
Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Experimental Preparation
2.2. Single Light Wavelength Experiment
2.3. Light Wavelength Change Experiment
2.4. Statistical Analysis
3. Results
3.1. Single Light Wavelength Experiment
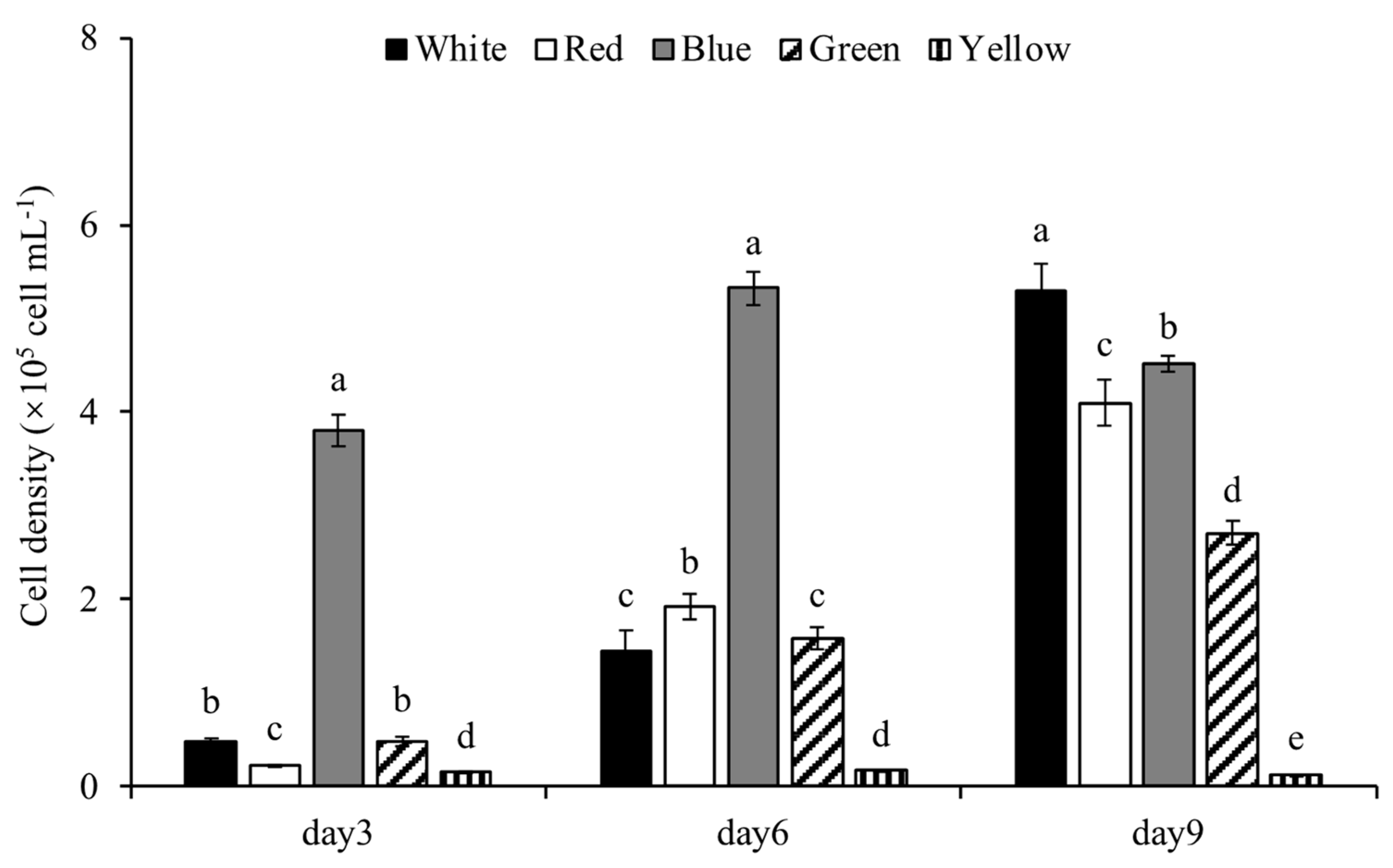
3.1.1. Cell Density
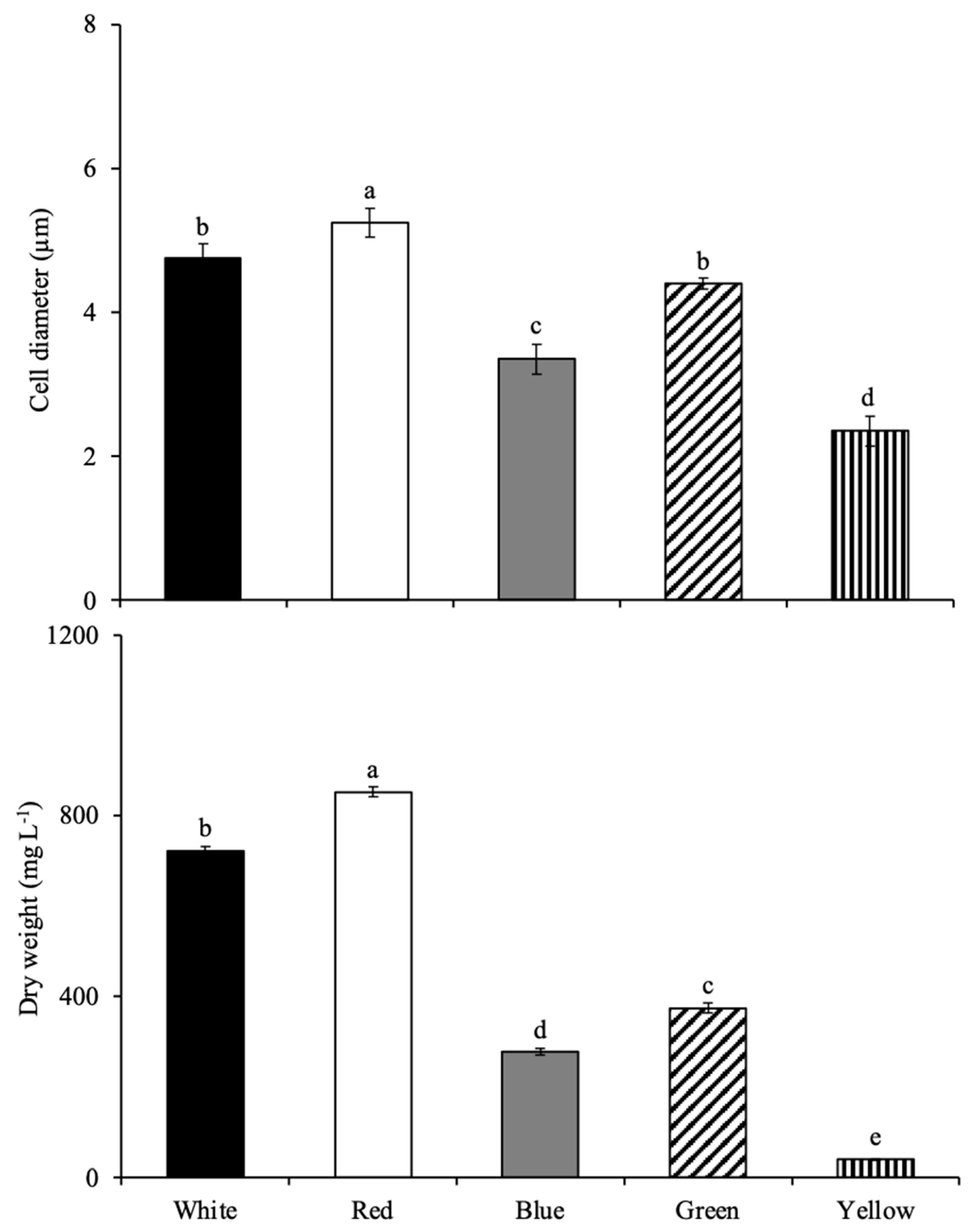
3.1.2. Growth
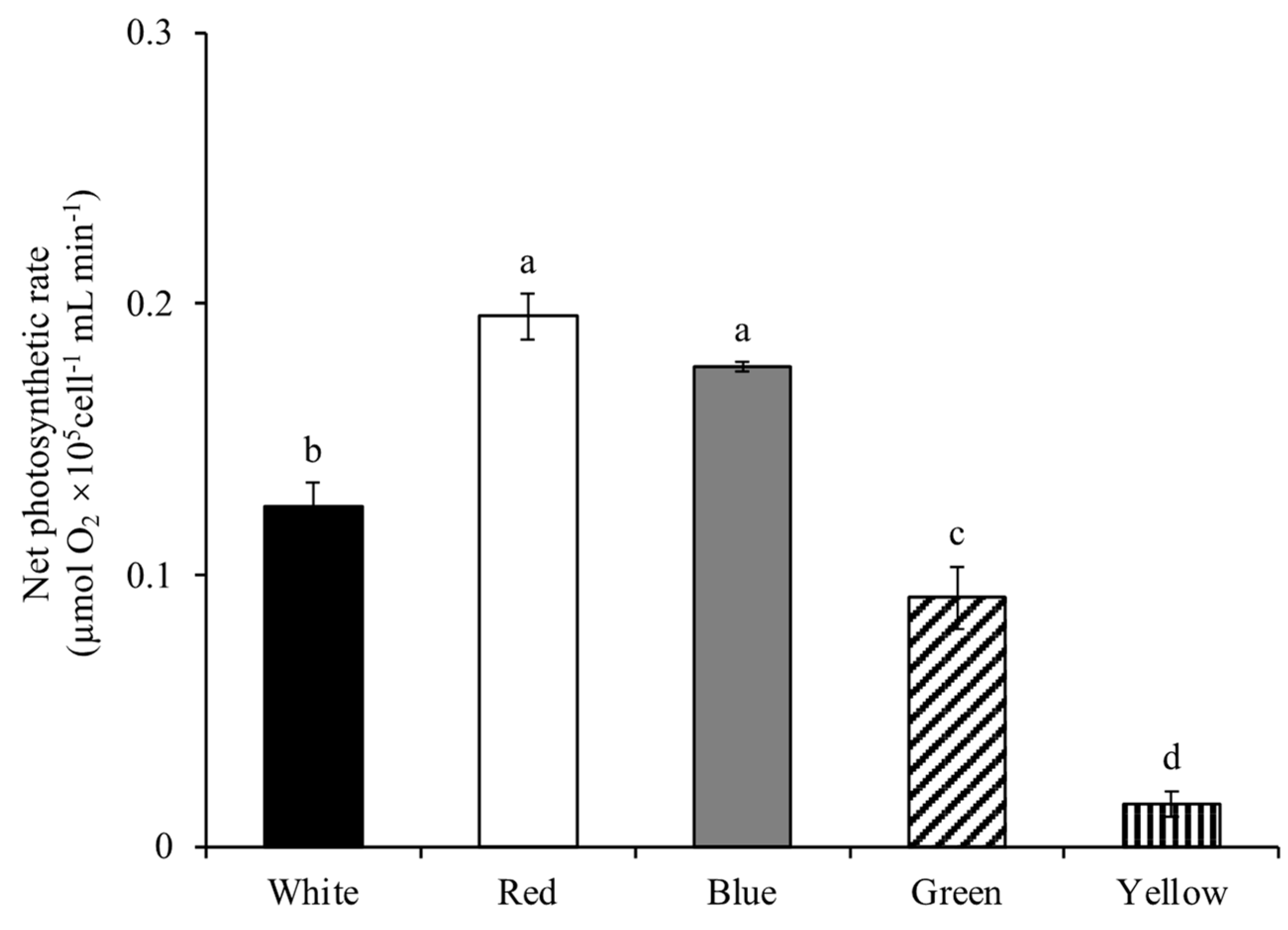
3.1.3. Net Photosynthetic Rate
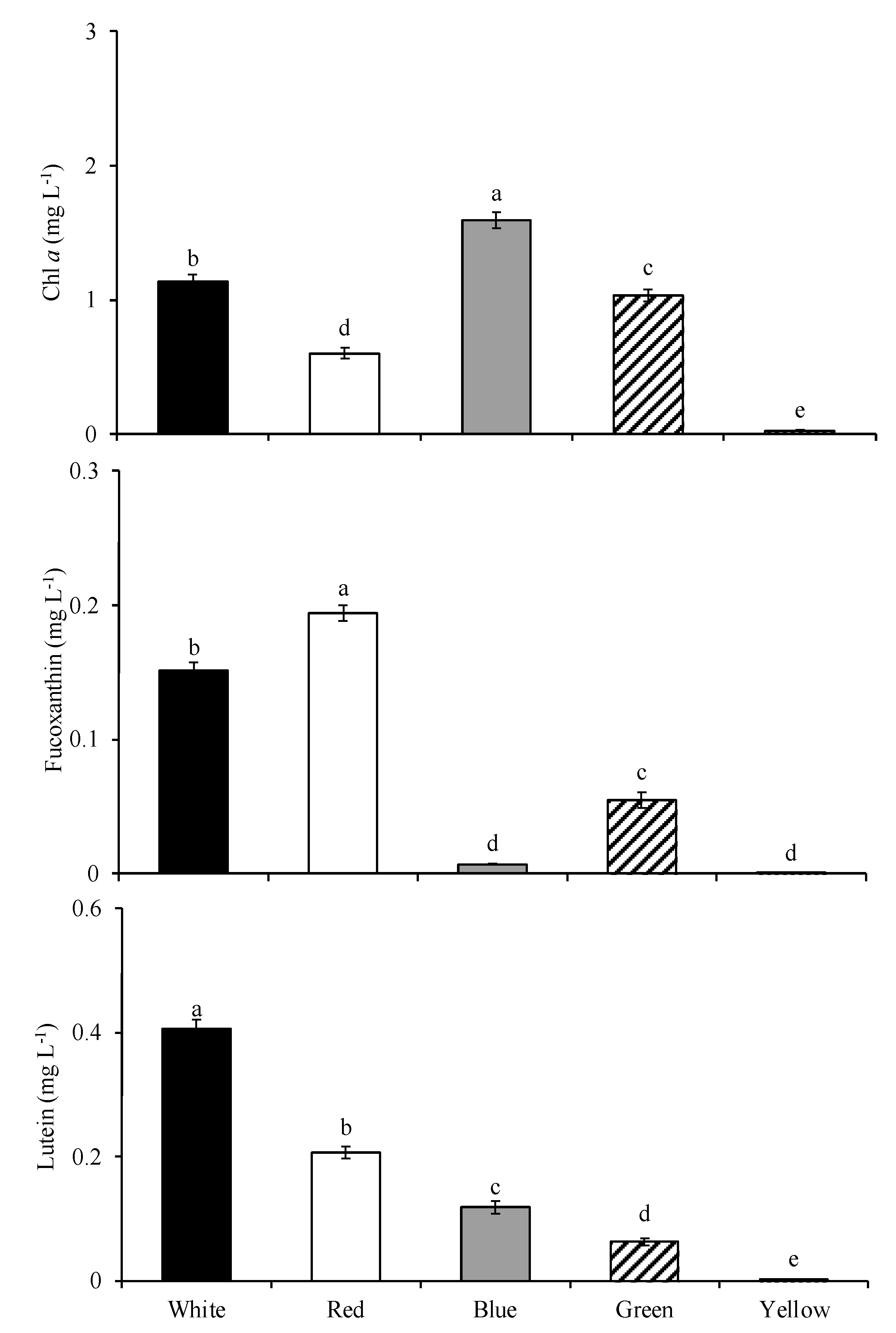
3.1.4. Pigment Contents
3.2. Light Wavelength Change Experiment
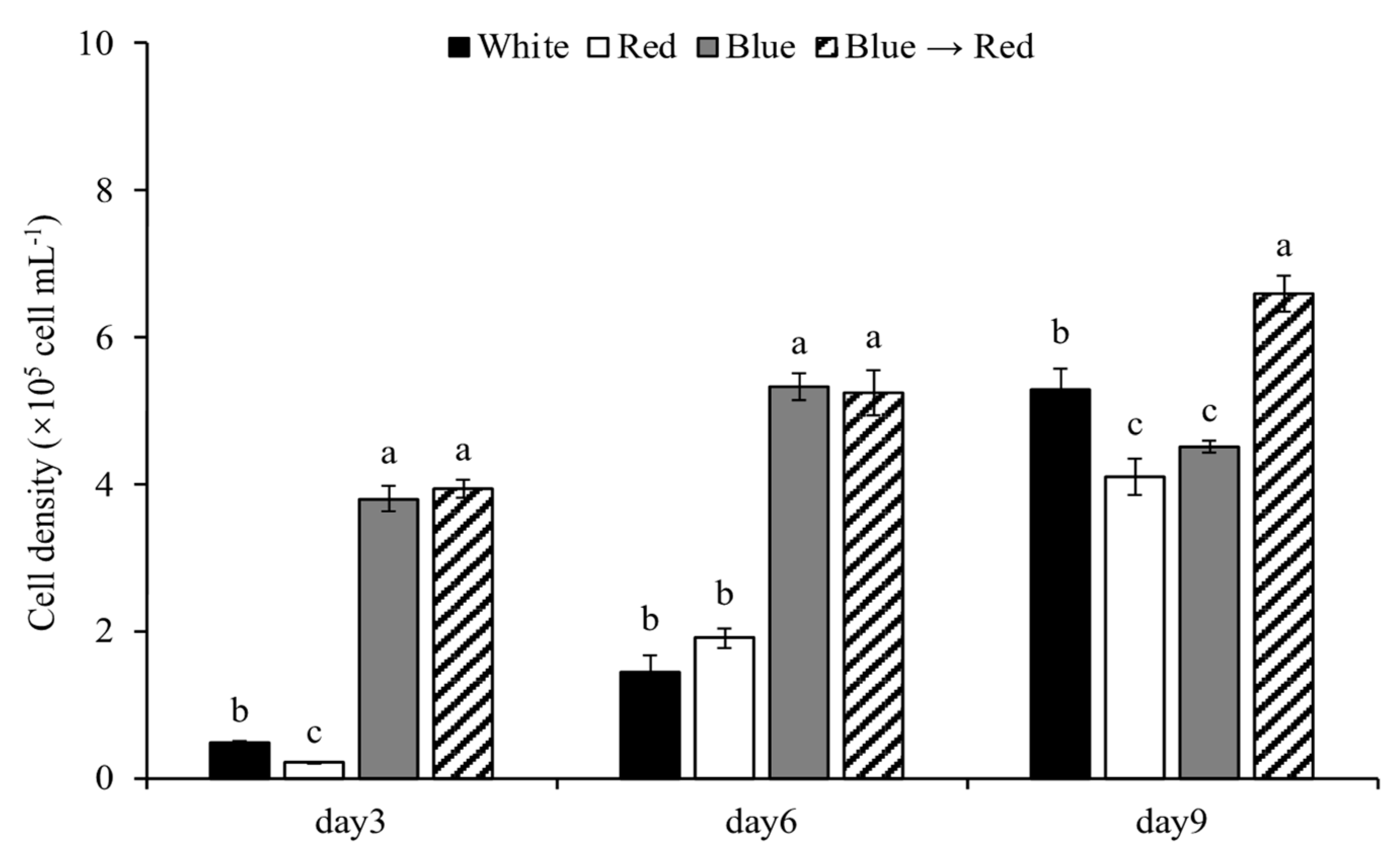
3.2.1. Cell Density
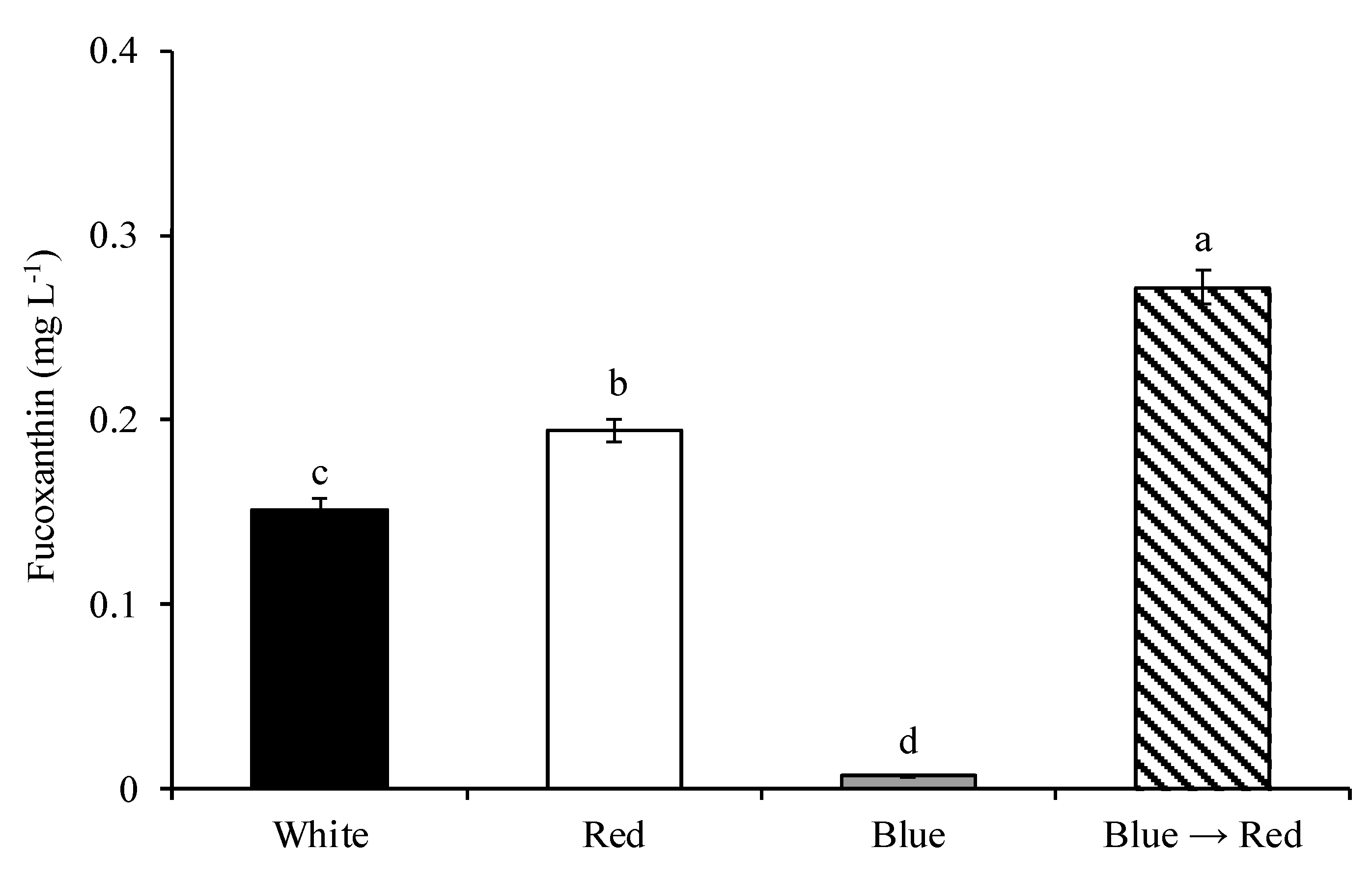
3.2.2. Fucoxanthin Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, R.E. Phycology, 5th ed.; Cambridge University Press: Cambridge, UK, 2018; pp. 492–506. [Google Scholar]
- Ramanna, L.; Rawat, I.; Bux, F. Light enhancement strategies improve microalgal biomass productivity. Renew. Sust. Energ. Rev. 2017, 80, 765–773. [Google Scholar] [CrossRef]
- Ma, S.; Zeng, W.; Huang, Y.; Zhu, X.; Xia, A.; Zhu, X.; Liao, Q. Revealing the synergistic effects of cells, pigments, and light spectra on light transfer during microalgae growth: A comprehensive light attenuation model. Bioresour. Technol. 2022, 348, 126777. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Sánchez-Saavedra, M.; Maeda-Martínez, A.N.; Acosta-Galindo, S. Effect of different light spectra on the growth and biochemical composition of Tisochrysis lutea. J. Appl. Phycol. 2016, 28, 839–847. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Liu, J.; Qin, R. Light absorption and growth response of Dunaliella under different light qualities. J. Appl. Phycol. 2020, 32, 1041–1052. [Google Scholar] [CrossRef]
- Li, X.; Manuel, J.; Crunkleton, D.W.; Johannes, T.W. Effect of blue and red-orange LEDs on the growth and biochemical profile of Chlamydomonas reinhardtii. J. Appl. Phycol. 2021, 33, 1367–1377. [Google Scholar] [CrossRef]
- Mercado, J.M.; del Pilar Sánchez-Saavedra, M.; Correa-Reyes, G.; Lubián, L.; Montero, O.; Figueroa, F.L. Blue light effect on growth, light absorption characteristics and photosynthesis of five benthic diatom strains. Aquat. Bot. 2004, 78, 265–277. [Google Scholar] [CrossRef]
- Marchetti, J.; Bougaran, G.; Jauffrais, T.; Lefebvre, S.; Rouxel, C.; Saint-Jean, B.; Lukomska, E.; Robert, R.; Cadoret, J.P. Effect of blue light on the biochemical composition and photosynthetic activity of Isochrysis sp. (T-iso). J. Appl. Phycol. 2013, 25, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Ying, K.; Wu, K.; Tang, S.; Zhou, J.; Cai, Z. Promoting the growth of Haematococcus lacustris under high light intensity through the combination of light/dark cycle and light color. J. Mar. Sci. Eng. 2022, 10, 839. [Google Scholar] [CrossRef]
- Romero-Romero, C.C.; Sánchez-Saavedra, M. Effect of light quality on the growth and proximal composition of Amphora sp. J. Appl. Phycol. 2017, 29, 1203–1211. [Google Scholar] [CrossRef]
- Zhao, X.; Ma, R.; Liu, X.; Ho, S.H.; Xie, Y.; Chen, J. Strategies related to light quality and temperature to improve lutein production of marine microalga Chlamydomonas sp. Bioproc. Biosyst. Eng. 2019, 42, 435–443. [Google Scholar] [CrossRef]
- Li, D.; Yuan, Y.; Cheng, D.; Zhao, Q. Effect of light quality on growth rate, carbohydrate accumulation, fatty acid profile and lutein biosynthesis of Chlorella sp. AE10. Bioresour. Technol. 2019, 291, 121783. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.; da Conceição, D.P.; Corrêa, D.O.; Passos, M.F.; Campos, M.P.; Adamoski, D.; Galli-Terasawa, L.V.; Mariano, A.B.; Vargas, J.V.; Kava, V.M. Changes in gene expression and biochemical composition of Haematococcus pluvialis grown under different light colors. J. Appl. Phycol. 2022, 34, 729–743. [Google Scholar] [CrossRef]
- Pereira, S.; Otero, A. Haematococcus pluvialis bioprocess optimization: Effect of light quality, temperature and irradiance on growth, pigment content and photosynthetic response. Algal Res. 2020, 51, 102027. [Google Scholar] [CrossRef]
- Borella, L.; Diotto, D.; Barbera, E.; Fiorimonte, D.; Sforza, E.; Trivellin, N. Application of flashing blue-red LED to boost microalgae biomass productivity and energy efficiency in continuous photobioreactors. Energy 2022, 259, 125087. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, Y.; Zhang, Y.; Zhang, S.; Qu, J.; Wang, X.; Kong, R.; Han, C.; Liu, Z. Fucoxanthin: A promising medicinal and nutritional ingredient. Evid-Based Compl. Alt. 2015, 2015, 723515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarekarizi, A.; Hoffmann, L.; Burritt, D. Approaches for the sustainable production of fucoxanthin, a xanthophyll with potential health benefits. J. Appl. Phycol. 2019, 31, 281–299. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Y.; He, Y.; Liu, B.; Mou, H.; Chen, F.; Yang, S. Microalgae-Derived Pigments for the Food Industry. Mar. Drugs 2022, 21, 82. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Yang, B.; Sun, P.; Lu, X.; Liu, J.; Chen, F. Screening of diatom strains and characterization of Cyclotella cryptica as a potential fucoxanthin producer. Mar. Drugs 2016, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, H.; Wu, T.; Fu, Y.; He, Y.; Mao, X.; Chen, F. Storage carbon metabolism of Isochrysis zhangjiangensis under different light intensities and its application for co-production of fucoxanthin and stearidonic acid. Bioresour. Technol. 2019, 282, 94–102. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Webb, J.P.; Borowitzka, M.A. Bioremediation and other potential applications of coccolithophorid algae: A review. Algal Res. 2012, 1, 120–133. [Google Scholar] [CrossRef]
- Winter, A.; Henderiks, J.; Beaufort, L.; Rickaby, R.E.; Brown, C.W. Poleward expansion of the coccolithophore Emiliania huxleyi. J. Plankton Res. 2014, 36, 316–325. [Google Scholar] [CrossRef]
- Perrin, L.; Probert, I.; Langer, G.; Aloisi, G. Growth of the coccolithophore Emiliania huxleyi in light- and nutrient-limited batch reactors: Relevance for the BIOSOPE deep ecological niche of coccolithophores. Biogeosciences 2016, 13, 5983–6001. [Google Scholar] [CrossRef] [Green Version]
- Langer, G.; Nehrke, G.; Probert, I.; Ly, J.; Ziveri, P. Strain-specific responses of Emiliania huxleyi to changing seawater carbonate chemistry. Biogeosciences 2009, 6, 2637–2646. [Google Scholar] [CrossRef] [Green Version]
- Puig-Fàbregas, J.; Alcaraz-Rocha, P.; Fernández, E.; Rotllant, J.; Sobrino, C. Evaluation of actin as a reference for quantitative gene expression studies in Emiliania huxleyi (Prymnesiophyceae) under ocean acidification conditions. Phycologia 2021, 60, 148–157. [Google Scholar] [CrossRef]
- Patil, S.M.; Mohan, R.; Shetye, S.S.; Vaz, V.; Gazi, S.; Choudhari, P.P.; Jafar, S.A. Emiliania huxleyi biometry and calcification response to the Indian sector of the Southern Ocean environmental gradients. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 585, 110725. [Google Scholar] [CrossRef]
- Bakku, R.K.; Araie, H.; Hanawa, Y.; Shiraiwa, Y.; Suzuki, I. Changes in the accumulation of alkenones and lipids under nitrogen limitation and its relation to other energy storage metabolites in the haptophyte alga Emiliania huxleyi CCMP 2090. J. Appl. Phycol. 2018, 30, 23–36. [Google Scholar] [CrossRef]
- Aveiro, S.S.; Melo, T.; Figueiredo, A.; Domingues, P.; Pereira, H.; Maia, I.B.; Silva, J.; Domingues, R.M.; Nunes, C.; Moreira, A.S. The polar lipidome of cultured Emiliania huxleyi: A source of bioactive lipids with relevance for biotechnological applications. Biomolecules 2020, 10, 1434. [Google Scholar] [CrossRef]
- Vicente, B.; Matos, J.; Gomes, R.; Sapatinha, M.; Afonso, C.; Rodrigues, T.; Amorim, A.; Bandarra, N.M.; Cardoso, C. Production and bioaccessibility of Emiliania huxleyi biomass and bioactivity of its aqueous and ethanolic extracts. J. Appl. Phycol. 2021, 33, 3719–3729. [Google Scholar] [CrossRef]
- Thierstein, H.R.; Young, J.R. Coccolithophores: From Molecular Processes to Global Impact, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 51–73. [Google Scholar]
- Gebühr, C.; Sheward, R.M.; Herrle, J.O.; Bollmann, J. Strain-specific morphological response of the dominant calcifying phytoplankton species Emiliania huxleyi to salinity change. PLoS ONE 2021, 16, e0246745. [Google Scholar] [CrossRef]
- Lai, Y.S.; Eustance, E.; Shesh, T.; Frias, Z.; Rittmann, B.E. Achieving superior carbon transfer efficiency and pH control using membrane carbonation with a wide range of CO2 contents for the coccolithophore Emiliania huxleyi. Sci. Total Environ. 2022, 822, 153592. [Google Scholar] [CrossRef]
- Rigual-Hernández, A.S.; Sierro, F.J.; Flores, J.A.; Trull, T.W.; Rodrigues, T.; Martrat, B.; Sikes, E.L.; Nodder, S.D.; Eriksen, R.S.; Davies, D.; et al. Influence of environmental variability and Emiliania huxleyi ecotypes on alkenone-derived temperature reconstructions in the subantarctic Southern Ocean. Sci. Total Environ. 2022, 812, 152474. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, V.; León, P.; Gordillo, F.J.; Jiménez, C.; Concepción, I.; Mackenzie, K.; Bresnan, E.; Segovia, M. High-CO2 levels rather than acidification restrict Emiliania huxleyi growth and performance. Microb. Ecol. 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Eppley, R.W.; Holmes, R.W.; Strickland, J.D. Sinking rates of marine phytoplankton measured with a fluorometer. J. Exp. Mar. Biol. Ecol. 1967, 1, 191–208. [Google Scholar] [CrossRef]
- LeGresley, M.; McDermott, G. Counting chamber methods for quantitative phytoplankton analysis—Haemocytometer, Palmer-Maloney cell and Sedgewick-Rafter cell. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; UNESCO: Paris, France, 2010; pp. 25–30. [Google Scholar]
- Azov, Y. Effect of pH on inorganic carbon uptake in algal cultures. Appl. Environ. Microbiol. 1982, 43, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Zang, Z.; Xie, X.; Huang, A.; Wang, G. The influence of different flocculants on the physiological activity and fucoxanthin production of Phaeodactylum tricornutum. Process Biochem. 2014, 49, 681–687. [Google Scholar] [CrossRef]
- Teo, C.L.; Atta, M.; Bukhari, A.; Taisir, M.; Yusuf, A.M.; Idris, A. Enhancing growth and lipid production of marine microalgae for biodiesel production via the use of different LED wavelengths. Bioresour. Technol. 2014, 162, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Latsos, C.; van Houcke, J.; Blommaert, L.; Verbeeke, G.P.; Kromkamp, J.; Timmermans, K.R. Effect of light quality and quantity on productivity and phycoerythrin concentration in the cryptophyte Rhodomonas sp. J. Appl. Phycol. 2021, 33, 729–741. [Google Scholar] [CrossRef]
- Gorai, T.; Katayama, T.; Obata, M.; Murata, A.; Taguchi, S. Low blue light enhances growth rate, light absorption, and photosynthetic characteristics of four marine phytoplankton species. J. Exp. Mar. Biol. Ecol. 2014, 459, 87–95. [Google Scholar] [CrossRef]
- Vadiveloo, A.; Moheimani, N.R.; Cosgrove, J.J.; Bahri, P.A.; Parlevliet, D. Effect of different light spectra on the growth and productivity of acclimated Nannochloropsis sp.(Eustigmatophyceae). Algal Res. 2015, 8, 121–127. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, W.; Guðmundsson, Ó.; Paglia, G.; Herjólfsson, G.; Andrésson, Ó.S.; Palsson, B.Ø.; Brynjólfsson, S. Enhancement of carotenoid biosynthesis in the green microalga Dunaliella salina with light-emitting diodes and adaptive laboratory evolution. Appl. Microbiol. Biotechnol. 2013, 97, 2395–2403. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Harvey, P.J. Carotenoid production by Dunaliella salina under red light. Antioxidants 2019, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Pagels, F.; Bonomi-Barufi, J.; Vega, J.; Abdala-Díaz, R.; Vasconcelos, V.; Guedes, A.; Figueroa, F.L. Light quality triggers biochemical modulation of Cyanobium sp.—Photobiology as tool for biotechnological optimization. J. Appl. Phycol. 2020, 32, 2851–2861. [Google Scholar] [CrossRef]
- Amaro, H.M.; Pagels, F.; Azevedo, I.C.; Azevedo, J.; Sousa Pinto, I.; Malcata, F.X.; Guedes, A. Light-emitting diodes—A plus on microalgae biomass and high-value metabolite production. J. Appl. Phycol. 2020, 32, 3605–3618. [Google Scholar] [CrossRef]
- Garrido, J.L.; Brunet, C.; Rodríguez, F. Pigment variations in Emiliania huxleyi (CCMP370) as a response to changes in light intensity or quality. Environ. Microbiol. 2016, 18, 4412–4425. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, S.C.; Harris, G.; Webster, R.; Leonardos, N.; Geider, R.J.; Raines, C.A. Characterization and expression analysis of the LHCF gene family in Emiliania huxleyi (haptophyta) reveals differential responses to light and CO2. J. Phycol. 2010, 46, 123–134. [Google Scholar] [CrossRef]
- Yan, C.; Luo, X.; Zheng, Z. Effects of various LED light qualities and light intensity supply strategies on purification of slurry from anaerobic digestion process by Chlorella vulgaris. Int. Biodeterior. Biodegradation 2013, 79, 81–87. [Google Scholar] [CrossRef]
- Ra, C.H.; Kang, C.H.; Jung, J.H.; Jeong, G.T.; Kim, S.K. Effects of light-emitting diodes (LEDs) on the accumulation of lipid content using a two-phase culture process with three microalgae. Bioresour. Technol. 2016, 212, 254–261. [Google Scholar] [CrossRef]
- Su, Y.; Lundholm, N.; Friis, S.M.; Ellegaard, M. Implications for photonic applications of diatom growth and frustule nanostructure changes in response to different light wavelengths. Nano Res. 2015, 8, 2363–2372. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Fleurent, G.; Awwad, F.; Cheng, M.; Meddeb-Mouelhi, F.; Budge, S.M.; Germain, H.; Desgagné-Penix, I. Red light variation an effective alternative to regulate biomass and lipid profiles in Phaeodactylum tricornutum. Appl. Sci. 2020, 10, 2531. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Zheng, Z. Performance of mixed LED light wavelengths on biogas upgrade and biogas fluid removal by microalga Chlorella sp. Appl. Energy 2014, 113, 1008–1014. [Google Scholar] [CrossRef]
- Sirisuk, P.; Ra, C.H.; Jeong, G.T.; Kim, S.K. Effects of wavelength mixing ratio and photoperiod on microalgal biomass and lipid production in a two-phase culture system using LED illumination. Bioresour. Technol. 2018, 253, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Raqiba, H.; Sibi, G. Light emitting diode (LED) illumination for enhanced growth and cellular composition in three microalgae. Adv. Microb. Res. 2019, 3, 7. [Google Scholar]
- Kim, S.H.; Sunwoo, I.Y.; Hong, H.J.; Awah, C.C.; Jeong, G.T.; Kim, S.K. Lipid and unsaturated fatty acid productions from three microalgae using nitrate and light-emitting diodes with complementary LED wavelength in a two-phase culture system. Bioprocess Biosyst. Eng. 2019, 42, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Jungandreas, A.; Costa, B.S.; Jakob, T.; Von Bergen, M.; Baumann, S.; Wilhelm, C. The acclimation of Phaeodactylum tricornutum to blue and red light does not influence the photosynthetic light reaction but strongly disturbs the carbon allocation pattern. PLoS ONE 2014, 9, e99727. [Google Scholar] [CrossRef] [Green Version]
- Granata, T.; Habermacher, P.; Härri, V.; Egli, M. The influence of bio-optical properties of Emiliania huxleyi and Tetraselmis sp. on biomass and lipid production when exposed to different light spectra and intensities of an adjustable LED array and standard light sources. SN Appl. Sci. 2019, 1, 524. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Paglia, G.; Magnúsdóttir, M.; Steinarsdóttir, E.A.; Gudmundsson, S.; Palsson, B.Ø.; Andrésson, Ó.S.; Brynjólfsson, S. Effects of abiotic stressors on lutein production in the green microalga Dunaliella salina. Microb. Cell Fact. 2014, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Liu, F.; Wang, Q.; Gong, Q.; Gao, X. Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae). J. Mar. Sci. Eng. 2023, 11, 456. https://doi.org/10.3390/jmse11020456
Zhang J, Liu F, Wang Q, Gong Q, Gao X. Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae). Journal of Marine Science and Engineering. 2023; 11(2):456. https://doi.org/10.3390/jmse11020456
Chicago/Turabian StyleZhang, Jing, Falong Liu, Qiaohan Wang, Qingli Gong, and Xu Gao. 2023. "Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae)" Journal of Marine Science and Engineering 11, no. 2: 456. https://doi.org/10.3390/jmse11020456
APA StyleZhang, J., Liu, F., Wang, Q., Gong, Q., & Gao, X. (2023). Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae). Journal of Marine Science and Engineering, 11(2), 456. https://doi.org/10.3390/jmse11020456