
Plankton Genes and Extracellular Organic Substances in the Ocean


Abstract
:1. Introduction
2. Inorganic Matter (IM) and Organic Matter (OM) in the Oceans
2.1. Sources of Available N and Fe
2.2. Origins and Classes of Allochthonous IM and OM
2.3. Origins and Classes of Autochthonous OM
2.4. Autochthonous Particulate Organic Matter (POM)
2.5. Autochthonous Dissolved Organic Matter (DOM)
3. DOM with Signaling and Allelopathic Functions
4. Molecules in Intraspecific and Interspecific Signaling
5. Dimethylsulfide (DMS) in Signaling and Structuring Consortia
6. Consortia Structured by Rheological Properties, including Stickiness, of Polymers
7. Prey-Capture and Predator-Avoidance
8. Predator–Prey Interactions
9. Mucus Trophic Structures (“Mucus Traps”)
10. Mucus as a Retention Tool
11. The Roles of Cross-Linked Gels, Rheological Changes and Reactive Oxygen Species in Toxicity to Fish
- Zhang and Zhang [106], using a mechanochemical technique, single-molecule force spectroscopy with an atomic force microscope (AFM), explored the relation between rate-of-strain and opposing stress in pulling and breaking single molecular bonds in both polysaccharides and proteins. For example, they found breaking forces in the typically algal polysaccharides, λ-, β-, and ι-carrageenan all to be around 500 to 700 pN.
- In contrast, disulfide links in protein were found by Wiita et al. [107] to have a lower breaking strength of around 100 to 200 pN, and they can be weakened by dithiothreitol (DTT). Disulfide links are important as they strengthen both proteins [108,109] and polysaccharides [109] in biological systems by forming cross-links between polymer chains.
12. Mechanisms of Killing Microbes
13. Quorum Sensing
14. Scales (Granulometry) of Toxicity
15. Hydrophobicity, Organic Aggregate Size and Rheology
16. Molecules, Produced by other Organisms and Associated Bacteria, That Are Toxic and Allelopathic to Phytoplankton
17. Organic Polymers and Gaia
18. Polymer Modulation of Fluxes: Discovering the Genomes Involved
19. Vertical Organic Flux of OM: Succinct Examples of Modulation by Polymers
20. Vertical Organic Flux between the Photic Zone and the Deep Ocean: The Bigger Picture
21. Ocean Foam
22. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hansell, D.A.; Carlson, C.A.; Repeta, D.J.; Schlitzer, R. Dissolved organic matter in the ocean: A controversy stimulates new insights. Oceanography 2009, 22, 202–211. [Google Scholar] [CrossRef] [Green Version]
- McCarren, J.; Becker, J.W.; Repeta, D.J.; Shi, Y.; Young, C.R.; Malmstrom, R.R.; Chisholm, S.W.; DeLong, E.F. Microbial community transcriptomes reveal microbes and metabolic pathways associated with dissolved organic matter turnover in the sea. Proc. Natl. Acad. Sci. USA 2010, 107, 16420–16427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, N.; Herndl, G.J.; Hansell, D.A.; Benner, R.; Kattner, G.; Wilhelm, S.W.; Kirchman, D.L.; Weinbauer, M.G.; Luo, T.; Chen, F.; et al. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Genet. 2010, 8, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Sun, X.X.; Seuront, L. Thalassorheology, organic matter and plankton: Towards a more viscous approach in plankton ecology. J. Plankton Res. 2015, 37, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Benner, R.; Amon, R.M. The size-reactivity continuum of major bioelements in the ocean. Annu. Rev. Mar. Sci. 2015, 7, 185–205. [Google Scholar] [CrossRef]
- Brown, E.R.; Cepeda, M.R.; Mascuch, S.J.; Poulson-Ellestad, K.L.; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep. 2019, 36, 1093–1116. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Shikata, T.; Nukata, A.; Ichiki, S.; Nagasoe, S.; Matsubara, T.; Shimasaki, Y.; Nakao, M.; Yamaguchi, K.; Oshima, Y.; et al. Extracellular polysaccharide-protein complexes of a harmful alga mediate the allelopathic control it exerts within the phytoplankton community. ISME J. 2009, 3, 808–817. [Google Scholar] [CrossRef] [Green Version]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent exopolymer particles: Effects on carbon cycling in the ocean. Prog. Oceanogr. 2016, 151, 13–37. [Google Scholar] [CrossRef] [Green Version]
- Wurl, O.; Ekau, W.; Landing, W.M.; Zappa, C.J. Sea surface microlayer in a changing ocean—A perspective. Elem. Sci. Anthr. 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Berdalet, E.; Chin, W.-C.; Denis, M.; Ding, H.; Duan, J.; Elias, F.; Emri, I.; Karn, S.K.; Li, Z.; et al. The rôles of plankton and neuston microbial organic matter in climate regulation. J. Plankton Res. 2021, 43, 801–821. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species; Harvard University Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Dawkins, R. The Extended Selfish Gene; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Biddanda, B.; Dila, D.; Weinke, A.; Mancuso, J.; Villar-Argaiz, M.; Medina-Sánchez, J.M.; González-Olalla, J.M.; Carrillo, P. Housekeeping in the hydrosphere: Microbial cooking, cleaning, and control under stress. Life 2021, 11, 152. [Google Scholar] [CrossRef]
- Chiura, H. Generalized gene transfer by virus-like particles from marine bacteria. Aquat. Microb. Ecol. 1997, 13, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) IPCC Summary for Policy Makers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Rosenwasser, S.; Ziv, C.; van Creveld, S.G.; Vardi, A. Virocell metabolism: Metabolic innovations during host–virus interactions in the ocean. Trends Microbiol. 2016, 24, 821–832. [Google Scholar] [CrossRef]
- Liu, X.; Stock, C.A.; Dunne, J.P.; Lee, M.; Shevliakova, E.; Malyshev, S.; Milly, P.C.D. Simulated global coastal ecosystem responses to a half-century increase in river nitrogen loads. Geophys. Res. Lett. 2021, 48, e2021GL094367. [Google Scholar] [CrossRef]
- Hamilton, D.S.; Perron, M.M.; Bond, T.C.; Bowie, A.R.; Buchholz, R.R.; Guieu, C.; Ito, A.; Maenhaut, W.; Myriokefalitakis, S.; Olgun, N.; et al. Earth, Wind, Fire, and Pollution: Aerosol Nutrient Sources and Impacts on Ocean Biogeochemistry. Annu. Rev. Mar. Sci. 2022, 14, 303–330. [Google Scholar] [CrossRef] [PubMed]
- Jickells, T.D.; Buitenhuis, E.; Altieri, K.; Baker, A.R.; Capone, D.; Duce, R.A.; Dentener, F.; Fennel, K.; Kanakidou, M.; LaRoche, J.; et al. A reevaluation of the magnitude and impacts of anthropogenic atmospheric nitrogen inputs on the ocean. Glob. Biogeochem. Cycles 2017, 31, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Benner, R. Molecular properties are a primary control on the microbial utilization of dissolved organic matter in the ocean. Limnol. Oceanogr. 2019, 65, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Bienfang, P.; Laws, E.; Johnson, W. Phytoplankton sinking rate determination: Technical and theoretical aspects, an improved methodology. J. Exp. Mar. Biol. Ecol. 1977, 30, 283–300. [Google Scholar] [CrossRef]
- Bienfang, P.K. Phytoplankton sinking rates in oligotrophic waters off Hawaii, USA. Mar. Biol. 1980, 61, 69–77. [Google Scholar] [CrossRef]
- Wakeham, S.G.; Lee, C.; Farrington, J.W.; Gagosian, R.B. Biogeochemistry of particulate organic matter in the oceans: Results from sediment trap experiments. Deep. Sea Res. 1984, 31, 509–528. [Google Scholar] [CrossRef]
- Rothstein, J.P. Slip on Superhydrophobic Surfaces. Annu. Rev. Fluid Mech. 2010, 42, 89–109. [Google Scholar] [CrossRef]
- Conlisk, A.T. Essentials of Micro- and Nano-Fluidics; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Jenkinson, I.R.; Sun, J. Drag increase and drag reduction found in phytoplankton and bacterial cultures in laminar flow: Are cell surfaces and EPS producing rheological thickening and a Lotus-leaf Effect? Deep. Sea Res. 2014, 101, 216–230. [Google Scholar] [CrossRef]
- Lim, A.S.; Jeong, H.J.; Jang, T.Y.; Jang, S.H.; Franks, P.J. Inhibition of growth rate and swimming speed of the harmful dinoflagellate Cochlodinium polykrikoides by diatoms: Implications for red tide formation. Harmful Algae 2014, 37, 53–61. [Google Scholar] [CrossRef]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S. In vitro antiviral activities of sulfated polysaccharides from a marine microalga (Cochlodinium polykrikoides) against human immunodeficiency virus and other enveloped viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Gillard, J.; Frenkel, J.; Devos, V.; Sabbe, K.; Paul, C.; Rempt, M.; Inzé, D.; Pohnert, G.; Vuylsteke, M.; Vyverman, W. Metabolomics enables the structure elucidation of a diatom sex pheromone. Angew. Chem. Int. Ed. 2012, 52, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.; Vyverman, W.; Pohnert, G. Pheromone signaling during sexual reproduction in algae. Plant J. 2014, 79, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Tschochner, H.; Lottspeich, F.; Sumper, M. The sexual inducer of Volvox carteri: Purification, chemical characterization and identification of its gene. EMBO J. 1987, 6, 2203–2207. [Google Scholar] [CrossRef]
- Mages, H.-W.; Tschochner, H.; Sumper, M. The sexual inducer of Volvox carteri: Primary structure deduced from cDNA se-quence. FEBS Lett. 1988, 234, 407–410. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.R.; Poulin, R.X.; Mojib, N.; Kubanek, J. Chemical ecology of marine plankton. Nat. Prod. Rep. 2016, 33, 843–860. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Sun, J. Rheological properties of natural waters with regard to plankton thin layers. A short review. J. Mar. Syst. 2010, 83, 287–297. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlusich, J.J.P.; Pelletier, E.; Zinger, L.; Lombard, F.; Zingone, A.; Colin, S.; Gasol, J.M.; Dorrell, R.G.; Henry, N.; Scalco, E.; et al. A robust approach to estimate relative phytoplankton cell abundances from metagenomes. Mol. Ecol. Resour. 2022, 23, 16–40. [Google Scholar] [CrossRef] [PubMed]
- Santschi, P.H.; Chin, W.-C.; Quigg, A.; Xu, C.; Kamalanathan, M.; Lin, P.; Shiu, R.-F. Marine gel interactions with hydrophilic and hydrophobic pollutants. Gels 2021, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Poulson-Ellestad, K.; Mcmillan, E.; Montoya, J.P.; Kubanek, J. Are offshore phytoplankton susceptible to Karenia brevis allelopathy? J. Plankton Res. 2014, 36, 1344–1356. [Google Scholar] [CrossRef] [Green Version]
- Poulson-Ellestad, K.L.; Jones, C.M.; Roy, J.; Viant, M.R.; Fernández, F.M.; Kubanek, J.; Nunn, B.L. Metabolomics and proteomics reveal impacts of chemically mediated competition on marine plankton. Proc. Natl. Acad. Sci. USA 2014, 111, 9009–9014. [Google Scholar] [CrossRef] [Green Version]
- Hakanen, P.; Suikkanen, S.; Kremp, A. Allelopathic activity of the toxic dinoflagellate Alexandrium ostenfeldii: Intra-population variability and response of co-occurring dinoflagellates. Harmful Algae 2014, 39, 287–294. [Google Scholar] [CrossRef]
- Seuront, L.; Stanley, H.E. Anomalous diffusion and multifractality enhance mating encounters in the ocean. Proc. Natl. Acad. Sci. USA 2014, 111, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, F.; Fong, J.; Sadovskaya, I.; Grard, T.; Vinogradov, E. Structural characterization of the extracellular polysaccharide from Vibrio cholerae O1 El-Tor. PLoS ONE 2014, 9, e86751. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Kjelleberg, S.; McDougald, D. Relative contributions of vibrio polysaccharide and quorum sensing to the resistance of Vibrio cholerae to predation by heterotrophic protists. PLoS ONE 2013, 8, e56338. [Google Scholar] [CrossRef] [Green Version]
- Dadon-Pilosof, A.; Conley, K.R.; Jacobi, Y.; Haber, M.; Lombard, F.; Sutherland, K.R.; Steindler, L.; Tikochinski, Y.; Richter, M.; Glöckner, F.O.; et al. Surface properties of SAR11 bacteria facilitate grazing avoidance. Nat. Microbiol. 2017, 2, 1608–1615. [Google Scholar] [CrossRef]
- Rosa, M.; Ward, J.E.; Holohan, B.A.; Shumway, S.E.; Wikfors, G.H. Physicochemical surface properties of microalgae and their combined effects on particle selection by suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol. 2017, 486, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Garcés, E.; Alacid, E.; Reñé, A.; Petrou, K.; Simó, R. Host-released dimethylsulphide activates the dinoflagellate parasitoid Parvilucifera sinerae. ISME J. 2013, 7, 1065–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetacek, V. A watery arms race. Nature 2001, 411, 745. [Google Scholar] [CrossRef]
- Savoca, M.S.; Nevitt, G.A. Evidence that dimethyl sulfide facilitates a tritrophic mutualism between marine primary producers and top predators. Proc. Natl. Acad. Sci. USA 2014, 111, 4157–4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amo, L.; Rodríguez-Gironés, M.; Barbosa, A. Olfactory detection of dimethyl sulphide in a krill-eating Antarctic penguin. Mar. Ecol. Prog. Ser. 2013, 474, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Kerfahi, D.; Harvey, B.P.; Kim, H.; Yang, Y.; Adams, J.M.; Hall-Spencer, J.M. Whole community and functional gene changes of biofilms on marine plastic debris in response to ocean acidification. Microb. Ecol. 2022, 1, 1–13. [Google Scholar] [CrossRef]
- Imai, I.; Inaba, N.; Yamamoto, K. Harmful algal blooms and environmentally friendly control strategies in Japan. Fish. Sci. 2021, 87, 437–464. [Google Scholar] [CrossRef]
- Karn, S.K.; Bhambri, A.; Jenkinson, I.R.; Duan, J.; Kumar, A. The roles of biomolecules in corrosion induction and inhibition of corrosion: A possible insight. Corros. Rev. 2020, 38, 403–421. [Google Scholar] [CrossRef]
- Qin, B.; Deng, J.; Shi, K.; Wang, J.; Brookes, J.; Zhou, J.; Zhang, Y.; Zhu, G.; Paerl, H.W.; Wu, L. extreme climate anomalies enhancing cyanobacterial blooms in eutrophic Lake Taihu, China. Water Resour. Res. 2021, 57, e2020WR029371. [Google Scholar] [CrossRef]
- McManus, M.; Greer, A.; Timmerman, A.; Sevadjian, J.; Woodson, C.; Cowen, R.; Fong, D.; Monismith, S.; Cheriton, O. Characterization of the biological, physical, and chemical properties of a toxic thin layer in a temperate marine system. Mar. Ecol. Prog. Ser. 2021, 678, 17–35. [Google Scholar] [CrossRef]
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734. [Google Scholar] [CrossRef]
- Duan, Z.; Tan, X.; Zeng, Q. Key physiological traits and chemical properties of extracellular polymeric substances determining colony formation in a cyanobacterium. J. Oceanol. Limnol. 2022, 40, 1720–1731. [Google Scholar] [CrossRef]
- Guadayol, Ò.; Mendonca, T.; Segura-Noguera, M.; Wright, A.J.; Tassieri, M.; Humphries, S. Microrheology reveals microscale viscosity gradients in planktonic systems. Proc. Natl. Acad. Sci. USA 2020, 118, e2011389118. [Google Scholar] [CrossRef]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef] [PubMed]
- Lombard, F.; Koski, M.; Kiørboe, T. Copepods use chemical trails to find sinking marine snow aggregates. Limnol. Oceanogr. 2012, 58, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2009, 19, 493–511. [Google Scholar] [CrossRef]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous relations in the arctic marine food web: Interactions between toxin producing Pseudo-nitzschia diatoms and Calanus copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [Green Version]
- Prince, E.K.; Irmer, F.; Pohnert, G. Domoic Acid Improves the Competitive Ability of Pseudo-nitzschia delicatissima against the Diatom Skeletonema marinoi. Mar. Drugs 2013, 11, 2398–2412. [Google Scholar] [CrossRef] [Green Version]
- Hicks, G.R.F.; Grahame, J. Mucus production and its role in the feeding behaviour of Diarthrodes nobilis (Copepoda: Harpacticoida). J. Mar. Biol. Assoc. UK 1979, 59, 321–330. [Google Scholar] [CrossRef]
- Riemann, F.; Schrage, M. The mucus-trap hypothesis on feeding of aquatic nematodes and implications for biodegradation and sediment texture. Oecologia 1978, 34, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.T.; Krock, B.; Hansen, P.J. Production and excretion of okadaic acid, pectenotoxin-2 and a novel dinophysistoxin from the DSP-causing marine dinoflagellate Dinophysis acuta—Effects of light, food availability and growth phase. Harmful Algae 2013, 23, 34–45. [Google Scholar] [CrossRef]
- Ojamäe, K.; Hansen, P.J.; Lips, I. Mass entrapment and lysis of Mesodinium rubrum cells in mucus threads observed in cultures with Dinophysis. Harmful Algae 2016, 55, 77–84. [Google Scholar] [CrossRef]
- Jiang, H.; Kulis, D.M.; Brosnahan, M.L.; Anderson, D.M. Behavioral and mechanistic characteristics of the predator-prey interaction between the dinoflagellate Dinophysis acuminata and the ciliate Mesodinium rubrum. Harmful Algae 2018, 77, 43–54. [Google Scholar] [CrossRef]
- Honsell, G.; Bonifacio, A.; De Bortoli, M.; Penna, A.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Sosa, S.; Yasumoto, T.; et al. New Insights on Cytological and Metabolic Features of Ostreopsis cf. ovata Fukuyo (Dinophyceae): A Multidisciplinary Approach. PLoS ONE 2013, 8, e57291. [Google Scholar] [CrossRef]
- Blossom, H.E.; Daugbjerg, N.; Hansen, P.J. Toxic mucus traps: A novel mechanism that mediates prey uptake in the mixotrophic dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Blossom, H.E.; Hansen, P.J. The loss of mixotrophy in Alexandrium pseudogonyaulax: Implications for trade-offs between toxicity, mucus trap production, and phagotrophy. Limnol. Oceanogr. 2020, 66, 528–542. [Google Scholar] [CrossRef]
- Stehnach, M.R.; Waisbord, N.; Walkama, D.M.; Guasto, J.S. Viscophobic turning dictates microalgae transport in viscosity gradients. Nat. Phys. 2021, 17, 926–930. [Google Scholar] [CrossRef]
- Falciatore, A.; Ribera d’Alcalà, M.; Croot, P.; Bowler, C. Perception of environmental signals by a marine diatom. Science 2000, 288, 2363–2366. [Google Scholar] [CrossRef]
- Wild, C.; Huettel, M.; Klueter, A.; Kremb, S.G.; Rasheed, M.Y.M.; Jørgensen, B.B. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature 2004, 428, 66–70. [Google Scholar] [CrossRef]
- de Goeij, J.M.; van Oevelen, D.; Vermeij, M.J.A.; Osinga, R.; Middelburg, J.J.; de Goeij, A.F.P.M.; Admiraal, W. Surviving in a Marine Desert: The Sponge Loop Retains Resources Within Coral Reefs. Science 2013, 342, 108–110. [Google Scholar] [CrossRef]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Calzetta, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. Multifaceted activity of N-acetyl-l-cysteine in chronic obstructive pulmonary disease. Expert Rev. Respir. Med. 2018, 12, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Z.; Albright, L.J. Anti-phytoplankton therapy of finfish: The mucolytic agent L-cysteine ethyl ester. Dis. Aquat. Org. 1994, 20, 197–202. [Google Scholar] [CrossRef]
- Powell, M.D.; Ransome, J.; Barney, M.; Duijf, R.M.M.; Flik, G. Effect of Dietary Inclusion of N-Acetyl Cysteine on Mucus Viscosity and Susceptibility of Rainbow Trout, Oncorhynchus mykiss, and Atlantic Salmon, Salmo salar, to Amoebic Gill Disease. J. World Aquac. Soc. 2007, 38, 435–442. [Google Scholar] [CrossRef]
- Gutiérrez-Praena, D.; Risalde, M.; Pichardo, S.; Jos, A.; Moyano, R.; Blanco, A.; Vasconcelos, V.; Cameán, A. Histopathological and immunohistochemical analysis of Tilapia (Oreochromis niloticus) exposed to cylindrospermopsin and the effectiveness of N-Acetylcysteine to prevent its toxic effects. Toxicon 2014, 78, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, I.R.; Connors, P.P. The occurrence of the red-tide organism, Gyrodinium aureolum Hulburt (Dinophyceae) around the south and west of Ireland in August and September, 1979. J. Sherkin Isl. 1980, 1, 127–146. [Google Scholar]
- Jenkinson, I.R. Oceanographic implications of non-newtonian properties found in phytoplankton cultures. Nature 1986, 323, 435–437. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Arzul, G. Mitigation by cysteine compounds of rheotoxicity, cytotoxicity and fish mortality caused by the dinoflagellates, Gymnodinium mikimotoi and G. cf. maguelonnense. In Harmful Algal Blooms 2000; Hallegraeff, G., Blackburn, S., Bolch, C., Lewis, R., Eds.; IOC of UNESCO: Paris, France, 2002; pp. 461–464. [Google Scholar]
- Jenkinson, I.R. Increases in viscosity may kill fish in some blooms. In Red Tides; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 435–438. [Google Scholar]
- Jenkinson, I.R.; Arzul, G. Effect of the flagellates, Gymnodinium mikimotoi, Heterosigma akashiwo, and Pavlova lutheri, on flow through fish gills. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M., Wyatt, T., Eds.; Xunta de Galicia: Pontevedra, Spain; IOC of UNESCO: Paris, France, 1998; pp. 425–428. [Google Scholar]
- Gentien, P.; Lunven, M.; Lazure, P.; Youenou, A.; Crassous, M.P. Motility and autotoxicity in Karenia mikimotoi (Dinophyceae). Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1937–1946. [Google Scholar] [CrossRef] [Green Version]
- Orlova, T.Y.; Aleksanin, A.I.; Lepskaya, E.V.; Efimova, K.V.; Selina, M.S.; Morozova, T.V.; Stonik, I.V.; Kachur, V.A.; Karpenko, A.A.; Vinnikov, K.A.; et al. A massive bloom of Karenia species (Dinophyceae) off the Kamchatka coast, Russia, in the fall of 2020. Harmful Algae 2022, 120, 102337. [Google Scholar] [CrossRef]
- Kim, D.; Oda, T. Possible Factors Responsible for the Fish-Killing Mechanisms of the red tide phytoplankton, Chattonella marina and Cochlodinium polykrikoides. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; Terrapub and Nagasaki University: Nagasaki, Japan, 2010; pp. 245–268. [Google Scholar]
- Berdalet, E.; Tester, P.A.; Chinain, M.; Fraga, S.; Lemée, R.; Litaker, W.; Penna, A.; Usup, G.; Vila, M.; Zingone, A. Harmful Algal Blooms in Benthic Systems: Recent Progress and Future Research. Oceanography 2017, 30, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Seuront, L.; Vincent, D. Increased seawater viscosity, Phaeocystis globosa spring bloom and Temora longicornis feeding and swimming behaviours. Mar. Ecol. Prog. Ser. 2008, 363, 131–145. [Google Scholar] [CrossRef]
- Kang, Z.; Yang, B.; Lai, J.; Ning, Y.; Zhong, Q.; Lu, D.; Liao, R.; Wang, P.; Dan, S.F.; She, Z.; et al. Phaeocystis globosa Bloom Monitoring: Based on P. globosa Induced Seawater Viscosity Modification Adjacent to a Nuclear Power Plant in Qinzhou Bay, China. J. Ocean Univ. China 2020, 19, 1207–1220. [Google Scholar] [CrossRef]
- Balkis-Ozdelice, N.; Durmuş, T.; Balci, M. A Preliminary Study on the Intense Pelagic and Benthic Mucilage Phenomenon Observed in the Sea of Marmara. Int. J. Environ. Geoinform. 2021, 8, 414–422. [Google Scholar] [CrossRef]
- Seuront, L.; Leterme, S.C.; Seymour, J.R.; Mitchell, J.G.; Ashcroft, D.; Noble, W.; Thomson, P.G.; Davidson, A.T.; Enden, R.V.D.; Scott, F.J.; et al. Role of microbial and phytoplanktonic communities in the control of seawater viscosity off East Antarctica (30–80° E). Deep. Sea Res. 2010, 57, 877–886. [Google Scholar] [CrossRef]
- Guzmán, H.M.; Cortés, J.; Glynn, P.W.; Richmond, R.H. Coral mortality associated with dinoflagellate blooms in the eastern Pacific. Mar. Ecol. Prog. Ser. 1990, 60, 299–303. [Google Scholar] [CrossRef]
- Kim, D.; Oda, T.; Muramatsu, T.; Kim, D.; Matsuyama, Y.; Honjo, T. Possible factors responsible for the toxicity of Cochlodinium polykrikoides, a red tide phytoplankton. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 132, 415–423. [Google Scholar] [CrossRef]
- Al Gheilani, H.; Al Azri, A.; Piontkovoski, S.; Debrotsov, S.; Al Amri, I.; Al Ambo Ali, I.; Al Jufaili, S.; Al Bousaidi, S.; Al Hajri, S.; Al Aisari, A.; et al. Blooms of Cochlodinium polykrikoides along the coast of Oman and their effect. In Proceedings of the 14th International Conference Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Pagou, K., Hallegraeff, G., Eds.; International Society for the Study of Harmful Algae: Copenhagen, Denmark; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2013; pp. 129–131. [Google Scholar]
- Lee, J.-S. Bioactive components from red tide plankton, Cochlodinium polykrikoides. Korean J. Fish. Aquat. Sci. 1996, 29, 165–173. [Google Scholar]
- Kim, C.S.; Lee, S.G.; Lee, C.K.; Kim, H.G.; Jung, J. Reactive oxygen species as causative agents in the ichthyotoxicity of the red tide dinoflagellate Cochlodinium polykrikoides. J. Plankton Res. 1999, 21, 2105–2115. [Google Scholar] [CrossRef]
- Flores-Leñero, A.; Vargas-Torres, V.; Paredes-Mella, J.; Norambuena, L.; Fuenzalida, G.; Lee-Chang, K.; Mardones, J.I. Heterosigma akashiwo in Patagonian Fjords: Genetics, Growth, Pigment Signature and Role of PUFA and ROS in Ichthyotoxicity. Toxins 2022, 14, 577. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Nagasoe, S.; Tameishi, M.; Shikata, T.; Zou, Y.; Jiang, Z.; Matsubara, T.; Shimasaki, Y.; Yamaguchi, K.; Oshima, Y.; et al. The role of interactions between Prorocentrum minimum and Heterosigma akashiwo in bloom formation. Hydrobiologia 2010, 641, 33–44. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, L.; Sims, I.; Beuzenberg, V.; Gillespie, P. Mass accumulation of mucilage caused by dinoflagellate polysaccharide exudates in Tasman Bay, New Zealand. Harmful Algae 2002, 1, 69–83. [Google Scholar] [CrossRef]
- Honsell, G.; Cabrini, M.; Darin, M. Gonyaulax fragilis (Schütt) Kofoid: A dinoflagellate from gelatinous aggregates of the Northern Adriatic Sea. G. Bot. Ital. 1992, 126, 749–751. [Google Scholar] [CrossRef]
- Pompei, M.; Mazziotti, C.; Guerrini, F.; Cangini, M.; Pigozzi, S.; Benzi, M.; Palamidesi, S.; Boni, L.; Pistocchi, R. Correlation between the presence of Gonyaulax fragilis (Dinophyceae) and the mucilage phenomena of the Emilia-Romagna coast (northern Adriatic Sea). Harmful Algae 2003, 2, 301–316. [Google Scholar] [CrossRef]
- Carbonell-Moore, M.C.; Mertens, K.N. Should Gonyaulax hyalina and Gonyaulax fragilis (Dinophyceae) remain two different taxa? Phycologia 2019, 58, 685–689. [Google Scholar] [CrossRef]
- Riccardi, M.; Guerrini, F.; Serrazanetti, G.P.; Ventrella, V.; Pagliarani, A.; Pistocchi, R. Lipid and DNA features of Gonyaulax fragilis (Dinophyceae) as potential biomarkers in mucilage genesis. Harmful Algae 2010, 9, 359–366. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X. Single molecule mechanochemistry of macromolecules. Prog. Polym. Sci. 2003, 28, 1271–1295. [Google Scholar] [CrossRef]
- Wiita, A.P.; Ainavarapu, S.R.K.; Huang, H.H.; Fernandez, J.M. Force-dependent chemical kinetics of disulfide bond reduction observed with single-molecule techniques. Proc. Natl. Acad. Sci. USA 2006, 103, 7222–7227. [Google Scholar] [CrossRef] [Green Version]
- Alting, A.C.; Hamer, R.J.; de Kruif, C.G.; Paques, M.; Visschers, R.W. Number of thiol groups rather than the size of the aggregates determines the hardness of cold set whey protein gels. Food Hydrocoll. 2003, 17, 469–479. [Google Scholar] [CrossRef]
- Quigg, A.; Santschi, P.; Burd, A.; Chin, W.-C.; Kamalanathan, M.; Xu, C.; Ziervogel, K. From Nano-Gels to Marine Snow: A Synthesis of Gel Formation Processes and Modeling Efforts Involved with Particle Flux in the Ocean. Gels 2021, 7, 114. [Google Scholar] [CrossRef]
- Hu, X.-J.; Xu, Y.; Su, H.-C.; Xu, W.-J.; Wang, L.-H.; Xu, Y.-N.; Li, Z.-J.; Cao, Y.-C.; Wen, G.-L. Algicidal bacterium CZBC1 inhibits the growth of Oscillatoria chlorina, Oscillatoria tenuis, and Oscillatoria planctonica. AMB Express. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-O.; Kato, J.; Takiguchi, N.; Kuroda, A.; Ikeda, T.; Mitsutani, A.; Ohtake, H. Involvement of an Extracellular Protease in Algicidal Activity of the Marine Bacterium Pseudoalteromonas sp. Strain A28. Appl. Environ. Microbiol. 2000, 66, 4334–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Yang, Y.; Zhu, X.-Y.; Zhang, X.-H.; Yu, M. Metagenomic Insights Into the Microbial Assemblage Capable of Quorum Sensing and Quorum Quenching in Particulate Organic Matter in the Yellow Sea. Front. Microbiol. 2021, 11, 602010. [Google Scholar] [CrossRef] [PubMed]
- Svedrup, H.U.; Johnson, M.W.; Fleming, R.H. The Oceans: Their Physics, Chemistry, and General Biology, 1st ed.; Prentice Hall: New York, NY, USA, 1942. [Google Scholar]
- Endo, M. Calcium Ion as a Second Messenger With Special Reference to Excitation-Contraction Coupling. J. Pharmacol. Sci. 2006, 100, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Ianora, A.; Boersma, M.; Casotti, R.; Fontana, A.; Harder, J.; Hoffmann, F.; Pavia, H.; Potin, P.; Poulet, S.A.; Toth, G. New trends in marine chemical ecology. Estuaries Coasts 2006, 29, 531–551. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Wyatt, T. Selection and control of Deborah numbers in plankton ecology. J. Plankton Res. 1992, 14, 1697–1721. [Google Scholar] [CrossRef]
- Seuront, L.; Lacheze, C.; Doubell, M.J.; Seymour, J.R.; Van Dongen-Vogels, V.; Newton, K.; Alderkamp, A.C.; Mitchell, J.G. The influence of Phaeocystis globosa on microscale spatial patterns of chlorophyll a and bulk-phase seawater viscosity. Biodegradation 2007, 83, 173–188. [Google Scholar] [CrossRef]
- Wyatt, T.; Ribera d’Alcalà, M. Dissolved organic matter and planktonic engineerin. CIESM Workshop Monogrs. 2006, 28, 13–23. [Google Scholar]
- Hastings, A.; Byers, J.; Crooks, J.A.; Cuddington, K.; Jones, C.G.; Lambrinos, J.G.; Talley, T.S.; Wilson, W.G. Ecosystem engineering in space and time. Ecol. Lett. 2007, 10, 153–164. [Google Scholar] [CrossRef]
- Reddington, K.; Eccles, D.; O’Grady, J.; Drown, D.M.; Hansen, L.H.; Nielsen, T.K.; Ducluzeau, A.-L.; Leggett, R.M.; Heavens, D.; Peel, N.; et al. Metagenomic analysis of planktonic riverine microbial consortia using nanopore sequencing reveals insight into river microbe taxonomy and function. Gigascience 2020, 9, giaa053. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Wyatt, T. Management by phytoplankton of physical oceanographic parameters. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier Intercept: Paris, France, 1995; pp. 603–607. [Google Scholar]
- Camacho-Chab, J.C.; Lango-Reynoso, F.; Castañeda-Chávez, M.D.R.; Galaviz-Villa, I.; Hinojosa-Garro, D.; Ortega-Morales, B.O. Implications of Extracellular Polymeric Substance Matrices of Microbial Habitats Associated with Coastal Aquaculture Systems. Water 2016, 8, 369. [Google Scholar] [CrossRef]
- Sretenovic, S.; Stojković, B.; Dogsa, I.; Kostanjšek, R.; Poberaj, I.; Stopar, D. An early mechanical coupling of planktonic bacteria in dilute suspensions. Nat. Commun. 2017, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Jatt, A.N.; Tang, K.; Liu, J.; Zhang, Z.; Zhang, X.-H. Quorum sensing in marine snow and its possible influence on production of extracellular hydrolytic enzymes in marine snow bacterium Pantoea ananatis B9. FEMS Microbiol. Ecol. 2014, 91, 1–13. [Google Scholar] [CrossRef]
- Decho, A.W.; Gutierrez, T. Microbial Extracellular Polymeric Substances (EPSs) in Ocean Systems. Front. Microbiol. 2017, 8, 922. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Tay, Q.X.M.; Kjelleberg, S.; Rice, S.A.; McDougald, D. Quorum sensing-regulated chitin metabolism provides grazing resistance to Vibrio cholerae biofilms. ISME J. 2015, 9, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Svenningsen, S.L.; Middelboe, M. Quorum Sensing Determines the Choice of Antiphage Defense Strategy in Vibrio anguillarum. Mbio 2015, 6, e00627-15. [Google Scholar] [CrossRef] [Green Version]
- Pappas, K.M.; Weingart, C.L.; Winans, S.C. Chemical communication in proteobacteria: Biochemical and structural studies of signal synthases and receptors required for intercellular signalling. Mol. Microbiol. 2004, 53, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Grossart, H.-P.; Schlingloff, A.; Kiørboe, T. Possible Quorum Sensing in Marine Snow Bacteria: Production of Acylated Homoserine Lactones by Roseobacter Strains Isolated from Marine Snow. Appl. Environ. Microbiol. 2002, 68, 4111–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsek, M.R.; Fuqua, C.; Bordi, C.; Ansaldi, M.; Gon, S.; Jourlin-Castelli, C.; Iobbi-Nivol, C.; Méjean, V. Biofilms 2003: Emerging Themes and Challenges in Studies of Surface-Associated Microbial Life. J. Bacteriol. 2004, 186, 4502–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, K.; Joint, I.; Daykin, M.; Milton, D.L.; Williams, P.; Camara, M. Disruption of quorum sensing in seawater abolishes attraction of zoospores of the green alga Ulva to bacterial biofilms. Environ. Microbiol. 2004, 7, 229–240. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Tait, K.; Taylor, A.; Brownlee, C.; Joint, I. Acyl-homoserine lactones modulate the settlement rate of zoospores of the marine alga Ulva intestinalis via a novel chemokinetic mechanism. Plant Cell Environ. 2006, 29, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, J.; Lan, C.; Lai, J.; Wang, J.; Wei, F. Inhibitory effect of increase in water viscosity of Phaeocystis globosa on Feeding of Temora stylifera. Guangxi Sci. 2021, 28, 56–64, (In Chinese, English abstract). [Google Scholar]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Hohne, D.N.; Younger, J.G.; Solomon, M.J. Flexible Microfluidic Device for Mechanical Property Characterization of Soft Viscoelastic Solids Such as Bacterial Biofilms. Langmuir 2009, 25, 7743–7751. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Chin, W.-C.; Chiu, M.-H.; Xu, C.; Lin, P.; Schwehr, K.A.; Quigg, A.; Santschi, P.H. Sunlight induced aggregation of dissolved organic matter: Role of proteins in linking organic carbon and nitrogen cycling in seawater. Sci. Total. Environ. 2018, 654, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, N.; Dai, X.; Tao, W.; Jenkinson, I.R.; Li, Z. Enhanced dewaterability of sludge during anaerobic digestion with thermal hydrolysis pretreatment: New insights through structure evolution. Water Res. 2018, 131, 177–185. [Google Scholar] [CrossRef]
- Lear, G. (Ed.) Biofilms in Bioremediation; Caister Academic Press: Poole, UK, 2016; Available online: https://www.lulu.com/spotlight/caister (accessed on 15 February 2023).
- Verdugo, P. Marine Microgels. Annu. Rev. Mar. Sci. 2012, 4, 375–400. [Google Scholar] [CrossRef]
- Verdugo, P. Marine Biopolymer Dynamics, Gel Formation, and Carbon Cycling in the Ocean. Gels 2021, 7, 136. [Google Scholar] [CrossRef]
- Drost-Hansen, W. Vicinal hydration of biopolymers: Cell biological consequences. In Water and the Cell; Pollack, G.H., Cameron, I.L., Wheatley, D.N., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 175–217. [Google Scholar]
- Mari, X. Does ocean acidification induce an upward flux of marine aggregates? Biogeosciences 2008, 5, 1023–1031. [Google Scholar] [CrossRef] [Green Version]
- Bartual, A.; Cera, I.V.; Flecha, S.; Prieto, L. Effect of dissolved polyunsaturated aldehydes on the size distribution of transparent exopolymeric particles in an experimental diatom bloom. Mar. Biol. 2017, 164, 120. [Google Scholar] [CrossRef]
- Klun, K.; Šket, P.; Beran, A.; Falnoga, I.; Faganeli, J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water 2022, 15, 111. [Google Scholar] [CrossRef]
- Vidal-Melgosa, S.; Sichert, A.; Ben Francis, T.; Bartosik, D.; Niggemann, J.; Wichels, A.; Willats, W.G.T.; Fuchs, B.M.; Teeling, H.; Becher, D.; et al. Diatom fucan polysaccharide precipitates carbon during algal blooms. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.; Lefevre, D.; Thyssen, M.; Jenkinson, I.R.; Grégori, G. Pulsed export of carbon in the north-western Mediterranean Sea. J. Oceanol. Limnol. 2023, 41, 189–202. [Google Scholar] [CrossRef]
- Liu, L.; Qin, B.; Zhu, G.; Zhang, Y.; Gao, G.; Gong, Z.; Huang, Q. Distribution of dissolved acidic polysaccharides (dAPS) during cyanobacteria blooms in northern Lake Taihu. Limnology 2014, 16, 21–29. [Google Scholar] [CrossRef]
- Laabir, M.; Grignon-Dubois, M.; Masseret, E.; Rezzonico, B.; Soteras, G.; Rouquette, M.; Rieuvilleneuve, F.; Cecchi, P. Algicidal effects of Zostera marina L. and Zostera noltii Hornem. extracts on the neuro-toxic bloom-forming dinoflagellate Alexandrium catenella. Aquat. Bot. 2013, 111, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Onishi, Y.; Mohri, Y.; Tuji, A.; Ohgi, K.; Yamaguchi, A.; Imai, I. The seagrass Zostera marina harbors growth-inhibiting bacteria against the toxic dinoflagellate Alexandrium tamarense. Fish. Sci. 2014, 80, 353–362. [Google Scholar] [CrossRef]
- Imai, I.; Kido, T.; Yoshinaga, I.; Ohgi, K.; Nagai, S. Isolation of Microcystis-killer bacterium Agrobacterium vitis from the biofilm onthe surface of the water plant Egeria densa. In Proceedings of the 14th International Conference Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Pagou, K.A., Hallegraeff, G.M., Eds.; ISSHA: Copenhagen, Denmark; IOC of UNESCO: Paris, France, 2013; pp. 150–152. [Google Scholar]
- Yang, Y.; Hu, X.; Zhang, J.; Gong, Y. Community level physiological study of algicidal bacteria in the phycospheres of Skeletonema costatum and Scrippsiella trochoidea. Harmful Algae 2013, 28, 88–96. [Google Scholar] [CrossRef]
- Qian, X.; Chen, L.; Sui, Y.; Chen, C.; Zhang, W.; Zhou, J.; Dong, W.; Jiang, M.; Xin, F.; Ochsenreither, K. Biotechnological potential and applications of microbial consortia. Biotechnol. Adv. 2020, 40, 107500. [Google Scholar] [CrossRef]
- Kotay, S.M.; Das, D. Microbial hydrogen production from sewage sludge bioaugmented with a constructed microbial consortium. Int. J. Hydrog. Energy 2010, 35, 10653–10659. [Google Scholar] [CrossRef]
- Thakur, P.; Alaba, M.O.; Rauniyar, S.; Singh, R.N.; Saxena, P.; Bomgni, A.; Gnimpieba, E.Z.; Lushbough, C.; Goh, K.M.; Sani, R.K. Text-Mining to Identify Gene Sets Involved in Biocorrosion by Sulfate-Reducing Bacteria: A Semi-Automated Workflow. Microorganisms 2023, 11, 119. [Google Scholar] [CrossRef]
- Singh, A. Glycoproteomics. Nat. Methods 2021, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Alldredge, A.L.; Passow, U.; Logan, B.E. The abundance and significance of a class of large, transparent organic particles in the ocean. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 1131–1140. [Google Scholar] [CrossRef]
- Williams, P.; Carlucci, A.; Henrichs, S.; Van Vleet, E.; Horrigan, S.; Reid, F.; Robertson, K. Chemical and microbiological studies of sea-surface films in the Southern Gulf of California and off the West Coast of Baja California. Mar. Chem. 1986, 19, 17–98. [Google Scholar] [CrossRef]
- Carlson, D.J. Viscosity of sea-surface slicks. Nature 1987, 329, 823–825. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, L.; Liu, C.; Cai, W. Viscosity of sea surface microlayer in Jiaozhou Bay and adjacent sea area. Chin. J. Oceanol. Limnol. 2003, 21, 351–357. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Seuront, L.; Ding, H.; Elias, F. Biological modification of mechanical properties of the sea surface microlayer, influencing waves, ripples, foam and air-sea fluxes. Elem. Sci. Anthr. 2018, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Miles, J.; Lovelock, J. The Ages of Gaia. A Biography of Our Living Earth. J. Appl. Ecol. 1989, 26, 736. [Google Scholar] [CrossRef]
- Lenton, T.M. Gaia and natural selection. Nature 1998, 394, 439–447. [Google Scholar] [CrossRef]
- Thornton, D.C. Phytoplankton mucilage production in coastal waters: A dispersal mechanism in a front dominated system? Ethol. Ecol. Evol. 1999, 11, 179–185. [Google Scholar] [CrossRef]
- Hamilton, W.; Lenton, T. Spora and Gaia: How microbes fly with their clouds. Ethol. Ecol. Evol. 1998, 10, 1–16. [Google Scholar] [CrossRef]
- Teeling, H.; Fuchs, B.M.; Becher, D.; Klockow, C.; Gardebrecht, A.; Bennke, C.M.; Kassabgy, M.; Huang, S.; Mann, A.J.; Waldmann, J.; et al. Substrate-Controlled Succession of Marine Bacterioplankton Populations Induced by a Phytoplankton Bloom. Science 2012, 336, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Claustre, H.; Levy, M.; Siegel, D.A.; Weber, T. Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature 2019, 568, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Gregor, L.; Hauck, J.; Le Quéré, C.; Luijkx, I.T.; Olsen, A.; Peters, G.P.; et al. Global Carbon Budget 2022. Earth Syst. Sci. Data 2022, 14, 4811–4900. [Google Scholar] [CrossRef]
- Gulev, S.K.; Thorne, P.W. Chapter 2: Changing state of the climate system. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; pp. 287–422. [Google Scholar]
- Stabeno, P.J.; Monahan, E.C. The Influence of whitecaps on the albedo of the sea surface. In Oceanographic Science Library; Monahan, E.C., Mac Niocaill, G., Eds.; D. Reidel: Dordrecht, Germany; Galway University Press: Galway, Ireland, 1986; pp. 261–266. [Google Scholar]
- Evans, J.R.G.; Stride, E.P.J.; Edirisinghe, M.J.; Andrews, D.J.; Simon, R.R. Can oceanic foams limit global warming? Clim. Res. 2010, 42, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Cantat, I.; Cohen-Addad, S.; Elias, F.; Graner, F.; Höhler, R.; Pitois, O.; Rouyer, F.; Saint-Jalmes, A.; Cox, S. (Eds.) Foams; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Callaghan, A.H.; Deane, G.B.; Stokes, M.D.; Ward, B. Observed variation in the decay time of oceanic whitecap foam. J. Geophys. Res. Atmos. 2012, 117, C09015. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Blum, P.C.; Lyons, J.; Cheng, Y.S.; Yazzie, D.; Zhou, Y. Brevetoxin Concentrations in Marine Aerosol: Human Exposure Levels During a Karenia brevis Harmful Algal Bloom. Bull. Environ. Contam. Toxicol. 2003, 70, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Montero, M.; Freer, E.; Jiménez-Montealegre, R.; Guzmán, J. Occurrence and predominance of the fish killer Cochlod-inium polykrikoides on the Pacific coast of Costa Rica. Afr. J. Mar. Sci. 2006, 28, 215–217. [Google Scholar] [CrossRef]
- Kesaulya, I.; Leterme, S.C.; Mitchell, J.G.; Seuront, L. The impact of turbulence and phytoplankton dynamics on foam formation, seawater viscosity and chlorophyll concentration in the eastern English Channel. Oceanologia 2008, 50, 167–182. [Google Scholar]
- Calvo, E.; Viquez, R.; Garcia, A. Alexandrium monilatum (Howell) Balech bloom in the Gulf of Nicoya, Puntarenas. Harmful Algae News 2005, 29, 1–2. [Google Scholar]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and Harmful Microalgae of the World Ocean; ISSHA: Copenhagen, Denmark; IOC of UNESCO: Paris, France, 2016. [Google Scholar]


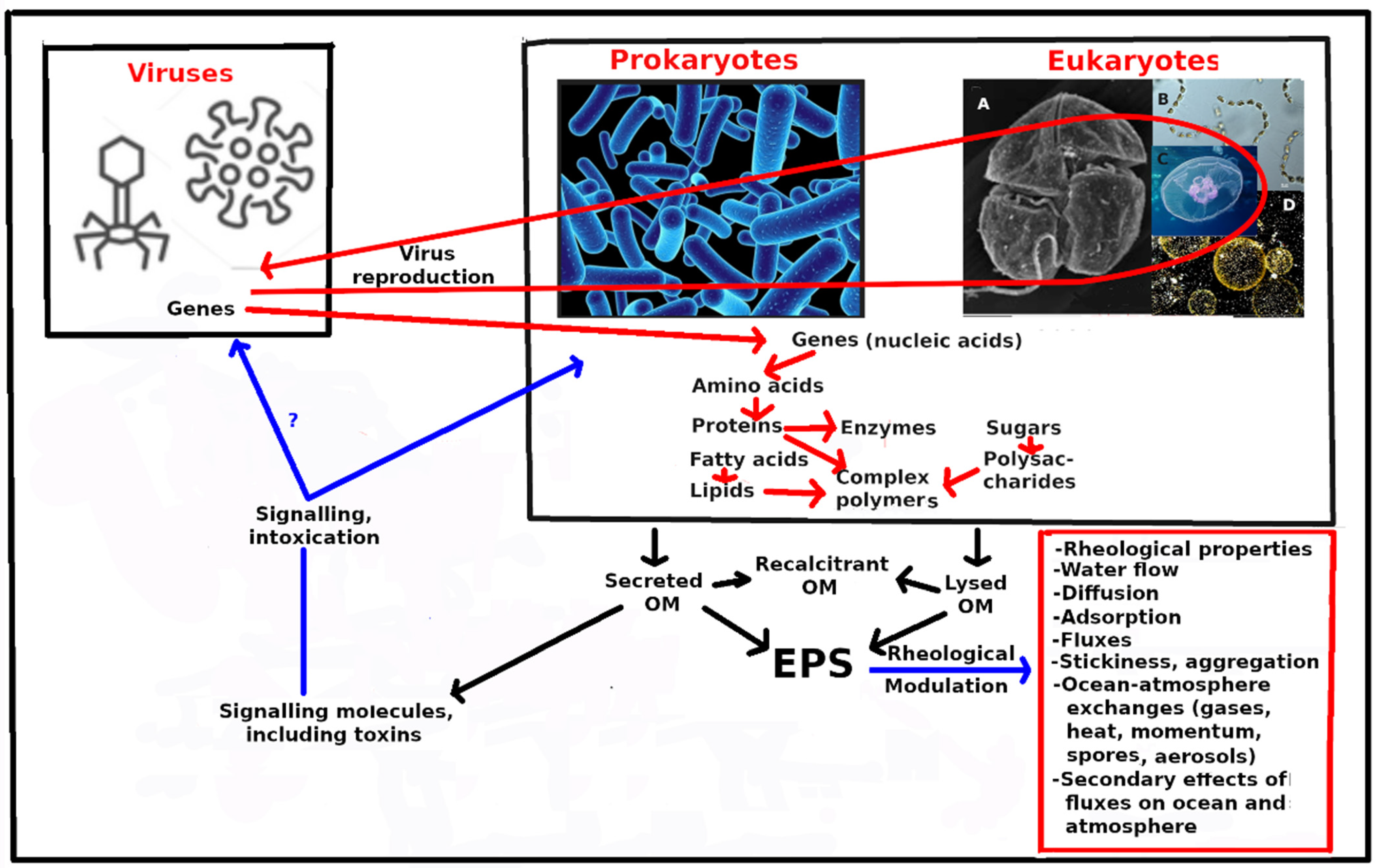{kind=link}
| Coastal Ocean Ref. [17] | Whole Ocean Refs. [18,19] | Open Ocean Ref. [19] | |
|---|---|---|---|
| Inorganic N (IN) | |||
| Atmospheric deposition (DIN) | +4.5 | ||
| River input (DIN) | +20.4 | +23 [19] | +17 (a) |
| Denitrification (water + sediments) (DIN) | −51.9 | ||
| Coast-ward net influx from offshore (DIN) | +47.4 | ||
| Total ΔTIN | +20.4 | ||
| Organic and inorganic N | |||
| Sedimentary burial (TON) | −12.3 | ||
| River input (TON) | +27.1 | ||
| River input (DON) | +11 (a) [19] | >0 to <11 (a) | |
| N2 fixation (TON) | +15.4 | 164 [19] | 0 |
| Oceanward net outflux to offshore (TON) | −50.2 | ||
| Total Δ TON | −20.0 | ||
| Discrepancy (ΔTIN − ΔTON) | +0.4 | ||
| Net community production (DIN + TON) | 35.5 | ||
| Atmospheric N deposition | 39 (b) [19] | >30 (b) | |
| Atmosphere-ocean Fe budget | |||
| Fe emissions from fires (<20 µm) | 1.1 [18] | ||
| Soluble Fe flux to the ocean (from dust) | 0.19 to 0.28 [18] | ||
| Soluble Fe flux to the ocean (from fires) | 0.035 to 0.063 [18] | ||
| Soluble Fe flux to the ocean (anthropogenic) | 0.016 to 0.034 [18] | ||
| Soluble Fe flux to the ocean (Total) | 0.24 to 0.38 [18] | ||
| Atmosphere-ocean P budget Soluble P flux to the ocean (from dust) | 0.031 to 0.094 [18] | ||
| Soluble P flux to the ocean (from fires) | 0.005 [18] | ||
| Soluble P flux to the ocean (anthropogenic) | 0.0094 to 0.11 [18] | ||
| Soluble P flux to the ocean (Total) | 0.045 to 0.21 [18] |
| Lipids | Proteins and CRAM | Polysaccharides | Formate | Corg. | % COC * | |
|---|---|---|---|---|---|---|
| HCH2-CH2- | HC-HCOR | HC-OH HC-O-C | HCOO | |||
| δ **/ppm | 0–1.8 | 1.8–3.0 | 3.0–4.6 | 8.0–9.0 | µmol L−1 | % |
| Tetraselmis sp. | ||||||
| 0.2 µm filtrate | 915.1 | |||||
| Retentate | 19.4 | 18.8 | 61.4 | 0.4 | 364 | 39.8 |
| Permeate | 35.7 | 40.9 | 20.8 | 2.6 | 507 | |
| Chaetoceros socialis | ||||||
| 0.2 µm filtrate | 2285 | |||||
| Retentate | 14.4 | 21.9 | 62.9 | 1.1 | 526 | 23.0 |
| Permeate | 16.4 | 36.4 | 45.5 | 1.7 | 1765 | |
| Prorocentrum minimum | ||||||
| 0.2 µm filtrate | 439.3 | |||||
| Retentate | 21.8 | 45.6 | 31.9 | 0.8 | 154 | 35.1 |
| Permeate | 17.4 | 56.7 | 24.6 | 1.2 | 418 | |
| Mechanism | Global Magnitude (PgCyr−1) | Depth Horizon | Remarks |
|---|---|---|---|
| Downward transport from the photic zone into the ocean interior. (Recompiled from Boyd et al. [167] and references therein.) | |||
| Biological gravitational pump | 4.9 to 9.1 | 100 m or euphotic zone. | |
| Mesopelagic migrant pump | 0.9 to 3.6 | ||
| Seasonal lipid pump | 0.25 to 1.0 | ||
| Mixed layer pump | 0.1 to 1.0 | Mixed layer. | |
| Large-scale subduction pump | 0.13 | ||
| Eddy subduction pump | 2.0 | 100 m. | Low confidence of precision |
| Sum of physical pumps | 1.8 to 2.1 | Mixed layer, euphotic zone or 74 m. | Low confidence of precision |
| Aquatic input of anthropogenic OC from the land, and from total primary production in situ. | |||
| Input of land-derived OC to the open ocean through the land-ocean aquatic continuum (LOAC) | 0.65 | Probably mostly input to photic zone, but an unknown fraction is rapidly subducted to below the photic zone. | Low confidence of precision [168] |
| Global marine primary productivity | 47 (±8) | IPCC [169] N.B. This estimate includes coastal waters. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenkinson, I.R. Plankton Genes and Extracellular Organic Substances in the Ocean. J. Mar. Sci. Eng. 2023, 11, 783. https://doi.org/10.3390/jmse11040783
Jenkinson IR. Plankton Genes and Extracellular Organic Substances in the Ocean. Journal of Marine Science and Engineering. 2023; 11(4):783. https://doi.org/10.3390/jmse11040783
Chicago/Turabian StyleJenkinson, Ian R. 2023. "Plankton Genes and Extracellular Organic Substances in the Ocean" Journal of Marine Science and Engineering 11, no. 4: 783. https://doi.org/10.3390/jmse11040783
APA StyleJenkinson, I. R. (2023). Plankton Genes and Extracellular Organic Substances in the Ocean. Journal of Marine Science and Engineering, 11(4), 783. https://doi.org/10.3390/jmse11040783