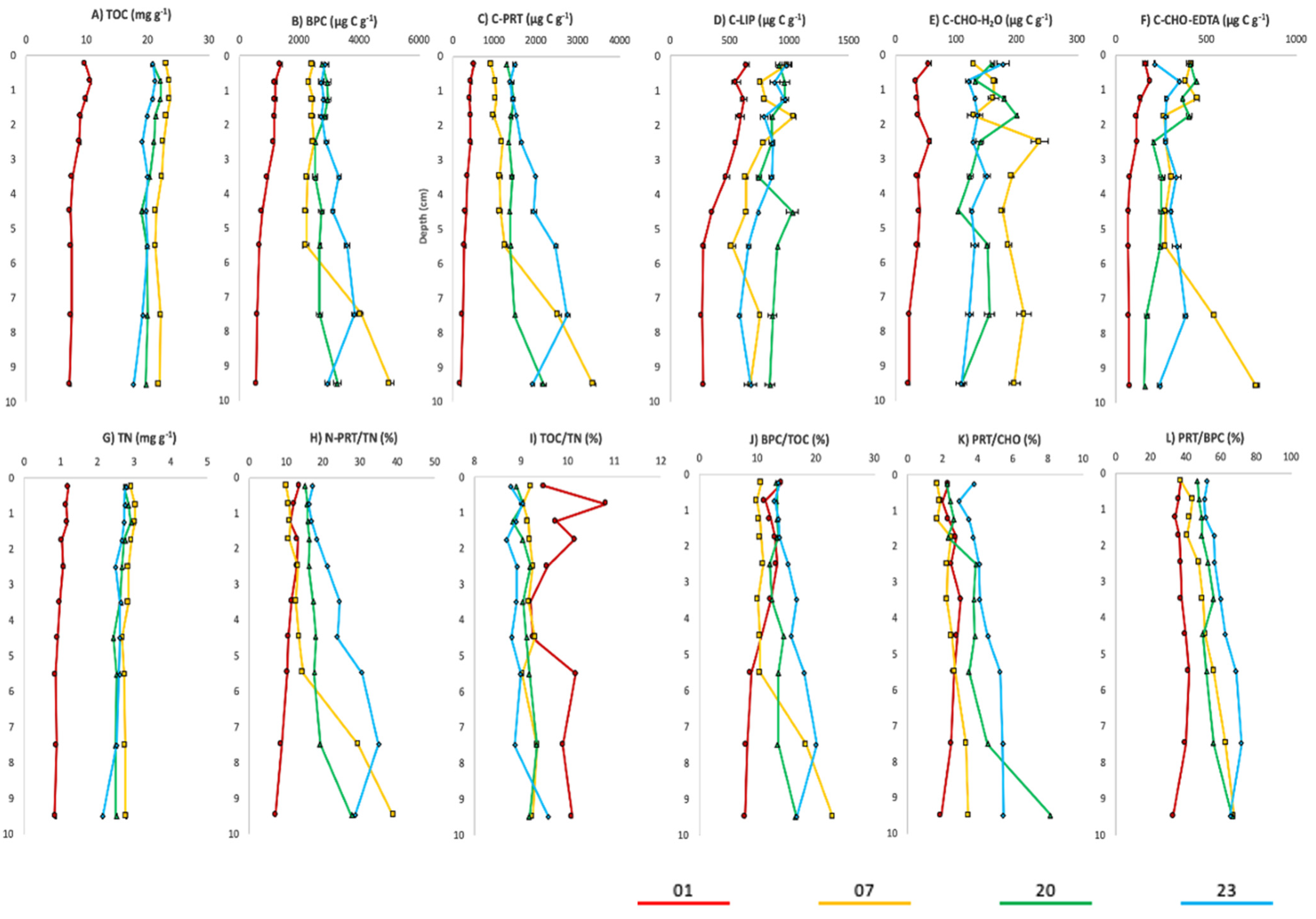
4.1. Patterns of Organic Matter Quantity and Nutritional Quality
4.2. Relationships between Organic Matter Pool and Living Foraminiferal Assemblage
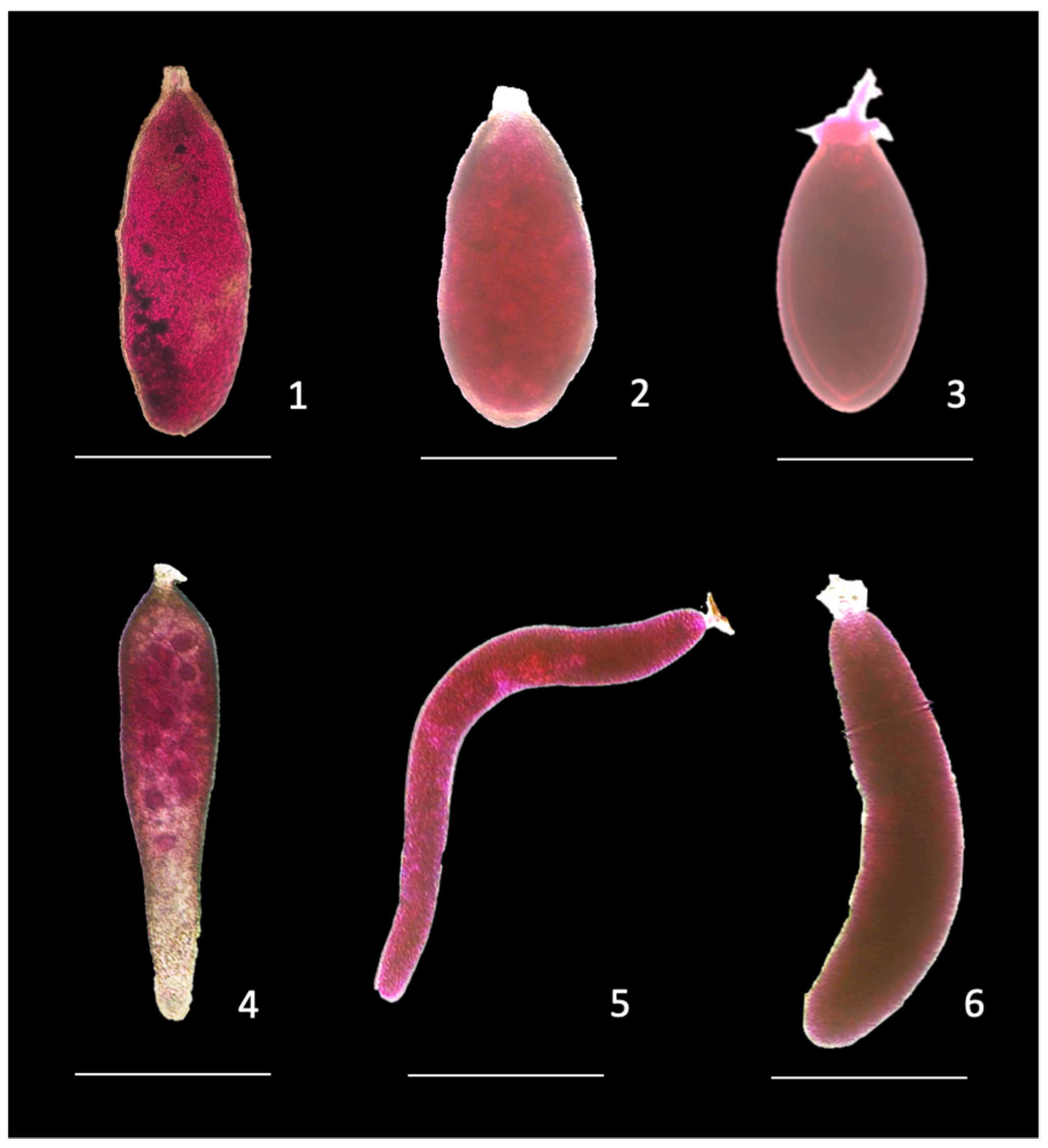
4.3. Relationships between High Organic Matter Accumulation in the Sediment, Foraminiferal Microhabitat and Actuopalentological Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furevik, T. Annual and Interannual Variability of Atlantic Water Temperatures in the Norwegian and Barents Seas: 1980–1996. Deep Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 383–404. [Google Scholar] [CrossRef]
- Dalpadado, P.; Ingvaldsen, R.; Hassel, A. Zooplankton Biomass Variation in Relation to Climatic Conditions in the Barents Sea. Polar Biol. 2003, 26, 233–241. [Google Scholar] [CrossRef]
- Sakshaug, E. Primary and Secondary Production in the Arctic Seas. In The Organic Carbon Cycle in the Arctic Ocean; Springer: Berlin/Heidelberg, Germany, 2004; pp. 57–81. [Google Scholar]
- Wassmann, P.; Slagstad, D.; Riser, C.W.; Reigstad, M. Modelling the Ecosystem Dynamics of the Barents Sea including the Marginal Ice Zone: II. Carbon Flux and Interannual Variability. J. Mar. Syst. 2006, 59, 1–24. [Google Scholar] [CrossRef]
- Barber, D.G.; Hop, H.; Mundy, C.J.; Else, B.; Dmitrenko, I.A.; Tremblay, J.-E.; Ehn, J.K.; Assmy, P.; Daase, M.; Candlish, L.M.; et al. Selected Physical, Biological and Biogeochemical Implications of a Rapidly Changing Arctic Marginal Ice Zone. Prog. Oceanogr. 2015, 139, 122–150. [Google Scholar] [CrossRef]
- Leu, E.; Mundy, C.J.; Assmy, P.; Campbell, K.; Gabrielsen, T.M.; Gosselin, M.; Juul-Pedersen, T.; Gradinger, R. Arctic Spring Awakening—Steering Principles behind the Phenology of Vernal Ice Algal Blooms. Prog. Oceanogr. 2015, 139, 151–170. [Google Scholar] [CrossRef]
- Michel, C.; Hamilton, J.; Hansen, E.; Barber, D.; Reigstad, M.; Iacozza, J.; Seuthe, L.; Niemi, A. Arctic Ocean Outflow Shelves in the Changing Arctic: A Review and Perspectives. Prog. Oceanogr. 2015, 139, 66–88. [Google Scholar] [CrossRef] [Green Version]
- Jeffries, M.O.; Richter-Menge, J.; Overland, J.E. Arctic Report Card 2015; NOAA: Washington, DC, USA, 2015. Available online: http://www.arctic.noaa.gov/Report-Card (accessed on 4 December 2022).
- Carmack, E.; Chapman, D.C. Wind-Driven Shelf/Basin Exchange on an Arctic Shelf: The Joint Roles of Ice Cover Extent and Shelf-Break Bathymetry. Geophys. Res. Lett. 2003, 30, 1778. [Google Scholar] [CrossRef]
- Tremblay, J.-É.; Simpson, K.; Martin, J.; Miller, L.; Gratton, Y.; Barber, D.; Price, N.M. Vertical Stability and the Annual Dynamics of Nutrients and Chlorophyll Fluorescence in the Coastal, Southeast Beaufort Sea. J. Geophys. Res. Ocean. 2008, 113, C07S90. [Google Scholar] [CrossRef]
- Stein, R.; Boucsein, B.; Meyer, H. Anoxia and High Primary Production in the Paleogene Central Arctic Ocean: First Detailed Records from Lomonosov Ridge. Geophys. Res. Lett. 2006, 33, L18606. [Google Scholar] [CrossRef]
- Cornwell, J.C.; Conley, D.J.; Owens, M.; Stevenson, J. A Sediment Chronology of the Eutrophication of Chesapeake Bay. Estuaries 1996, 19, 488–499. [Google Scholar] [CrossRef]
- Emeis, K.-C.; Struck, U.; Leipe, T.; Pollehne, F.; Kunzendorf, H.; Christiansen, C. Changes in the C, N, P Burial Rates in Some Baltic Sea Sediments over the Last 150 Years—Relevance to P Regeneration Rates and the Phosphorus Cycle. Mar. Geol. 2000, 167, 43–59. [Google Scholar] [CrossRef]
- Farías, L. Remineralization and Accumulation of Organic Carbon and Nitrogen in Marine Sediments of Eutrophic Bays: The Case of the Bay of Concepcion, Chile. Estuar. Coast. Shelf Sci. 2003, 57, 829–841. [Google Scholar] [CrossRef]
- Pusceddu, A.; Dell’Anno, A.; Fabiano, M.; Danovaro, R. Quantity and Bioavailability of Sediment Organic Matter as Signatures of Benthic Trophic Status. Mar. Ecol. Prog. Ser. 2009, 375, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Caridi, F.; Sabbatini, A.; Bensi, M.; Kovačević, V.; Lucchi, R.G.; Morigi, C.; Povea, P.; Negri, A. Benthic Foraminiferal Assemblages and Environmental Drivers along the Kveithola Trough (NW Barents Sea). J. Mar. Syst. 2021, 224, 103616. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006; p. 426. [Google Scholar]
- Louvari, M.A.; Drinia, H.; Kontakiotis, G.; Di Bella, L.; Antonarakou, A.; Anastasakis, G. Impact of Latest-Glacial to Holocene Sea-Level Oscillations on Central Aegean Shelf Ecosystems: A Benthic Foraminiferal Palaeoenvironmental Assessment of South Evoikos Gulf, Greece. J. Mar. Syst. 2019, 199, 103181. [Google Scholar] [CrossRef]
- Murray, J.W.; Alve, E. Benthic Foraminifera as Indicators of Environmental Change: Estuaries, Shelf and Upper Slope. In Environmental Quaternary Micropalaeontology; Haslett, S.R., Ed.; Hodder Arnold: London, UK, 2002; pp. 59–90. [Google Scholar]
- Nigam, R.; Saraswat, R.; Panchang, R. Application of Foraminifers in Ecotoxicology: Retrospect, Perspect and Prospect. Environ. Int. 2006, 32, 273–283. [Google Scholar] [CrossRef]
- Frontalini, F.; Coccioni, R. Benthic Foraminifera as Bioindicators of Pollution: A Review of Italian Research over the Last Three Decades. Rev. De Micropaléontologie 2011, 54, 115–127. [Google Scholar] [CrossRef]
- Fossile, E.; Sabbatini, A.; Spagnoli, F.; Caridi, F.; Dell’Anno, A.; De Marco, R.; Dinelli, E.; Droghini, E.; Tramontana, M.; Negri, A. Sensitivity of Foraminiferal-Based Indices to Evaluate the Ecological Quality Status of Marine Coastal Benthic Systems: A Case Study of the Gulf of Manfredonia (Southern Adriatic Sea). Mar. Pollut. Bull. 2021, 163, 111933. [Google Scholar] [CrossRef]
- Jorissen, F.J.; Barmawidjaja, D.M.; Puskaric, S.; Van der Zwaan, G.J. Vertical Distribution of Benthic Foraminifera in the Northern Adriatic Sea: The Relation with the Organic Flux. Mar. Micropaleontol. 1992, 19, 131–146. [Google Scholar] [CrossRef]
- Moodley, L.; Boschker, H.T.S.; Middelburg, J.J.; Pel, R.; Herman, P.M.J.; De Deckere, E.; Heip, C.H.R. Ecological Significance of Benthic Foraminifera: 13C Labelling Experiments. Mar. Ecol. Prog. Ser. 2000, 202, 289–295. [Google Scholar] [CrossRef]
- Barmawidjaja, D.M.; Van der Zwaan, G.J.; Jorissen, F.J.; Puskaric, S. 150 Years of Eutrophication in the Northern Adriatic Sea: Evidence from a Benthic Foraminiferal Record. Mar. Geol. 1995, 122, 367–384. [Google Scholar] [CrossRef]
- Loubere, P.; Fariduddin, M. Benthic Foraminifera and the Flux of Organic Carbon to the Seabed. Mod. Foraminifer. 1999, 181–199. [Google Scholar]
- Gooday, A.J. Deep-Sea Benthic Foraminiferal Species Which Exploit Phytodetritus: Characteristic Features and Controls on Distribution. Mar. Micropaleontol. 1993, 22, 187–205. [Google Scholar] [CrossRef]
- Altenbach, A.V.; Pflaumann, U.; Schiebel, R.; Thies, A.; Timm, S.; Trauth, M. Scaling Percentages and Distributional Patterns of Benthic Foraminifera with Flux Rates of Organic Carbon. J. Foraminifer. Res. 1999, 29, 173–185. [Google Scholar]
- Graf: Benthic-Pelagic Coupling: A Benthic View—Google Scholar. Available online: https://scholar.google.com/scholar_lookup?title=Benthic-pelagic%20coupling%3A%20a%20benthic%20view&publication_year=1992&author=G.%20Graf (accessed on 30 November 2022).
- Goedkoop, W.; Gullberg, K.R.; Johnson, R.K.; Ahlgren, I. Microbial Response of a Freshwater Benthic Community to a Simulated Diatom Sedimentation Event: Interactive Effects of Benthic Fauna. Microb. Ecol. 1997, 34, 131–143. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Barranguet, C.; Boschker, H.T.; Herman, P.M.; Moens, T.; Heip, C.H. The Fate of Intertidal Microphytobenthos Carbon: An in Situ 13C-Labeling Study. Limnol. Oceanogr. 2000, 45, 1224–1234. [Google Scholar] [CrossRef] [Green Version]
- Mackensen, A.; Douglas, R.G. Down-Core Distribution of Live and Dead Deep-Water Benthic Foraminifera in Box Cores from the Weddell Sea and the California Continental Borderland. Deep Sea Res. Part A Oceanogr. Res. Pap. 1989, 36, 879–900. [Google Scholar] [CrossRef]
- Gooday, A.J.; Bernhard, J.M.; Levin, L.A.; Suhr, S.B. Foraminifera in the Arabian Sea Oxygen Minimum Zone and Other Oxygen-deficient Settings: Taxonomic Composition, Diversity, and Relation to Metazoan Faunas. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 25–54. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Sen Gupta, B.K. Foraminifera of Oxygen-Depleted Environments. In Modern Foraminifera; Springer: Berlin/Heidelberg, Germany, 1999; pp. 201–216. [Google Scholar]
- Mallon, J.; Glock, N.; Schönfeld, J. The Response of Benthic Foraminifera to Low-Oxygen Conditions of the Peruvian Oxygen Minimum Zone. In Anoxia; Springer: Berlin/Heidelberg, Germany, 2012; pp. 305–321. [Google Scholar]
- Sergeeva, N.G.; Gooday, A.J.; Mazlumyan, S.A.; Kolesnikova, E.A.; Lichtschlag, A.; Kosheleva, T.N.; Anikeeva, O.V. Meiobenthos of the Oxic/Anoxic Interface in the Southwestern Region of the Black Sea: Abundance and Taxonomic Composition. In Anoxia; Springer: Berlin/Heidelberg, Germany, 2012; pp. 369–401. [Google Scholar]
- Corliss, B.H. Microhabitats of Benthic Foraminifera within Deep-Sea Sediments. Nature 1985, 314, 435–438. [Google Scholar] [CrossRef]
- Corliss, B.H.; Emerson, S. Distribution of Rose Bengal Stained Deep-Sea Benthic Foraminifera from the Nova Scotian Continental Margin and Gulf of Maine. Deep Sea Res. Part A. Oceanogr. Res. Pap. 1990, 37, 381–400. [Google Scholar] [CrossRef]
- Risgaard-Petersen, N.; Langezaal, A.M.; Ingvardsen, S.; Schmid, M.C.; Jetten, M.S.; Op den Camp, H.J.; Derksen, J.W.; Pina-Ochoa, E.; Eriksson, S.P.; Peter Nielsen, L. Evidence for Complete Denitrification in a Benthic Foraminifer. Nature 2006, 443, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Pina-Ochoa, E.; Alvarez-Cobelas, M. Seasonal Nitrogen Dynamics in a Seepage Lake Receiving High Nitrogen Loads. Mar. Freshw. Res. 2009, 60, 435–445. [Google Scholar] [CrossRef]
- Koho, K.A.; Piña-Ochoa, E. Benthic Foraminifera: Inhabitants of Low-Oxygen Environments. In Anoxia; Cellular Origin, Life in Extreme Habitats and Astrobiology; Altenbach, A.V., Bernhard, J.M., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 21, pp. 249–285. ISBN 978-94-007-1895-1. [Google Scholar]
- Korsun, S.A.; Pogodina, I.A.; Tarasov, G.A.; Matishov, G.G. Foraminifera of the Barents Sea: Hydrobiology and Quaternary Paleoecology. Kola Sci. Cent. Publ. Russ. Acad. Sci. Apatity 1994, 140. Available online: https://scholar.google.com/scholar?hl=it&as_sdt=0,5&cluster=1408039786455229986 (accessed on 4 December 2022).
- Ivanova, E.V.; Ovsepyan, E.A.; Risebrobakken, B.; Vetrov, A.A. Downcore Distribution of Living Calcareous Foraminifera and Stable Isotopes in the Western Barents Sea. J. Foraminifer. Res. 2008, 38, 337–356. [Google Scholar] [CrossRef]
- Saher, M.H.; Kristensen, D.K.; Hald, M.; Korsun, S.; Jørgensen, L.L. Benthic Foraminifera Assemblages in the Central Barents Sea: An Evaluation of Combining Live and Total Fauna Studies in Tracking Environmental Change. Nor. J. Geol. /Nor. Geol. Foren. 2009, 89, 149–161. [Google Scholar]
- Dijkstra, N. Benthic Foraminifera as Indicators of Natural Variability and Anthropogenic Impact; University of Tromsø: Tromsø, Norway, 2013. [Google Scholar]
- Carbonara, K.; Mezgec, K.; Varagona, G.; Musco, M.E.; Lucchi, R.G.; Villa, G.; Morigi, C.; Melis, R.; Caffau, M. Palaeoclimatic Changes in Kveithola, Svalbard, during the Late Pleistocene Deglaciation and Holocene: Evidences from Microfossil and Sedimentary Records. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 463, 136–149. [Google Scholar] [CrossRef]
- Ivanova, E.; Murdmaa, I.; de Vernal, A.; Risebrobakken, B.; Peyve, A.; Brice, C.; Seitkalieva, E.; Pisarev, S. Postglacial Paleoceanography and Paleoenvironments in the Northwestern Barents Sea. Quat. Res. 2019, 92, 430–449. [Google Scholar] [CrossRef]
- Gamboa-Sojo, V.M.; Husum, K.; Caridi, F.; Lucchi, R.G.; Bensi, M.; Kovačević, V.; Sabbatini, A.; Langone, L.; Dominiczak, A.T.; Povea, P.; et al. Living and Dead Foraminiferal Assemblages of the Last Decades from Kveithola Trough: Taphonomic Processes and Ecological Highlights. Mar. Micropaleontol. 2021, 166, 102014. [Google Scholar] [CrossRef]
- Steinsund, P.I.; Hald, M. Recent Calcium Carbonate Dissolution in the Barents Sea: Paleoceanographic Applications. Mar. Geol. 1994, 117, 303–316. [Google Scholar] [CrossRef]
- Hald, M.; Steinsund, P.I. Benthic Foraminifera and Carbonate Dissolution in the Surface Sediments of the Barents and Kara Seas. Ber. Zur Polarforsch. 1996, 212, 285–307. [Google Scholar]
- Basov, V.A.; Slobodin, V.Y. Complexes of Recent and Late Cenozoic Foraminifera of WeStern Soviet Arctic. The Antropogene Period of the Arctic and Subarctic. Tr. Nauchn. Issled. Inst. Geol. Arkt. Gosud. Geol. Kom. SSSR 1965, 143, 190–210. [Google Scholar]
- Digas, L.A. Distribution of Foraminifers in Surface Sediments of the Barents Sea and Adjacent Parts of the Norwegian-Greenland Basin. PhD Thesis, Saratov State University, Saratov, Russia, 1969; 27p. (In Russian). [Google Scholar]
- Digas, L.A. A Zoogeographic Zonation of the Barents Sea Based on Foraminifera. Geol. Issues South Ural. Volga [Vopr. Geol. Yuzhnogo Ural. I Povolzhja]. Saratov Saratov State Univ. Publ 1970, 127–142. Available online: https://scholar.google.com/scholar?hl=it&as_sdt=0%2C5&q=Digas%2C+1970&btnG= (accessed on 4 December 2022).
- Østby, K.L.; Nagy, J. Foraminiferal Distribution in the Western Barents Sea, Recent and Quaternary. Polar Res. 1982, 1982, 53–87. [Google Scholar] [CrossRef]
- Polyak, L.V. Foraminifers of Bottom Sediments of the Barents and Kara Sea and Their Stratigraphic Significance; Abstract of Kandidat Disseration; LGI: Leningrad, Russia, 1985. [Google Scholar]
- Hald, M.; Steinsund, P.I. Distribution of Surface Sediment Benthic Foraminifera in the Southwestern Barents Sea. J. Foraminifer. Res. 1992, 22, 347–362. [Google Scholar] [CrossRef]
- Kucharska, M.; Kujawa, A.; Pawłowska, J.; Łącka, M.; Szymańska, N.; Lønne, O.J.; Zajączkowski, M. Seasonal Changes in Foraminiferal Assemblages along Environmental Gradients in Adventfjorden (West Spitsbergen). Polar Biol. 2019, 42, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Fossile, E.; Nardelli, M.P.; Jouini, A.; Lansard, B.; Pusceddu, A.; Moccia, D.; Michel, E.; Péron, O.; Howa, H.; Mojtahid, M. Benthic Foraminifera as Tracers of Brine Production in the Storfjorden “Sea Ice Factory”. Biogeosciences 2020, 17, 1933–1953. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini, A.; Morigi, C.; Negri, A.; Gooday, A.J. Distribution and Biodiversity of Stained Monothalamous Foraminifera from Tempelfjord, Svalbard. J. Foraminifer. Res. 2007, 37, 93–106. [Google Scholar] [CrossRef]
- Fichez, R. Composition and Fate of Organic-Matter in Submarine Cave Sediments-Implications for the Biogeochemical Cycle of Organic-Carbon. Oceanol. Acta 1991, 14, 369–377. [Google Scholar]
- Rebesco, M.; Liu, Y.; Camerlenghi, A.; Winsborrow, M.; Laberg, J.S.; Caburlotto, A.; Diviacco, P.; Accettella, D.; Sauli, C.; Wardell, N.; et al. Deglaciation of the Western Margin of the Barents Sea Ice Sheet—A Swath Bathymetric and Sub-Bottom Seismic Study from the Kveithola Trough. Mar. Geol. 2011, 279, 141–147. [Google Scholar] [CrossRef]
- Rüther, D.C.; Bjarnadóttir, L.R.; Junttila, J.; Husum, K.; Rasmussen, T.L.; Lucchi, R.G.; Andreassen, K. Pattern and Timing of the Northwestern Barents Sea Ice Sheet Deglaciation and Indications of Episodic Holocene Deposition: Barents Sea Ice Sheet Deglaciation and Episodic Holocene Deposition. Boreas 2012, 41, 494–512. [Google Scholar] [CrossRef]
- Bjarnadóttir, L.R.; Rüther, D.C.; Winsborrow, M.C.M.; Andreassen, K. Grounding-Line Dynamics during the Last Deglaciation of Kveithola, W Barents Sea, as Revealed by Seabed Geomorphology and Shallow Seismic Stratigraphy: Grounding Line Dynamics, W Barents Sea. Boreas 2013, 42, 84–107. [Google Scholar] [CrossRef]
- Ottesen, D.; Dowdeswell, J.A. Assemblages of Submarine Landforms Produced by Tidewater Glaciers in Svalbard. J. Geophys. Res. Earth Surf. 2006, 111, F01016. [Google Scholar] [CrossRef]
- Dowdeswell, J.A.; Ottesen, D.; Evans, J.; Cofaigh, C.; Anderson, J.B. Submarine Glacial Landforms and Rates of Ice-Stream Collapse. Geology 2008, 36, 819–822. [Google Scholar] [CrossRef]
- Rebesco, M.; Özmaral, A.; Urgeles, R.; Accettella, D.; Lucchi, R.G.; Rüther, D.; Winsborrow, M.; Llopart, J.; Caburlotto, A.; Lantzsch, H.; et al. Evolution of a High-Latitude Sediment Drift inside a Glacially-Carved Trough Based on High-Resolution Seismic Stratigraphy (Kveithola, NW Barents Sea). Quat. Sci. Rev. 2016, 147, 178–193. [Google Scholar] [CrossRef] [Green Version]
- Lucchi, R.G.; Kovacevic, V.; De Vittor, C.; Bazzaro, M.; Bensi, M.; De Ponte, D.; Laterza, R.; Musco, M.E.; Relitti, F.; Rui, L. Burster: Bottom Currents in a Stagnant Environment; 2016. Available online: https://scholar.google.com/scholar?hl=it&as_sdt=0%2C5&q=+Lucchi%2C+R.G.%3B+Kovacevic%2C+V.%3B+De+Vittor%2C+C.%3B+Bazzaro%2C+M.%3B+Bensi%2C+M.%3B+De+Ponte%2C+D.%3B+Laterza%2C+R.%3B+Musco%2C+M.E.%3B+Relitti%2C+F.%3B+Rui%2C+L.+Bur-ster%3A+Bottom+Currents+in+a+Stagnant+Environment%3B+2016&btnG= (accessed on 4 December 2022).
- Hanebuth, T.J.; Lantzsch, H.; Bergenthal, M.; Caburlotto, A.; Dippold, S.; Düßmann, R.; Freudenthal, T.; Hörner, T.; Kaszemeik, K.; Klar, S. CORIBAR-Ice Dynamics and Meltwater Deposits: Coring in the Kveithola Trough, NW Barents Sea. Cruise MSM30. 16 July–15 August 2013, Tromsø (Norway)-Tromsø (Norway). Ber. MARUM-Zent. Für Mar. Umweltwiss. Fachbereich Geowiss. Univ. Brem. 2013, 299. Available online: https://publications.marum.de/2347/ (accessed on 4 December 2022).
- Loeng, H. Features of the Physical Oceanographic Conditions of the Barents Sea. Polar Res. 1991, 10, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Piepenburg, D.; Schmid, M.K. Brittle Star Fauna (Echinodermata: Ophiuroidea) of the Arctic Northwestern Barents Sea: Composition, Abundance, Biomass and Spatial Distribution. Polar Biol. 1996, 16, 383–392. [Google Scholar] [CrossRef]
- Piepenburg, D.; Schmid, M.K. A Photographic Survey of the Epibenthic Megafauna of the Arctic Laptev Sea Shelf: Distribution, Abundance, and Estimates of Biomass and Organic Carbon Demand. Mar. Ecol. Prog. Ser. 1997, 147, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Grebmeier, J.M.; Cooper, L.W.; Feder, H.M.; Sirenko, B.I. Ecosystem Dynamics of the Pacific-Influenced Northern Bering and Chukchi Seas in the Amerasian Arctic. Prog. Oceanogr. 2006, 71, 331–361. [Google Scholar] [CrossRef]
- Pella, E.; Colombo, B. Study of Carbon, Hydrogen and Nitrogen Determination by Combustion-Gas Chromatography. Microchim. Acta 1973, 61, 697–719. [Google Scholar] [CrossRef]
- Nieuwenhuize, J.; Maas, Y.E.; Middelburg, J.J. Rapid Analysis of Organic Carbon and Nitrogen in Particulate Materials. Mar. Chem. 1994, 45, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Blasutto, O.; Cibic, T.; Vittor, C.D.; Umani, S.F. Microphytobenthic Primary Production and Sedimentary Carbohydrates along Salinity Gradients in the Lagoons of Grado and Marano (Northern Adriatic Sea). Hydrobiologia 2005, 550, 47–55. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Gerchakov, S.M.; Hatcher, P.G. Improved Technique for Analysis of Carbohydrates in Sediments 1. Limnol. Oceanogr. 1972, 17, 938–943. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R.; Fraschetti, S. A Three-Year Time Series of Elemental and Biochemical Composition of Organic Matter in Subtidal Sandy Sediments of the Ligurian Sea (Northwestern Mediterranean). Cont. Shelf Res. 1995, 15, 1453–1469. [Google Scholar] [CrossRef]
- Hartree, E.F. Determination of Protein: A Modification of the Lowry Method That Gives a Linear Photometric Response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Rice, D.L. The Detritus Nitrogen Problem: New Observations and Perspectives from Organic Geochemistry. Mar. Ecol. Prog. Ser. 1982, 9, 153–162. [Google Scholar] [CrossRef]
- Fabiano, M.; Pusceddu, A.; Dell’Anno, A.; Armeni, M.; Vanucci, S.; Lampitt, R.S.; Wolff, G.A.; Danovaro, R. Fluxes of Phytopigments and Labile Organic Matter to the Deep Ocean in the NE Atlantic Ocean. Prog. Oceanogr. 2001, 50, 89–104. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Marsh, J.B.; Weinstein, D.B. Simple Charring Method for Determination of Lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef]
- Danovaro, R.; Fabiano, M.; Della Croce, N. Labile Organic Matter and Microbial Biomasses in Deep-Sea Sediments (Eastern Mediterranean Sea). Deep Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 953–965. [Google Scholar] [CrossRef]
- Fabiano, M.; Chiantore, M.; Povero, P.; Cattaneo-Vietti, R.; Pusceddu, A.; Misic, C.; Albertelli, G. Short-Term Variations in Particulate Matter Flux in Terra Nova Bay, Ross Sea. Antarct. Sci. 1997, 9, 143–149. [Google Scholar] [CrossRef]
- Tselepides, A.; Polychronaki, T.; Marrale, D.; Akoumianaki, I.; Dell’Anno, A.; Pusceddu, A.; Danovaro, R. Organic Matter Composition of the Continental Shelf and Bathyal Sediments of the Cretan Sea (NE Mediterranean). Prog. Oceanogr. 2000, 46, 311–344. [Google Scholar] [CrossRef]
- Cividanes, S.; Incera, M.; López, J. Temporal Variability in the Biochemical Composition of Sedimentary Organic Matter in an Intertidal Flat of the Galician Coast (NW Spain). Oceanol. Acta 2002, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, A.; Mei, M.L.; Pusceddu, A.; Danovaro, R. Assessing the Trophic State and Eutrophication of Coastal Marine Systems: A New Approach Based on the Biochemical Composition of Sediment Organic Matter. Mar. Pollut. Bull. 2002, 44, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, A.; Bianchelli, S.; Canals, M.; Sanchez-Vidal, A.; De Madron, X.D.; Heussner, S.; Lykousis, V.; de Stigter, H.; Trincardi, F.; Danovaro, R. Organic Matter in Sediments of Canyons and Open Slopes of the Portuguese, Catalan, Southern Adriatic and Cretan Sea Margins. Deep Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 441–457. [Google Scholar] [CrossRef]
- Nardelli, M.P. Response of Benthic Foraminifera to Pollution through Experimental and In Situ Studies: Advances in Biological Aspects and Tools for Future Application in Biomonitoring; Università Politecnica delle Marche: Ancona, Italy, 2012. [Google Scholar]
- Sabbatini, A.; Bonatto, S.; Gooday, A.J.; Morigi, C.; Pancotti, I.; Pucci, F.; Negri, A. Modern Benthic Foraminifers at Northern Shallow Sites of Adriatic Sea and Soft-Walled, Monothalamous Taxa: A Brief Overview. Micropaleontology 2010, 56, 359–376. [Google Scholar] [CrossRef]
- Sabbatini, A.; Bonatto, S.; Bianchelli, S.; Pusceddu, A.; Danovaro, R.; Negri, A. Foraminiferal Assemblages and Trophic State in Coastal Sediments of the Adriatic Sea. J. Mar. Syst. 2012, 105–108, 163–174. [Google Scholar] [CrossRef]
- Murray, J.W.; Bowser, S.S. Mortality, Protoplasm Decay Rate, and Reliability of Staining Techniques to Recognize ‘Living’Foraminifera: A Review. J. Foraminifer. Res. 2000, 30, 66–70. [Google Scholar] [CrossRef]
- Bernhard, J.M. Distinguishing Live from Dead Foraminifera: Methods Review and Proper Applications. Micropaleontology 2000, 46, 38–46. [Google Scholar]
- Loeblich, A.R.; Tappan, H.N. Studies of Arctic Foraminifera. Smithson. Misc. Collect. 1953, 121, 150. [Google Scholar]
- Seidenkrantz, M.-S. Cassidulina Teretis Tappan and Cassidulina Neoteretis New Species (Foraminifera): Stratigraphic Markers for Deep Sea and Outer Shelf Areas. J. Micropalaeontol. 1995, 14, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Wollenburg, J.E.; Mackensen, A. Living Benthic Foraminifers from the Central Arctic Ocean: Faunal Composition, Standing Stock and Diversity. Mar. Micropaleontol. 1998, 34, 153–185. [Google Scholar] [CrossRef]
- Wollenburg, J.E.; Mackensen, A. On the Vertical Distribution of Living (Rose Bengal Stained) Benthic Foraminifers in the Arctic Ocean. J. Foraminifer. Res. 1998, 28, 268–285. [Google Scholar] [CrossRef]
- Wollenburg, J.E.; Mackensen, A. The Ecology and Distribution of Benthic Foraminifera at the Håkon Mosby Mud Volcano (SW Barents Sea Slope). Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 1336–1370. [Google Scholar] [CrossRef]
- Majewski, W.; Zajaczkowski, M. Benthic Foraminifera in Adventfjorden, Svalbard: Last 50 Years of Local Hydrographic Changes. J. Foraminifer. Res. 2007, 37, 107–124. [Google Scholar] [CrossRef]
- Gooday, A.J.; Bowser, S.S.; Cedhagen, T.; Cornelius, N.; Hald, M.; Korsun, S.; Pawlowski, J. Monothalamous Foraminiferans and Gromiids (Protista) from Western Svalbard: A Preliminary Survey. Mar. Biol. Res. 2005, 1, 290–312. [Google Scholar] [CrossRef]
- Gooday, A.J.; Malzone, M.G.; Bett, B.J.; Lamont, P.A. Decadal-Scale Changes in Shallow-Infaunal Foraminiferal Assemblages at the Porcupine Abyssal Plain, NE Atlantic. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1362–1382. [Google Scholar] [CrossRef]
- Majewski, W.; Pawłowski, J.; Zajączkowski, M. Monothalamous Foraminifera from West Spitsbergen Fjords, Svalbard: A Brief Overview. Pol. Polar Res. 2005, 26, 269–285. [Google Scholar]
- Sabbatini, A.; Nardelli, M.P.; Morigi, C.; Negri, A. Contribution of Soft-Shelled Monothalamous Taxa to Foraminiferal Assemblages in the Adriatic Sea. Acta Protozool. 2013, 52. [Google Scholar]
- Pielou, E.C. Ecological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 1975. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Jorissen, F.J.; de Stigter, H.C.; Widmark, J.G.V. A Conceptual Model Explaining Benthic Foraminiferal Microhabitats. Mar. Micropaleontol. 1995, 26, 3–15. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A Further Biodiversity Index Applicable to Species Lists: Variation in Taxonomic Distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, R.K. Permanova + for Primer: Guide to Software and Statisticl Methods; Primer-E Limited: Plymouth, UK, 2008; p. 218. [Google Scholar]
- Gribble, G.W. The Diversity of Naturally Produced Organohalogens. Chemosphere 2003, 52, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.; Croudace, I.W.; Rothwell, R.G. A Geochemical Application of the ITRAX Scanner to a Sediment Core Containing Eastern Mediterranean Sapropel Units. Geol. Soc. Lond. Spec. Publ. 2006, 267, 65–77. [Google Scholar] [CrossRef]
- Passier, H.F.; Middelburg, J.J.; de Lange, G.J.; Böttcher, M.E. Modes of Sapropel Formation in the Eastern Mediterranean: Some Constraints Based on Pyrite Properties. Mar. Geol. 1999, 153, 199–219. [Google Scholar] [CrossRef]
- Lucchi, R.G.; Rebesco, M.; Camerlenghi, A.; Busetti, M.; Tomadin, L.; Villa, G.; Persico, D.; Morigi, C.; Bonci, M.C.; Giorgetti, G. Mid-Late Pleistocene Glacimarine Sedimentary Processes of a High-Latitude, Deep-Sea Sediment Drift (Antarctic Peninsula Pacific Margin). Mar. Geol. 2002, 189, 343–370. [Google Scholar] [CrossRef]
- Lucchi, R.G.; Camerlenghi, A.; Rebesco, M.; Colmenero-Hidalgo, E.; Sierro, F.J.; Sagnotti, L.; Urgeles, R.; Melis, R.; Morigi, C.; Bárcena, M.-A.; et al. Postglacial Sedimentary Processes on the Storfjorden and Kveithola Trough Mouth Fans: Significance of Extreme Glacimarine Sedimentation. Glob. Planet. Chang. 2013, 111, 309–326. [Google Scholar] [CrossRef]
- Caricchi, C.; Lucchi, R.G.; Sagnotti, L.; Macrì, P.; Morigi, C.; Melis, R.; Caffau, M.; Rebesco, M.; Hanebuth, T.J. Paleomagnetism and Rock Magnetism from Sediments along a Continental Shelf-to-Slope Transect in the NW Barents Sea: Implications for Geomagnetic and Depositional Changes during the Past 15 Thousand Years. Glob. Planet. Chang. 2018, 160, 10–27. [Google Scholar] [CrossRef]
- Renaud, P.E.; Morata, N.; Carroll, M.L.; Denisenko, S.G.; Reigstad, M. Pelagic–Benthic Coupling in the Western Barents Sea: Processes and Time Scales. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2372–2380. [Google Scholar] [CrossRef]
- Wassmann, P.; Reigstad, M. Future Arctic Ocean Seasonal Ice Zones and Implications for Pelagic-Benthic Coupling. Oceanography 2011, 24, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Søreide, J.E.; Carroll, M.L.; Hop, H.; Ambrose, W.G., Jr.; Hegseth, E.N.; Falk-Petersen, S. Sympagic-Pelagic-Benthic Coupling in Arctic and Atlantic Waters around Svalbard Revealed by Stable Isotopic and Fatty Acid Tracers. Mar. Biol. Res. 2013, 9, 831–850. [Google Scholar] [CrossRef]
- Wlodarska-Kowalczuk, M.; Pearson, T.H.; Kendall, M.A. Benthic Response to Chronic Natural Physical Disturbance by Glacial Sedimentation in an Arctic Fjord. Mar. Ecol. Prog. Ser. 2005, 303, 31–41. [Google Scholar] [CrossRef]
- Kędra, M.; Renaud, P.E.; Andrade, H.; Goszczko, I.; Ambrose, W.G. Benthic Community Structure, Diversity, and Productivity in the Shallow Barents Sea Bank (Svalbard Bank). Mar. Biol. 2013, 160, 805–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ślubowska-Woldengen, M.; Koç, N.; Rasmussen, T.L.; Klitgaard-Kristensen, D.; Hald, M.; Jennings, A.E. Time-Slice Reconstructions of Ocean Circulation Changes on the Continental Shelf in the Nordic and Barents Seas during the Last 16,000 Cal Yr BP. Quat. Sci. Rev. 2008, 27, 1476–1492. [Google Scholar] [CrossRef]
- Grémare, A.; Medernach, L.; DeBovee, F.; Amouroux, J.M.; Vétion, G.; Albert, P. Relationships between Sedimentary Organics and Benthic Meiofauna on the Continental Shelf and the Upper Slope of the Gulf of Lions (NW Mediterranean). Mar. Ecol. Prog. Ser. 2002, 234, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Schubert, C.J.; Calvert, S.E. Nitrogen and Carbon Isotopic Composition of Marine and Terrestrial Organic Matter in Arctic Ocean Sediments:: Implications for Nutrient Utilization and Organic Matter Composition. Deep Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 789–810. [Google Scholar] [CrossRef]
- Goñi, M.A.; Teixeira, M.J.; Perkey, D.W. Sources and Distribution of Organic Matter in a River-Dominated Estuary (Winyah Bay, SC, USA). Estuar. Coast. Shelf Sci. 2003, 57, 1023–1048. [Google Scholar] [CrossRef]
- Meyers, P.A. Preservation of Elemental and Isotopic Source Identification of Sedimentary Organic Matter. Chem. Geol. 1994, 114, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Knies, J.; Martinez, P. Organic Matter Sedimentation in the Western Barents Sea Region: Terrestrial and Marine Contribution Based on Isotopic Composition and Organic Nitrogen Content. Nor. J. Geol./Nor. Geol. Foren. 2009, 89, 79–89. [Google Scholar]
- Pusceddu, A.; Bianchelli, S.; Gambi, C.; Danovaro, R. Assessment of Benthic Trophic Status of Marine Coastal Ecosystems: Significance of Meiofaunal Rare Taxa. Estuar. Coast. Shelf Sci. 2011, 93, 420–430. [Google Scholar] [CrossRef]
- Bianchelli, S.; Gambi, C.; Pusceddu, A.; Danovaro, R. Trophic Conditions and Meiofaunal Assemblages in the Bari Canyon and the Adjacent Open Slope (Adriatic Sea). Chem. Ecol. 2008, 24, 101–109. [Google Scholar] [CrossRef]
- Pusceddu, A.; Dell’Anno, A.; Danovaro, R.; Manini, E.; Sara, G.; Fabiano, M. Enzymatically Hydrolyzable Protein and Carbohydrate Sedimentary Pools as Indicators of the Trophic State of Detritus Sink Systems: A Case Study in a Mediterranean Coastal Lagoon. Estuaries 2003, 26, 641–650. [Google Scholar] [CrossRef]
- Lantzsch, H.; Hanebuth, T.J.J.; Horry, J.; Grave, M.; Rebesco, M.; Schwenk, T. Deglacial to Holocene History of Ice-Sheet Retreat and Bottom Current Strength on the Western Barents Sea Shelf. Quat. Sci. Rev. 2017, 173, 40–57. [Google Scholar] [CrossRef]
- Caridi, F.; Sabbatini, A.; Morigi, C.; Dell’Anno, A.; Negri, A.; Lucchi, R.G. Patterns and Environmental Drivers of Diversity and Community Composition of Macrofauna in the Kveithola Trough (NW Barents Sea). J. Sea Res. 2019, 153, 101780. [Google Scholar] [CrossRef]
- Morigi, C.; Jorissen, F.J.; Gervais, A.; Guichard, S.; Borsetti, A.M. Benthic Foraminiferal Faunas in Surface Sediments off NW Africa: Relationship with Organic Flux to the Ocean Floor. J. Foraminifer. Res. 2001, 31, 350–368. [Google Scholar] [CrossRef]
- Fontanier, C.; Jorissen, F.J.; Chaillou, G.; Anschutz, P.; Grémare, A.; Griveaud, C. Live Foraminiferal Faunas from a 2800 m Deep Lower Canyon Station from the Bay of Biscay: Faunal Response to Focusing of Refractory Organic Matter. Deep Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 1189–1227. [Google Scholar] [CrossRef]
- Gooday, A.J. A Response by Benthic Foraminifera to the Deposition of Phytodetritus in the Deep Sea. Nature 1988, 332, 70–73. [Google Scholar] [CrossRef]
- Gooday, A.J.; Turley, C.M. Responses by Benthic Organisms to Inputs of Organic Material to the Ocean Floor: A Review. Philos. Trans. R. Soc. London. Ser. A Math. Phys. Sci. 1990, 331, 119–138. [Google Scholar]
- Gooday, A.J.; Rathburn, A.E. Temporal Variability in Living Deep-Sea Benthic Foraminifera: A Review. Earth-Sci. Rev. 1999, 46, 187–212. [Google Scholar] [CrossRef]
- Gooday, A.J. Biological Responses to Seasonally Varying Fluxes of Organic Matter to the Ocean Floor: A Review. J. Oceanogr. 2002, 58, 305–332. [Google Scholar] [CrossRef]
- Kitazato, H.; Shirayama, Y.; Nakatsuka, T.; Fujiwara, S.; Shimanaga, M.; Kato, Y.; Okada, Y.; Kanda, J.; Yamaoka, A.; Masuzawa, T. Seasonal Phytodetritus Deposition and Responses of Bathyal Benthic Foraminiferal Populations in Sagami Bay, Japan: Preliminary Results from “Project Sagami 1996–1999”. Mar. Micropaleontol. 2000, 40, 135–149. [Google Scholar] [CrossRef]
- Licari, L.N. Communities and Microhabitats of Living Benthic Foraminifera from the Tropical East Atlantic: Impact of Different Productivity Regimes. J. Foraminifer. Res. 2003, 33, 10–31. [Google Scholar] [CrossRef]
- Saher, M.; Kristensen, D.K.; Hald, M.; Pavlova, O.; Jørgensen, L.L. Changes in Distribution of Calcareous Benthic Foraminifera in the Central Barents Sea between the Periods 1965–1992 and 2005–2006. Glob. Planet. Chang. 2012, 98–99, 81–96. [Google Scholar] [CrossRef]
- Mackensen, A.; Sejrup, H.P.; Jansen, E. The Distribution of Living Benthic Foraminifera on the Continental Slope and Rise off Southwest Norway. Mar. Micropaleontol. 1985, 9, 275–306. [Google Scholar] [CrossRef]
- Corliss, B.H.; Chen, C. Morphotype Patterns of Norwegian Sea Deep-Sea Benthic Foraminifera and Ecological Implications. Geology 1988, 16, 716–719. [Google Scholar] [CrossRef]
- Linke, P.; Lutze, G.F. Microhabitat Preferences of Benthic Foraminifera—A Static Concept or a Dynamic Adaptation to Optimize Food Acquisition? Mar. Micropaleontol. 1993, 20, 215–234. [Google Scholar] [CrossRef] [Green Version]
- Yak, L.P.; Solheim, A. Late-and Postglacial Environments in the Northern Barents Sea West of Franz Josef Land. Polar Res. 1994, 13, 197–207. [Google Scholar] [CrossRef]
- Polyak, L.; Korsun, S.; Febo, L.A.; Stanovoy, V.; Khusid, T.; Hald, M.; Paulsen, B.E.; Lubinski, D.J. Benthic Foraminiferal Assemblages from the Southern Kara Sea, a River-Influenced Arctic Marine Environment. J. Foraminifer. Res. 2002, 32, 252–273. [Google Scholar] [CrossRef] [Green Version]
- Wollenburg, J.E.; Kuhnt, W. The Response of Benthic Foraminifers to Carbon Flux and Primary Production in the Arctic Ocean. Mar. Micropaleontol. 2000, 40, 189–231. [Google Scholar] [CrossRef]
- Rytter, F. Modern Distribution of Benthic Foraminifera on the North Icelandic Shelf and Slope. J. Foraminifer. Res. 2002, 32, 217–244. [Google Scholar] [CrossRef]
- Wollenburg, J.E.; Knies, J.; Mackensen, A. High-Resolution Paleoproductivity Fluctuations during the Past 24 Kyr as Indicated by Benthic Foraminifera in the Marginal Arctic Ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 204, 209–238. [Google Scholar] [CrossRef]
- Wollenburg, J.E.; Kuhnt, W.; Mackensen, A. Changes in Arctic Ocean Paleoproductivity and Hydrography during the Last 145 Kyr: The Benthic Foraminiferal Record. Paleoceanography 2001, 16, 65–77. [Google Scholar] [CrossRef]
- Husum, K.; Hald, M. Early Holocene Cooling Events in Malangenfjord and the Adjoining Shelf, North-East Norwegian Sea. Polar Res. 2002, 21, 267–274. [Google Scholar] [CrossRef]
- Husum, K.; Hald, M. Modern Foraminiferal Distribution in the Subarctic Malangen Fjord and Adjoining Shelf, Northern Norway. J. Foraminifer. Res. 2004, 34, 34–48. [Google Scholar] [CrossRef]
- Ernst, S.; van der Zwaan, B. Effects of Experimentally Induced Raised Levels of Organic Flux and Oxygen Depletion on a Continental Slope Benthic Foraminiferal Community. Deep Sea Res. Part I Oceanogr. Res. Pap. 2004, 51, 1709–1739. [Google Scholar] [CrossRef]
- Caulle, C.; Koho, K.A.; Mojtahid, M.; Reichart, G.J.; Jorissen, F.J. Live (Rose Bengal Stained) Foraminiferal Faunas from the Northern Arabian Sea: Faunal Succession within and below the OMZ. Biogeosciences 2014, 11, 1155–1175. [Google Scholar] [CrossRef] [Green Version]
- Ekeroth, N.; Blomqvist, S.; Hall, P.O. Nutrient Fluxes from Reduced Baltic Sea Sediment: Effects of Oxygenation and Macrobenthos. Mar. Ecol. Prog. Ser. 2016, 544, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Keil, R.G.; Montluçon, D.B.; Prahl, F.G.; Hedges, J.I. Sorptive Preservation of Labile Organic Matter in Marine Sediments. Nature 1994, 370, 549–552. [Google Scholar] [CrossRef]
- Knicker, H.; Hatcher, P.G. Survival of Protein in an Organic-Rich Sediment: Possible Protection by Encapsulation in Organic Matter. Naturwissenschaften 1997, 84, 231–234. [Google Scholar] [CrossRef]
- Rohling, E.J.; Marino, G.; Grant, K.M. Mediterranean Climate and Oceanography, and the Periodic Development of Anoxic Events (Sapropels). Earth-Sci. Rev. 2015, 143, 62–97. [Google Scholar] [CrossRef]
- Lourens, L.J. Revised Tuning of Ocean Drilling Program Site 964 and KC01B (Mediterranean) and Implications for the Δ18O, Tephra, Calcareous Nannofossil, and Geomagnetic Reversal Chronologies of the Past 1.1 Myr. Paleoceanography 2004, 19, PA3010. [Google Scholar] [CrossRef]
- Morigi, C. Benthic Environmental Changes in the Eastern Mediterranean Sea during Sapropel S5 Deposition. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 273, 258–271. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Renaud, S.; Bollmann, J. Response of Planktic Foraminiferal Size to Late Quaternary Climate Change. Paleoceanography 2003, 18. [Google Scholar] [CrossRef]
- Richter, R. Aktuopaläontologie Und Paläobiologie, Eine Abgrenzung. Senckenbergiana 1928, 10, 285–292. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cruise | Site | Sea Floor Morphology | Coordinates | Water Depth (m) | Analyses |
|---|---|---|---|---|---|
| GeoB176 | 01 | Grounding-Zone Wedges (GZW) | 74°51.53’ N 16°05.81’ E | 371 | Foraminifera, sediment, biochemistry |
| 07 | Main drift | 74°50.74′ N 17°38.35′ E | 298 | Foraminifera, sediment, biochemistry | |
| 20 | Minor drift | 74°50.74′ N 18°10.53′ E | 333 | Foraminifera, sediment, biochemistry | |
| 22 | Northern Channel | 74°59.69′ N 17°59.59′ E | 167 | Foraminifera sediment | |
| 23 | 75° 0.46′ N 17° 58.85′ E | 150 | Biochemistry |
| Abundance | S | H′ | J | ALD10 | |
|---|---|---|---|---|---|
| (ind./10 cm−2) | (n° Species) | (cm) | |||
| (0–10 cm; >150 μm) | |||||
| 01 | 52 ± 44 | 56 | 3.17 | 0.78 | 2.93 |
| 07 | 91 ± 102 | 38 | 1.89 | 0.52 | 3.06 |
| 20 | 48 ± 62 | 34 | 1.79 | 0.51 | 2.24 |
| 22 | 29 ± 40 | 48 | 2.73 | 0.7 | 1.07 |
| (0–10 cm; 63–150 μm) | |||||
| 01 | 208 ± 300 | 66 | 3.48 | 0.83 | 1.83 |
| 07 | 395 ± 405 | 63 | 3.04 | 0.73 | 2.38 |
| 20 | 126 ± 148 | 55 | 3.16 | 0.79 | 1.64 |
| 22 | 252 ± 364 | 48 | 2.73 | 0.7 | 1.3 |
| (0–10 cm; >63 μm) | |||||
| 01 | 261 ± 342 | 84 | 3.67 | 0.83 | |
| 07 | 487 ± 491 | 56 | 2.81 | 0.7 | |
| 20 | 174 ± 208 | 53 | 2.66 | 0.67 | |
| 22 | 281 ± 402 | 53 | 3.05 | 0.77 |
| PERMANOVA | SIMPER | ||||
|---|---|---|---|---|---|
| P | Dissimilarity | Explanatory Variable | Contribution | Cumulative | |
| (%) | (Taxon) | (%) | (%) | ||
| 01 vs. 07 | ** | 82.55 | Nonionellina labradorica | 9.94 | 9.94 |
| Adercotryma glomeratum | 9.71 | 19.65 | |||
| Nonionella iridea | 5.76 | 25.42 | |||
| Stainforthia fusiformis | 5.53 | 30.95 | |||
| Pullenia quinqueloba | 4.63 | 35.58 | |||
| Melonis zaandami | 4.43 | 40.01 | |||
| Lagenammina sp. D1 | 3.76 | 43.77 | |||
| Alabaminella weddellensis | 3.42 | 47.19 | |||
| Lagenammina difflugiformis | 3.41 | 50.6 | |||
| 01 vs. 20 | * | 83.03 | Nonionellina labradorica | 16.85 | 16.85 |
| Trifarina angulosa | 4.98 | 21.85 | |||
| Melonis zaandami | 4.88 | 26.74 | |||
| Trifarina fluens | 4.57 | 31.31 | |||
| Nonionella iridea | 4.53 | 35.84 | |||
| Lagenammina sp. D1 | 4.49 | 40.33 | |||
| Alabaminella weddellensis | 3.66 | 43.99 | |||
| Lagenammina difflugiformis | 3.34 | 47.32 | |||
| Psammophaga sp. (Artic) | 3.29 | 50.61 | |||
| 01 vs. 22 | ** | 93.45 | Adercotryma glomeratum | 9.38 | 9.384 |
| Cylindrogullmia sp. | 7.13 | 16.52 | |||
| Melonis zaandami | 5.42 | 21.93 | |||
| Trifarina fluens | 4.75 | 26.68 | |||
| Trifarina angulosa | 4.57 | 31.26 | |||
| Lagenammina sp. D1 | 4.50 | 35.75 | |||
| Cuneata arctica | 4.20 | 39.95 | |||
| Nonionella iridea | 3.42 | 43.37 | |||
| Lagenammina difflugiformis | 3.37 | 46.75 | |||
| Islandiella helenae | 3.13 | 49.88 | |||
| Leptohalysis scottii | 2.81 | 52.69 | |||
| 07 vs. 20 | ns | 78.15 | Nonionellina labradorica | 19.88 | 19.88 |
| Adercotryma glomeratum | 8.47 | 28.35 | |||
| Stainforthia fusiformis | 5.47 | 33.83 | |||
| Nonionella iridea | 5.02 | 38.84 | |||
| Pullenia quinqueloba | 4.56 | 43.41 | |||
| Melonis zaandami | 4.13 | 47.54 | |||
| Bolivinellina pseudopunctata | 3.44 | 50.98 | |||
| 07 vs. 22 | * | 84.86 | Adercotryma glomeratum | 13.07 | 13.07 |
| Nonionellina labradorica | 10.23 | 23.29 | |||
| Cylindrogullmia sp. | 6.51 | 29.8 | |||
| Stainforthia fusiformis | 5.45 | 35.25 | |||
| Pullenia quinqueloba | 4.57 | 39.82 | |||
| Nonionella iridea | 3.96 | 43.78 | |||
| Bolivinellina pseudopunctata | 3.77 | 47.55 | |||
| Melonis zaandami | 3.73 | 51.28 | |||
| 20 vs. 22 | * | 87.89 | Nonionellina labradorica | 16.25 | 16.25 |
| Adercotryma glomeratum | 10.35 | 26.60 | |||
| Cylindrogullmia sp. | 7.37 | 33.97 | |||
| Cuneata arctica | 5.04 | 39.01 | |||
| Trifarina angulosa | 3.65 | 42.66 | |||
| Trifarina fluens | 3.46 | 46.12 | |||
| Psammophaga sp. (Artic) | 3.42 | 49.53 | |||
| Reophax scorpiurus | 3.21 | 52.74 |
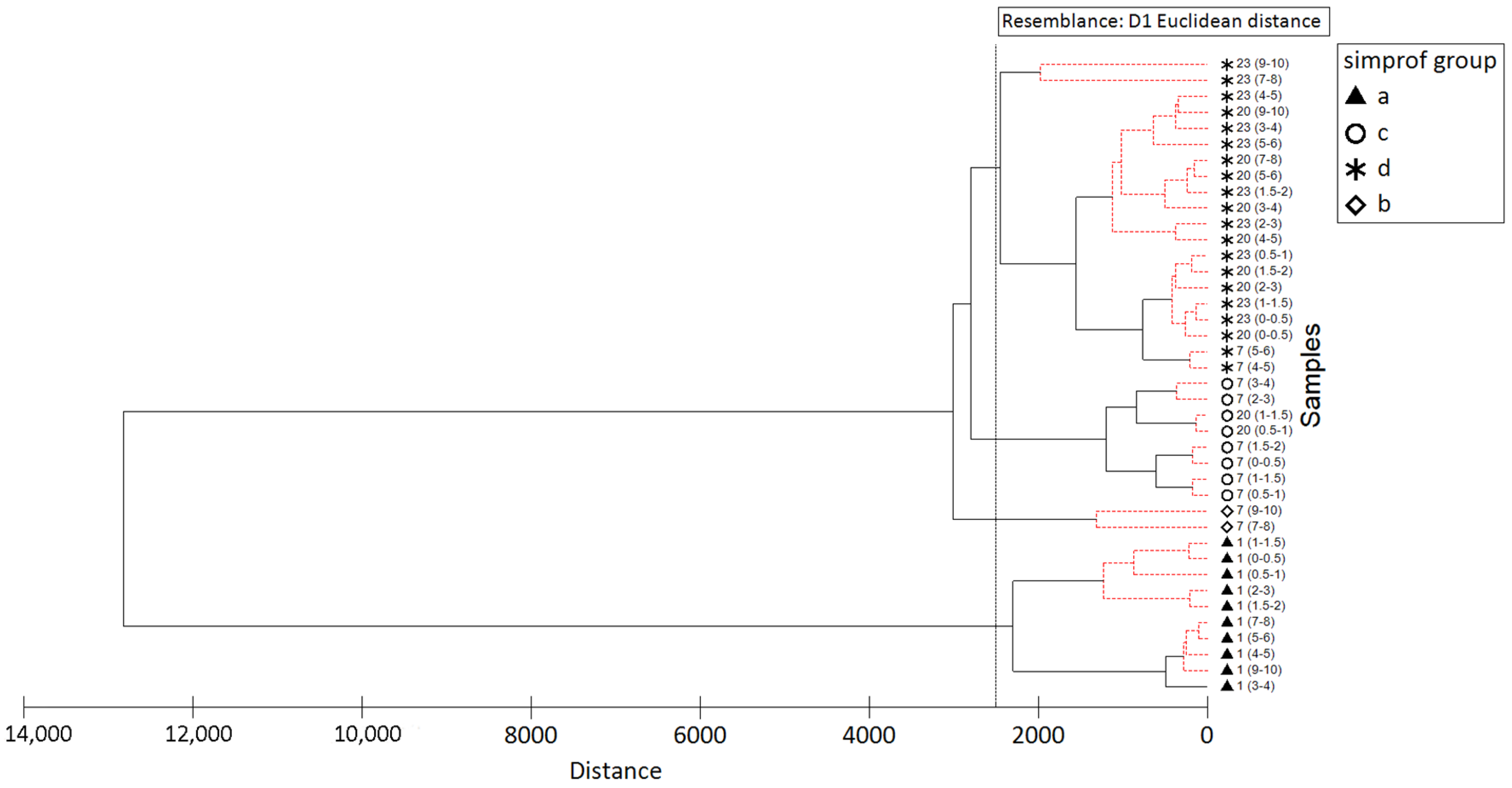
| Variable | Simprof Group | A | B | C | D |
|---|---|---|---|---|---|
| Sedimentary Organic Matter | C-CHO-H2O (μgC/g) | 37.76 | 204.00 | 165.25 | 141.57 |
| C-CHO-EDTA (μgC/g) | 107.57 | 657.50 | 362.50 | 280.50 | |
| C-PRT (μgC/g) | 353.27 | 2940.50 | 1132.00 | 1630.29 | |
| C-LIP (μgC/g) | 460.56 | 717.00 | 865.50 | 806.44 | |
| TOC (μgC/g) | 8455.69 | 21,975.25 | 22,731.09 | 20,088.19 | |
| BCP (μgC/g) | 959.17 | 4519.27 | 2525.27 | 2858.86 | |
| TN (μgN/g) | 1002.20 | 2757.53 | 2911.28 | 2594.60 | |
| N-PRT/TN | 0.11 | 0.34 | 0.12 | 0.20 | |
| TOC/TN | 9.84 | 9.29 | 9.11 | 9.04 | |
| Foraminiferal Assemblage | Total species richness (S) | 82 | 20 | 74 | 90 |
| N. of exclusive species | 23 | 1 | 12 | 17 | |
| Most caracteristics species | Melonis zaandami | Adercotryma glomeratum | Nonionellina labradorica | Adercotryma glomeratum | |
| (25% similarity within group) | Lagenammina sp. D1 | Trifarina fluens | Stainforthia fusiformis | Cylindrogullmia sp. | |
| Lagenammina difflugiformis | Reophax scorpiurus | Nonionella iridea | Cuneata arctica |
| Variable | F | P | Variance (%) | Variance Cumulative (%) | |
|---|---|---|---|---|---|
| (a) | N-PRT/TN | 5.04 | *** | 12 | 12 |
| BCP | 7.26 | *** | 14 | 26 | |
| C-PRT | 3.13 | *** | 6 | 32 | |
| TN | 1.68 | ns | 3 | 35 | |
| TOC | 2.38 | ** | 4 | 39 | |
| TOC/TN | 2.00 | ** | 3 | 43 | |
| C-LIP | 1.73 | * | 3 | 46 | |
| C-CHO-EDTA | 0.90 | ns | 2 | 47 | |
| C-CHO-H2O | 0.53 | ns | 1 | 48 | |
| (b) | N-PRT/TN | 13.98 | *** | 27 | 27 |
| C-LIP | 15.72 | ** | 22 | 49 | |
| C-CHO-EDTA | 2.17 | ns | 3 | 52 | |
| C-PRT | 12.65 | ** | 13 | 65 | |
| TOC/TN | 2.30 | ns | 2 | 67 | |
| C-CHO-H2O | 2.00 | ns | 2 | 69 | |
| BCP | 1.73 | ns | 2 | 70 | |
| TOC | 0.81 | ns | 1 | 71 | |
| TN | 6.29 | * | 5 | 76 | |
| (c) | C-LIP | 7.78 | *** | 17 | 17 |
| C-PRT | 6.64 | *** | 13 | 30 | |
| N-PRT/TN | 4.97 | *** | 9 | 38 | |
| BCP | 4.44 | *** | 9 | 45 | |
| TOC/TN | 1.57 | ns | 2 | 48 | |
| TOC | 1.27 | ns | 2 | 49 | |
| TN | 3.37 | ** | 5 | 54 | |
| C-CHO-EDTA | 0.83 | ns | 1 | 55 | |
| C-CHO-H2O | 0.65 | ns | 1 | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbatini, A.; Bazzaro, M.; Caridi, F.; De Vittor, C.; Esposito, V.; Lucchi, R.G.; Negri, A.; Morigi, C. Benthic Foraminifera and Productivity Regimes in the Kveithola Trough (Barents Sea)—Ecological Implications in a Changing Arctic and Actuopaleontological Meaning. J. Mar. Sci. Eng. 2023, 11, 237. https://doi.org/10.3390/jmse11020237
Sabbatini A, Bazzaro M, Caridi F, De Vittor C, Esposito V, Lucchi RG, Negri A, Morigi C. Benthic Foraminifera and Productivity Regimes in the Kveithola Trough (Barents Sea)—Ecological Implications in a Changing Arctic and Actuopaleontological Meaning. Journal of Marine Science and Engineering. 2023; 11(2):237. https://doi.org/10.3390/jmse11020237
Chicago/Turabian StyleSabbatini, Anna, Matteo Bazzaro, Francesca Caridi, Cinzia De Vittor, Valentina Esposito, Renata Giulia Lucchi, Alessandra Negri, and Caterina Morigi. 2023. "Benthic Foraminifera and Productivity Regimes in the Kveithola Trough (Barents Sea)—Ecological Implications in a Changing Arctic and Actuopaleontological Meaning" Journal of Marine Science and Engineering 11, no. 2: 237. https://doi.org/10.3390/jmse11020237
APA StyleSabbatini, A., Bazzaro, M., Caridi, F., De Vittor, C., Esposito, V., Lucchi, R. G., Negri, A., & Morigi, C. (2023). Benthic Foraminifera and Productivity Regimes in the Kveithola Trough (Barents Sea)—Ecological Implications in a Changing Arctic and Actuopaleontological Meaning. Journal of Marine Science and Engineering, 11(2), 237. https://doi.org/10.3390/jmse11020237