Posidonia oceanica as a Source of Chromophoric Dissolved Organic Matter for the Oligotrophic NW Mediterranean Coast

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
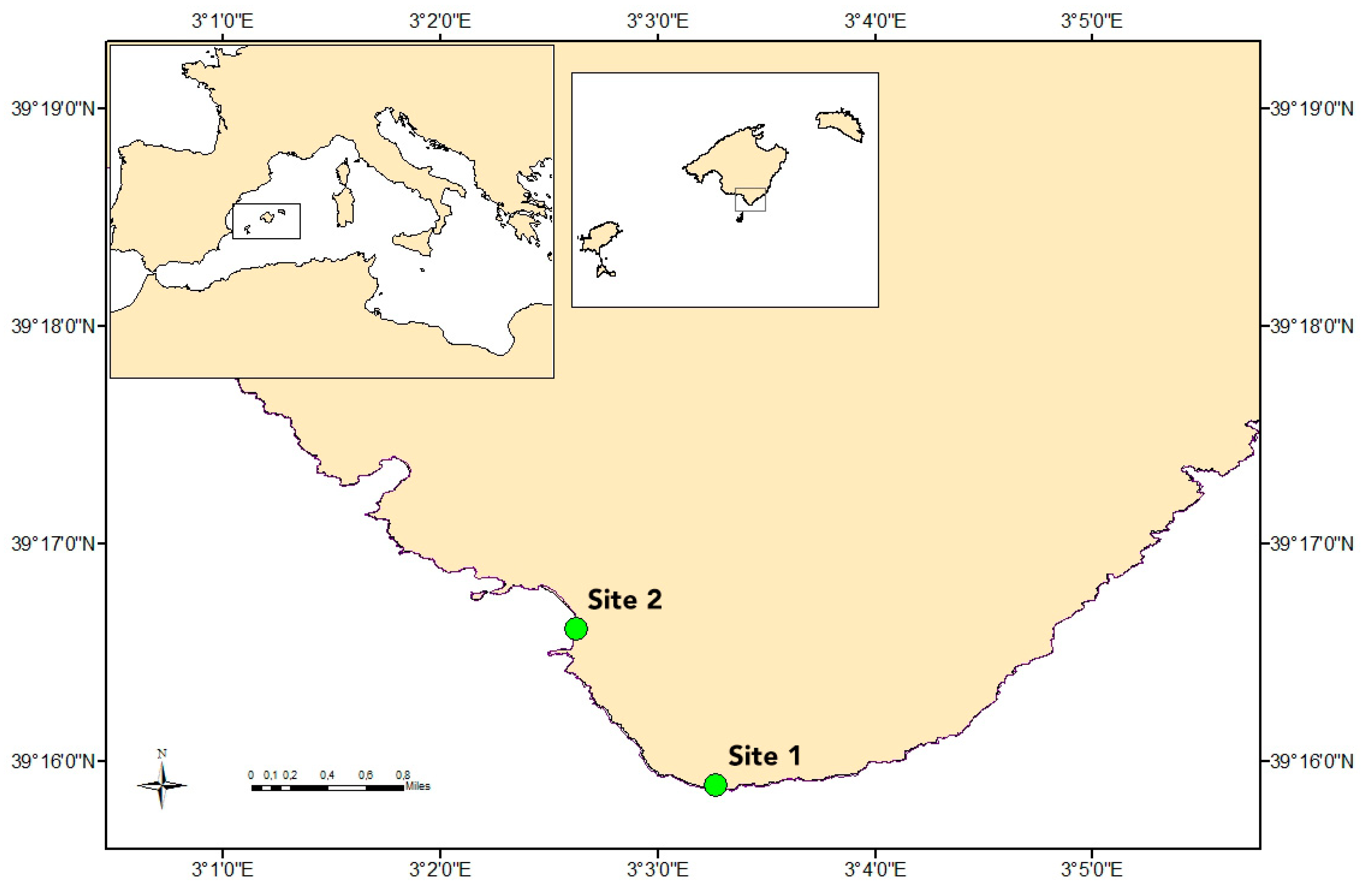
2.1. Sampling Strategy
2.2. Physical and Biogeochemical Variables
2.3. CDOM Measurements
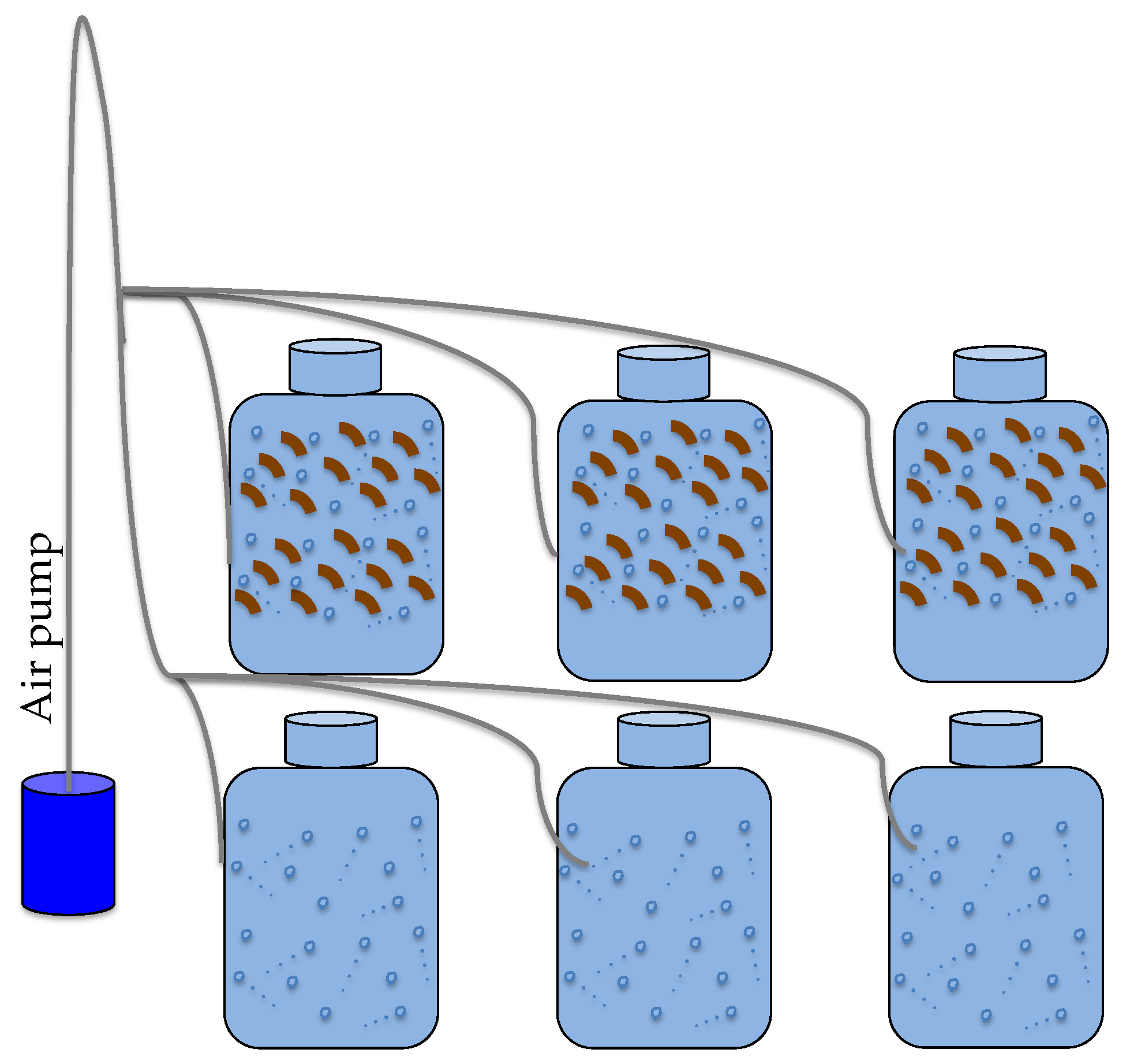
2.4. Experimental Assessment of CDOM Release by P. oceanica Leaf Litter
2.5. Statistical Analysis
3. Results
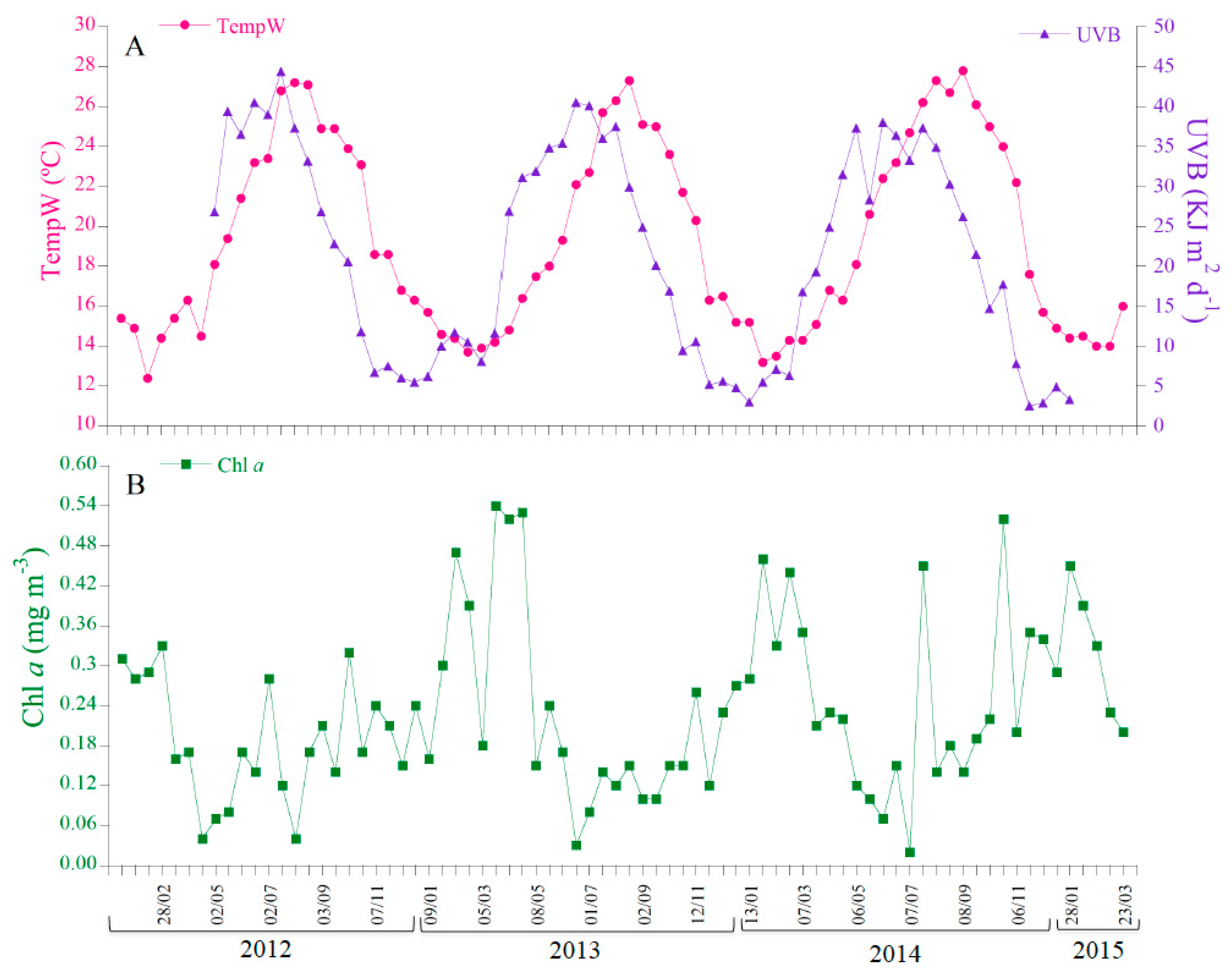
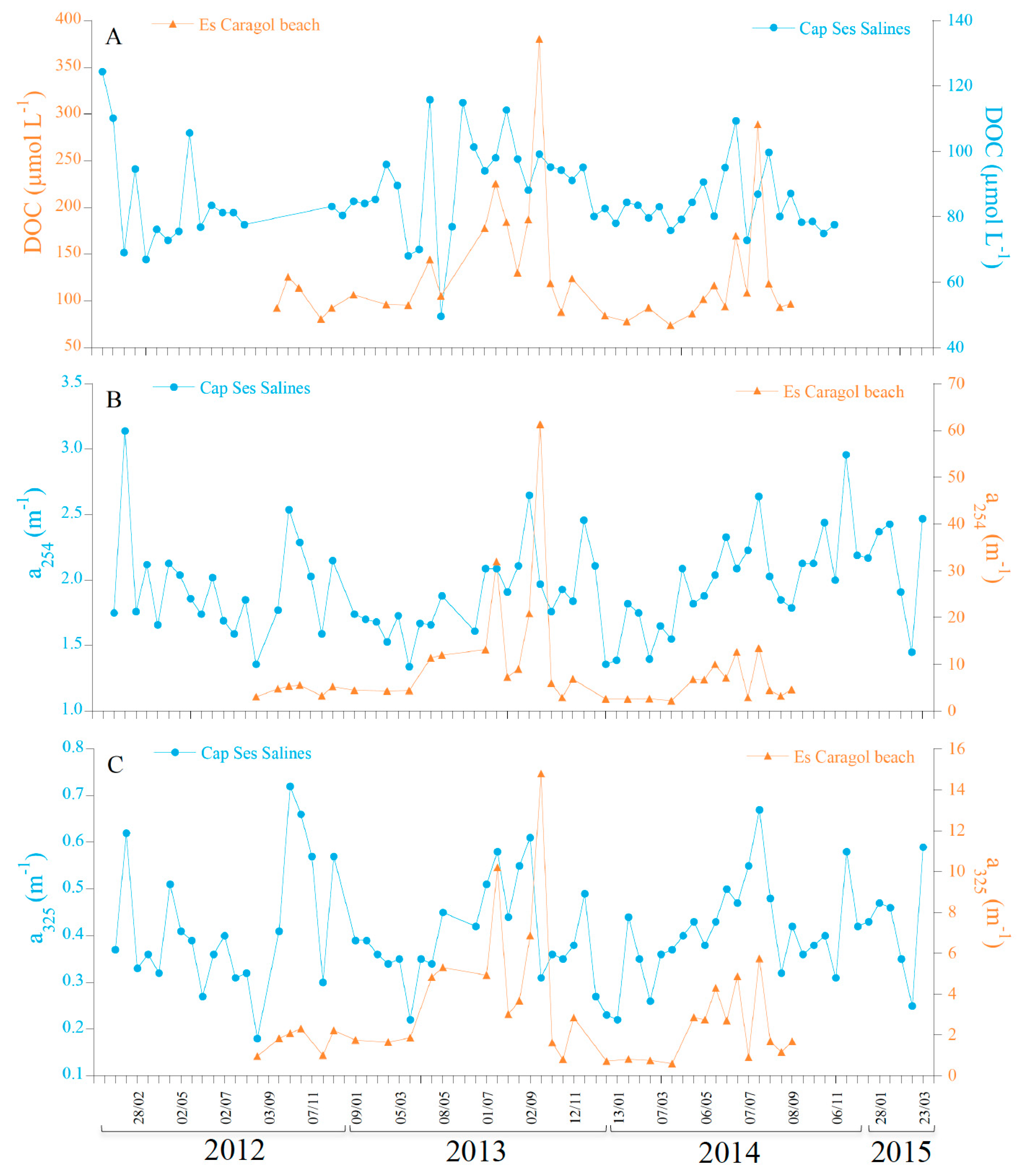
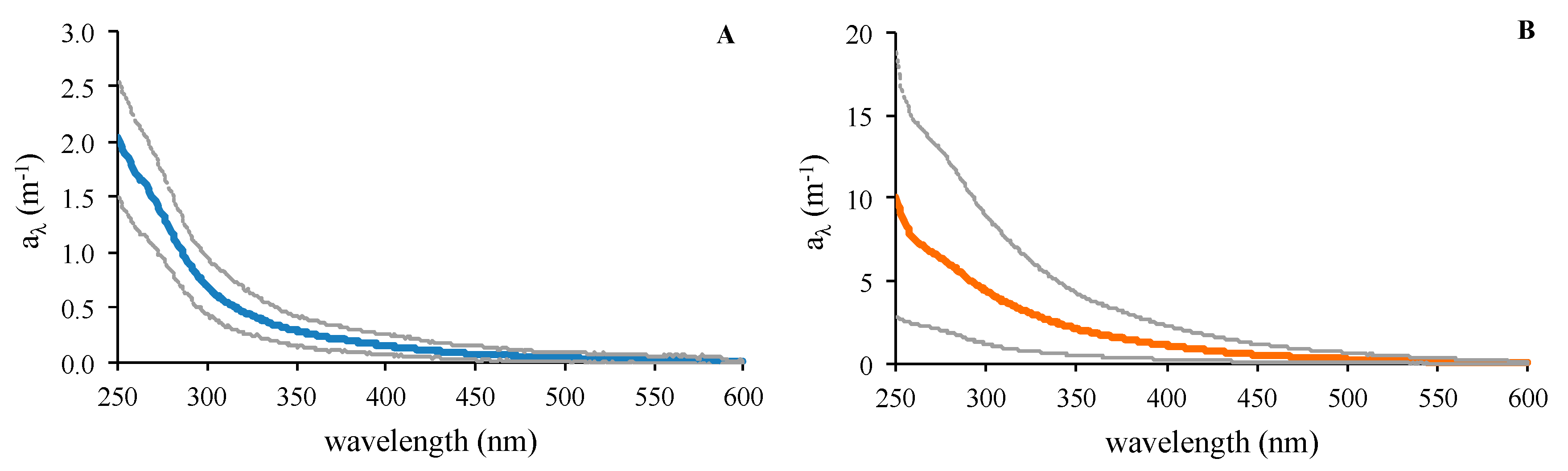
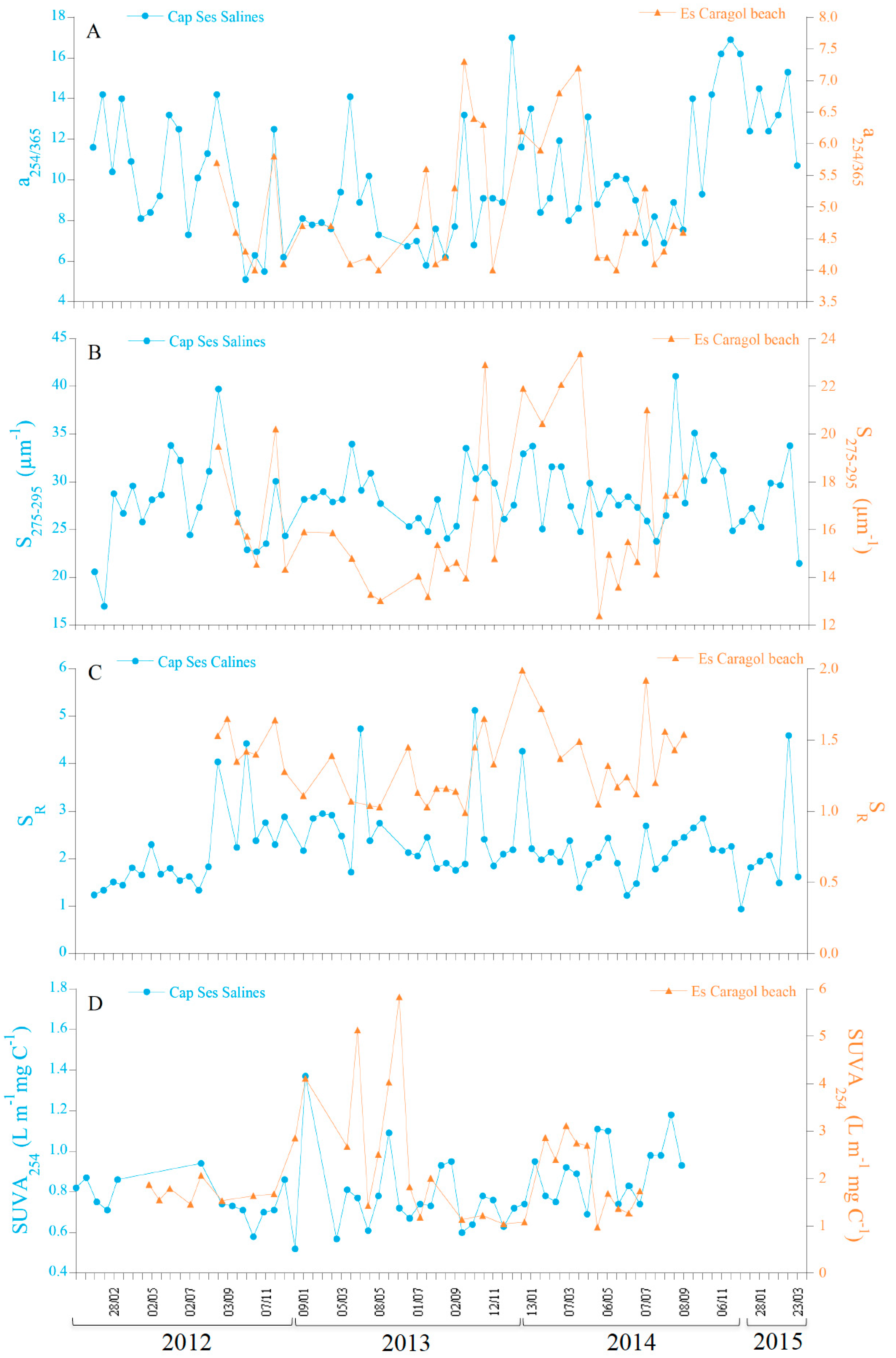
3.1. Time-Series at Cap Ses Salines and Es Caragol Beach
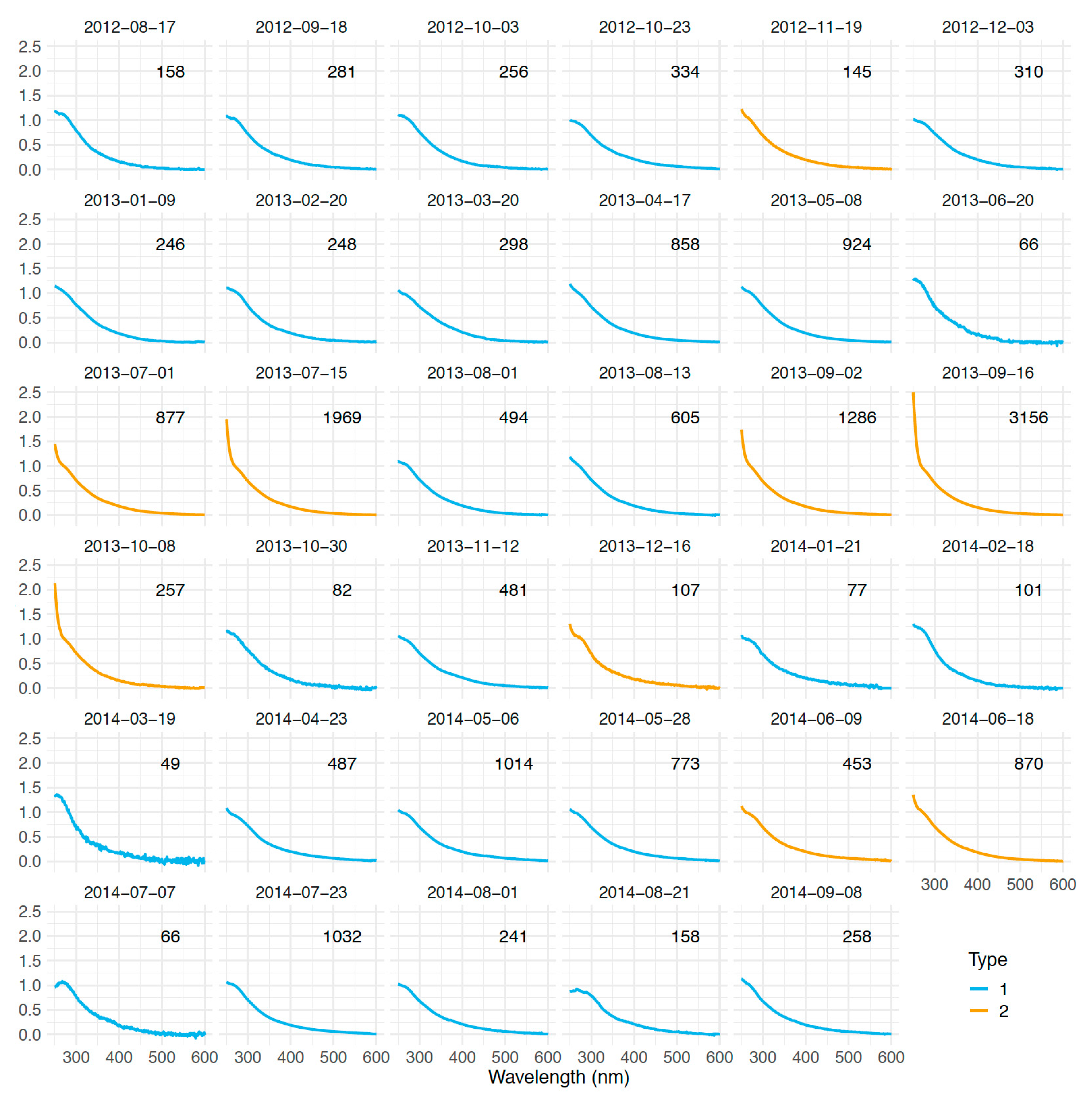
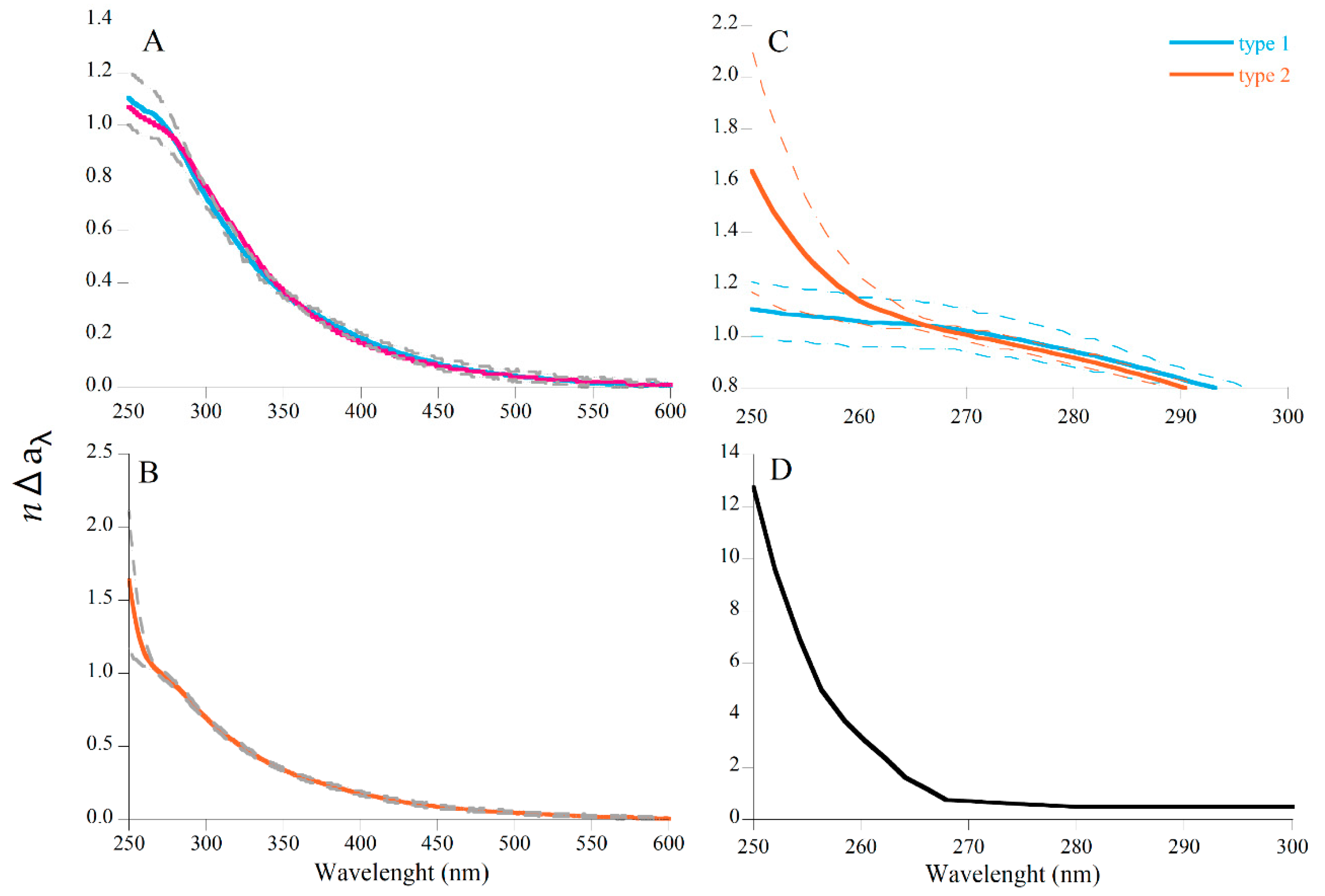
3.2. Comparison of In Situ and Laboratory P. oceanica Leaf Litter Absorption Spectra
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morel, A.; Maritorena, S. Bio-optical properties of oceanic waters: A reappraisal. J. Geophys. Res. 2001, 106, 7163–7180. [Google Scholar] [CrossRef] [Green Version]
- Coble, P.G. Marine optical biogeochemistry: The chemistry of ocean color. Chem. Rev. 2007, 107, 402–418. [Google Scholar] [CrossRef] [PubMed]
- Mopper, K.; Kieber, D.J.; Stubbins, A. Marine Photochemistry of Organic Matter: Processes and Impacts. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 389–450. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Siegel, D.A.; Maritorena, S.; Nelson, N.B. Global distribution and dynamics of colored dissolved and detrital organic materials. J. Geophys. Res. 2002, 107. [Google Scholar] [CrossRef]
- Zepp, R.G.; Erickson, D.J.; Paul, N.D.; Sulzberger, B. Interactive effects of solar UV radiation and climate change on biogeochemical cycling. Photochem. Photobiol. Sci. 2007, 6, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Jaffé, R.; McKnight, D.; Maie, N.; Cory, R.; McDowell, W.H.; Campbell, J.L. Spatial and temporal variations in DOM composition in ecosystems: The importance of long-term monitoring of optical properties. J. Geophys. Res. Biogeosci. 2008, 113, G04032. [Google Scholar] [CrossRef]
- Cawley, K.M.; Ding, Y.; Fourqurean, J.; Jaffé, R. Characterising the sources and fate of dissolved organic matter in Shark Bay, Australia: A preliminary study using optical properties and stable carbon isotopes. Mar. Freshw. Res. 2012, 63, 1098–1107. [Google Scholar] [CrossRef] [Green Version]
- Stedmon, C.A.; Nelson, N.B. The Optical Properties of DOM in the Ocean. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 481–508. [Google Scholar] [CrossRef]
- Pain, A.J.; Martin, J.B.; Young, C.R.; Huang, L.; Valle-Levinson, A. Organic matter quantity and quality across salinity gradients in conduit- vs. diffuse flow-dominated subterranean estuaries. Limnol. Oceanogr. 2019, 64, 1386–1402. [Google Scholar] [CrossRef] [Green Version]
- Coble, P.G.; Chuanmin, H.; Gould, R.W.; Chang, G.; Wood, A.M. Colored Dissolved Organic in the Coastal Ocean: An optical tool for coastal zone environmental assessment and management. Oceanogr. Coast. Ocean Opt. Dyn. 2004, 17, 50–59. [Google Scholar]
- Romera-Castillo, C.; Sarmento, H.; Álvarez-Salgado, X.A.; Gasol, J.M.; Marrasé, C. Production of chromophoric dissolved organic matter by marine phytoplankton. Limnol. Oceanogr. 2010, 55, 446–454. [Google Scholar] [CrossRef]
- Chari, N.V.H.K.; Keerthi, S.; Sarma, N.S.; Pandi, R.S.; Chiranjeevulu, G.; Kiran, R.; Koduru, U. Fluorescence and absorption characteristics of dissolved organic matter excreted by phytoplankton species of western Bay of Bengal under axenic laboratory condition. J. Exp. Mar. Bio. Ecol. 2013, 445, 148–155. [Google Scholar] [CrossRef]
- Li, G.; Xie, H.; Song, G.; Gosselin, M. Production of Chromophoric Dissolved Organic Matter (CDOM) in Laboratory Cultures od Arctic Sea Ice Algae. Water 2019, 11, 926. [Google Scholar] [CrossRef] [Green Version]
- Hulatt, C.J.; Thomas, D.N.; Bowers, D.G.; Norman, L.; Zhang, C. Exudation and decomposition of chromophoric dissolved organic matter (CDOM) from some temperate macroalgae. Estuar. Coast. Shelf Sci. 2009, 84, 147–153. [Google Scholar] [CrossRef]
- Swanson, A.K.; Druehl, L.D. Induction, exudation and the UV protective role of kelp phlorotannins. Aquat. Bot. 2002, 73, 241–253. [Google Scholar] [CrossRef]
- Henderson, R.K.; Baker, A.; Parsons, S.A.; Jefferson, B. Characterisation of algogenic organic matter extracted from cyanobacteria, green algae and diatoms. Water Res. 2008, 42, 3435–3445. [Google Scholar] [CrossRef]
- Clark, C.D.; De Bruyn, W.J.; Aiona, P.D. Temporal variation in optical properties of chromophoric dissolved organic matter (CDOM) in Southern California coastal waters with nearshore kelp and seagrass. Limnol. Oceanogr. 2016, 61, 32–46. [Google Scholar] [CrossRef] [Green Version]
- Shank, G.C.; Zepp, R.G.; Vähätalo, A.; Lee, R.; Bartels, E. Photobleaching kinetics of chromophoric dissolved organic matter derived from mangrove leaf litter and floating Sargassum colonies. Mar. Chem. 2010, 119, 162–171. [Google Scholar] [CrossRef]
- Maie, N.; Scully, N.M.; Pisani, O.; Jaffé, R. Composition of a protein-like fluorophore of dissolved organic matter in coastal wetland and estuarine ecosystems. Water Res. 2007, 41, 563–570. [Google Scholar] [CrossRef]
- Fellman, J.B.; D’Amore, D.V.; Hood, E.; Boone, R.D. Fluorescence characteristics and biodegradability of dissolved organic matter in forest and wetland soils from coastal temperate watersheds in southeast Alaska. Biogeochemistry 2008, 88, 169–184. [Google Scholar] [CrossRef]
- Clark, C.D.; Litz, L.P.; Grant, S.B. Salt marshes as a source of chromophoric dissolved organic matter (CDOM) to Southern California coastal waters. Limnol. Oceanogr. 2008, 53, 1923–1933. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Chiscano, C.L. Seagrass biomass and production: A reassessment. Aquat. Bot. 1999, 65, 159–174. [Google Scholar] [CrossRef]
- Gacia, E.; Duarte, C.M.; Middelburg, J.J. Carbon and nutrient deposition in a Mediterranean seagrass Posidonia oceanica meadow. Limnol. Oceanogr. 2002, 47, 23–32. [Google Scholar] [CrossRef]
- Marbà, N.; Duarte, C.M.; Holmer, M.; Martínez, R.; Basterretxea, G.; Orfila, A.; Jordi, A.; Tintoré, J. Effectiveness of protection of seagrass Posidonia oceanica populations in Cabrera National Park (Spain). Environ. Conserv. 2002, 29, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Coscieme, L. Cultural ecosystem services: The inspirational value of ecosystems in popular music. Ecosyst. Serv. 2015, 16, 121–124. [Google Scholar] [CrossRef]
- Hemminga, M.A.; Duarte, C.M. Seagrass Ecology; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Bethoux, J.; Copin-Montégut, G. Biological fixation of atmospheric the Mediterranean Sea. Limnol. Oceanogr. 1986, 31, 1353–1358. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosci. Discuss 2005, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Marbà, N.; Gacia, E.; Fourqurean, J.W.; Beggins, J.; Barrón, C.; Apostolaki, E.T. Seagrass community metabolism: Assessing the carbon sink capacity of seagrass meadows. Glob. Biogeochem. Cycles 2010, 24, GB4032. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Kennedy, H.; Marbà, N.; Hendriks, I.E. Assessing the capacity of seagrass meadows for carbon burial: Current limitations and future strategies. Ocean Coast. Manag. 2013, 83, 32–38. [Google Scholar] [CrossRef]
- Mazarrasa, I.; Marbà, N.; Lovelock, C.E.; Serrano, O.; Lavery, P.S.; Fourqurean, J.W.; Kennedy, H.; Mateo, M.A.; Krause-Jensen, D.; Steven, A.D.; et al. Seagrass meadows as a globally significant carbonate reservoir. Biogeosciences 2015, 12, 4993–5003. [Google Scholar] [CrossRef] [Green Version]
- Macreadie, P.I.; Serrano, O.; Maher, D.T.; Duarte, C.M.; Beardall, J. Addressing calcium carbonate cycling in blue carbon accounting. Limnol. Oceanogr. Lett. 2017, 2, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.-F.; Meinesz, A. Découverte de l ’herbier de Posidonies. Cah. Parc Natl. Port Cros 1982, 4, 1–79. [Google Scholar]
- Mateo, M.Á.; Sánchez-Lizaso, J.L.; Romero, J. Posidonia oceanica “banquettes”: A preliminary assessment of the relevance for meadow carbon and nutrients budget. Estuar. Coast. Shelf Sci. 2003, 56, 85–90. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Thibaut, T.; Verlaque, M. The necromass of the Posidonia oceanica seagrass meadow: Fate, role, ecosystem services and vulnerability. Hydrobiologia 2016, 781, 25–42. [Google Scholar] [CrossRef] [Green Version]
- Barrón, C.; Duarte, C.M. Dissolved organic matter release in a Posidonia oceanica meadow. Mar. Ecol. Prog. Ser. 2009, 374, 75–84. [Google Scholar] [CrossRef]
- Apostolaki, E.T.; Holmer, M.; Marbà, N.; Karakassis, I. Degrading seagrass (Posidonia oceanica) ecosystems: A source of dissolved matter in the Mediterranean. Hydrobiologia 2010, 649, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Lavery, P.S.; McMahon, K.; Weyers, J.; Boyce, M.C.; Oldham, C.E. Release of dissolved organic carbon from seagrass wrack and its implications for trophic connectivity. Mar. Ecol. Prog. Ser. 2013, 494, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, R.F.; Cable, J.E.; Cherrier, J. Leaching and microbial degradation of dissolved organic matter from salt marsh plants and seagrasses. Aquat. Sci. 2014, 76, 595–609. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, Z.; Zhou, C.; Wu, Y.; Arbi, I.; Zhang, J.; Huang, X.; Trevathan-Tackett, S.M. Leaching of dissolved organic matter from seagrass leaf litter and its biogeochemical implications. Acta Oceanol. Sin. 2018, 37, 84–90. [Google Scholar] [CrossRef]
- Ziegler, S.; Benner, R. Effects of solar radiation on dissolved organic matter cycling in a subtropical seagrass meadow. Limnol. Oceanogr. 2000, 45, 257–266. [Google Scholar] [CrossRef]
- Otis, D.B.; Carder, K.L.; English, D.C.; Ivey, J.E. CDOM transport from the Bahamas Banks. Coral Reefs 2004, 23, 152–160. [Google Scholar] [CrossRef]
- Stabenau, E.; Zepp, R.; Bartels, E.; Zika, R. Role of the seagrass Thalassia testudinum as a source of chromophoric dissolved organic matter in coastal south Florida. Mar. Ecol. Prog. Ser. 2004, 282, 59–72. [Google Scholar] [CrossRef]
- Álvarez, E.; Grau, A.M.; Marbà, N.; Carreras, D. Praderas de angiospermas marinas Islas Baleares. In ATLAS de las Praderas Marinas de España; Ruiz, J.M., Guillén, J.E., Segura, A.R., Otero, M.M., Eds.; IEO: Murcia, Spain; IEL: Alicante, Spain; UICN: Málaga, Spain, 2015; pp. 179–220. [Google Scholar]
- Agustí, S.; Martínez-Ayala, J.; Regaudie-de-gioux, A.; Duarte, C.M. Oligotrophication and Metabolic Slowing-Down of a NW Mediterranean Coastal Ecosystem. Front. Mar. Sci. 2017, 4, 432. [Google Scholar] [CrossRef] [Green Version]
- Tovar-Sánchez, A.; Basterretxea, G.; Rodellas, V.; Sánchez-Quiles, D.; García-Orellana, J.; Masqué, P.; Jordi, A.; López, J.M.; Garcia-Solsona, E. Contribution of groundwater discharge to the coastal dissolved nutrients and trace metal concentrations in Majorca Island: Karstic vs detrital systems. Environ. Sci. Technol. 2014, 48, 11819–11827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984. [Google Scholar]
- Weishaar, J.L.; Aiken, G.R.; Bergamaschi, B.A.; Fram, M.S.; Fujii, R.; Mopper, K. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 2003, 37, 4702–4708. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zheng, B.; Song, Y.; Qin, Y. Correlation between molecular absorption spectral slope ratios and fluorescence humification indices in characterizing CDOM. Aquat. Sci. 2011, 73, 103–112. [Google Scholar] [CrossRef]
- Spencer, R.G.M.; Butler, K.D.; Aiken, G.R. Dissolved organic carbon and chromophoric dissolved organic matter properties of rivers in the USA. J. Geophys. Res. 2012, 117. [Google Scholar] [CrossRef]
- Spencer, R.G.M.; Aiken, G.R.; Butler, K.D.; Dornblaser, M.M.; Striegl, R.G.; Hernes, P.J. Utilizing chromophoric dissolved organic matter measurements to derive export and reactivity of dissolved organic carbon exported to the Arctic Ocean: A case study of the Yukon River, Alaska. Geophys. Res. Lett. 2009, 36, L06401. [Google Scholar] [CrossRef]
- Lønborg, C.; Álvarez-Salgado, X.A. Tracing dissolved organic matter cycling in the eastern boundary of the temperate North Atlantic using absorption and fluorescence spectroscopy. Deep Res. Part I 2014, 85, 35–46. [Google Scholar] [CrossRef]
- Catalá, T.S.; Martínez-Pérez, A.M.; Nieto-Cid, M.; Álvarez, M.; Otero, J.; Emelianov, M.; lReche, I.; Arístegui, J.; Álvarez-Salgado, X.A. Dissolved Organic Matter (DOM) in the open Mediterranean Sea. I. Basin–wide distribution and drivers of chromophoric DOM. Prog. Oceanogr. 2018, 165, 35–51. [Google Scholar] [CrossRef]
- Fichot, C.G.; Benner, R. A novel method to estimate DOC concentrations from CDOM absorption coefficients in coastal waters. Geophys. Res. Lett. 2011, 38, L03610. [Google Scholar] [CrossRef]
- Del Vecchio, R.; Blough, N.V. Photobleaching of chromophoric dissolved organic matter in natural waters: Kinetics and modeling. Mar. Chem. 2002, 78, 231–253. [Google Scholar] [CrossRef]
- Nelson, N.B.; Carlson, C.A.; Steinberg, D.K. Production of chromophoric dissolved organic matter by Sargasso Sea microbes. Mar. Chem. 2004, 89, 273–287. [Google Scholar] [CrossRef]
- Dahlen, J.; Bertilsson, S.; Pettersson, C. Effects of UV-A irradiation on dissolved organic matter in humic surface waters. Environ. Int. 1996, 22, 501–506. [Google Scholar] [CrossRef]
- Engelhaupt, E.; Bianchi, T.S.; Wetzel, R.G.; Tarr, M.A. Photochemical transformations and bacterial utilization of high-molecular-weight dissolved organic carbon in a southern Louisiana tidal stream (Bayou Trepagnier). Biogeochemistry 2003, 62, 39–58. [Google Scholar] [CrossRef]
- Berggren, M.; Laudon, H.; Haei, M.; Ström, L.; Jansson, M. Efficient aquatic bacterial metabolism of dissolved low-molecular-weight compounds from terrestrial sources. ISME J. 2010, 4, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Helms, J.R.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef] [Green Version]
- Iuculano, F.; Duarte, C.M.; Marbà, N.; Agustí, S. Seagrass as major source of transparent exopolymer particles in the oligotrophic Mediterranean coast. Biogeosciences 2017, 14, 5069–5075. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall: London, UK, 2008. [Google Scholar] [CrossRef]
- Llabrés, M.; Agustí, S.; Alonso-Laita, P.; Herndl, G.J. Synechococcus and Prochlorococcus cell death induced by UV radiation and the penetration of lethal UVR in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2010, 399, 27–37. [Google Scholar] [CrossRef] [Green Version]
- García-Corral, L.S.; Martinez-Ayala, J.; Duarte, C.M.; Agustí, S. Experimental assessment of cumulative temperature and UV-B radiation effects on Mediterranean plankton metabolism. Front. Mar. Sci. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Nelson, N.B.; Siegel, D.A. The global distribution and dynamics of chromophoric dissolved organic matter. Annu. Rev. Mar. Sci. 2013, 5, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.S.; Coletti, L.J. In situ ultraviolet spectrophotometry for high resolution and long-term monitoring of nitrate, bromide and bisulfide in the ocean. Deep Sea Res. Part I 2002, 49, 1291–1305. [Google Scholar] [CrossRef]
- Vila-Reixach, G.; Gasol, J.M.; Cardelús, C.; Vidal, M. Seasonal dynamics and net production of dissolved organic carbon in an oligotrophic coastal environment. Mar. Ecol. Prog. Ser. 2012, 456, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Romera-Castillo, C.; Álvarez-Salgado, X.A.; Galí, M.; Gasol, J.M.; Marrasé, C. Combined effect of light exposure and microbial activity on distinct dissolved organic matter pools. A seasonal field study in an oligotrophic coastal system (Blanes Bay, NW Mediterranean). Mar. Chem. 2013, 148, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Guallar, C.; Flos, J. Linking phytoplankton primary production and chromophoric dissolved organic matter in the sea. Prog. Oceanogr. 2019, 176, 102116. [Google Scholar] [CrossRef]
- Sánchez-Pérez, E.D.; Pujo-Pay, M.; Ortega-Retuerta, E.; Conan, P.; Peters, F.; and Marrasé, C. Mismatched dynamics of dissolved organic carbon and chromophoric dissolved organic matter in the coastal NW Mediterranean Sea. Sci. Total Environ. 2020, 746, 141190. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, H.; D’Ortenzio, F.; D’Alcalà, M.R.; Claustre, H.; Sauzède, R.; Gacic, M. On the vertical distribution of the chlorophyll a concentration in the Mediterranean Sea: A basin-scale and seasonal approach. Biogeosciences 2015, 12, 5021–5039. [Google Scholar] [CrossRef] [Green Version]
- Iuculano, F.; Álvarez-Salgado, X.A.; Otero, J.; Catalá, T.S.; Sobrino, C.; Duarte, C.M.; Agustí, S. Patterns and Drivers of UV Absorbing Chromophoric Dissolved Organic Matter in the Euphotic Layer of the Open Ocean. Front. Mar. Sci. 2019, 6, 320. [Google Scholar] [CrossRef]
- Galletti, Y.; Gonnelli, M.; Brogi, S.R.; Vestri, S.; Santinelli, C. DOM dynamics in open waters of the Mediterranean Sea: New insights from optical properties. Deep Sea Res. Part I 2019, 144, 95–114. [Google Scholar] [CrossRef]
- Lønborg, C.; Nieto-Cid, M.; Hernando-Morales, V.; Hernández-Ruiz, M.; Teira, E.; Álvarez-Salgado, X.A. Photochemical alteration of dissolved organic matter and the subsequent effects on bacterial carbon cycling and diversity. FEMS Microbiol. Ecol. 2016, 92, fiw048. [Google Scholar] [CrossRef]
- Wada, S.; Omori, Y.; Kayamyo, Y.; Tashiro, Y.; Hama, T. Photoreactivity of dissolved organic matter from macroalgae. Reg. Stud. Mar. Sci. 2015, 2, 12–18. [Google Scholar] [CrossRef]
- Jiménez, M.A.; Beltran, R.; Traveset, A.; Calleja, M.; Delgado-Huertas, A.; Marbà, N. Aeolian transport of seagrass (Posidonia oceanica) beach-cast to terrestrial systems. Estuar. Coast. Shelf Sci. 2017, 196, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Elkalay, K.; Frangoulis, C.; Skliris, N.; Goffart, A.; Gobert, S.; Lepoint, G.; Hecq, J.H. A model of the seasonal dynamics of biomass and production of the seagrass Posidonia oceanica in the Bay of Calvi (Northwestern Mediterranean). Ecol. Modell. 2003, 167, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Barrón, C.; Apostolaki, E.T.; Duarte, C.M. Dissolved organic carbon fluxes by seagrass meadows and macroalgal beds. Front. Mar. Sci. 2014, 1, 42. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cap Ses Salines | Es Caragol Beach | |||||
|---|---|---|---|---|---|---|
| Variable | Mean ± SD | Min | Max | Mean ± SD | Min | Max |
| TempW (°C) | 21.2 ± 4.7 | 13.2 | 27.8 | |||
| UVB (KJ m2 d−1) | 23.7 ± 12.2 | 4.8 | 40.5 | |||
| Chl a (mg m3) | 0.21 ± 0.14 | 0.02 | 0.54 | |||
| DOC (µmol L−1) | 90.7 ± 11.3 | 68.1 | 115.8 | 129.5 ± 64.9 | 74.0 | 380.4 |
| a254 (m−1) | 1.90 ± 0.34 | 1.34 | 2.65 | 8.79 ± 10.91 | 2.21 | 61.29 |
| a325 (m−1) | 0.43 ± 0.13 | 0.18 | 0.72 | 3.05 ± 2.93 | 0.60 | 14.80 |
| Σaλ (m−1 nm) | 112.5 ± 28.3 | 58.0 | 174.7 | 640.5 ± 622.4 | 138.3 | 3249.4 |
| a254/365 | 8.7 ± 2.3 | 5.1 | 14.2 | 4.98 ± 0.99 | 4.01 | 7.33 |
| S275–295 (µm−1) SR | 28.38 ± 4.21 2.39 ± 0.86 | 22.68 1.23 | 41.10 5.13 | 16.66 ± 3.20 1.34 ± 0.25 | 12.38 0.99 | 23.37 1.99 |
| SUVA254 (m−1 L mg C−1) | 0.79 ± 0.18 | 0.52 | 1.38 | 2.19 ± 1.17 | 0.97 | 5.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iuculano, F.; Duarte, C.M.; Otero, J.; Álvarez-Salgado, X.A.; Agustí, S. Posidonia oceanica as a Source of Chromophoric Dissolved Organic Matter for the Oligotrophic NW Mediterranean Coast. J. Mar. Sci. Eng. 2020, 8, 911. https://doi.org/10.3390/jmse8110911
Iuculano F, Duarte CM, Otero J, Álvarez-Salgado XA, Agustí S. Posidonia oceanica as a Source of Chromophoric Dissolved Organic Matter for the Oligotrophic NW Mediterranean Coast. Journal of Marine Science and Engineering. 2020; 8(11):911. https://doi.org/10.3390/jmse8110911
Chicago/Turabian StyleIuculano, Francesca, Carlos M. Duarte, Jaime Otero, Xosé Antón Álvarez-Salgado, and Susana Agustí. 2020. "Posidonia oceanica as a Source of Chromophoric Dissolved Organic Matter for the Oligotrophic NW Mediterranean Coast" Journal of Marine Science and Engineering 8, no. 11: 911. https://doi.org/10.3390/jmse8110911
APA StyleIuculano, F., Duarte, C. M., Otero, J., Álvarez-Salgado, X. A., & Agustí, S. (2020). Posidonia oceanica as a Source of Chromophoric Dissolved Organic Matter for the Oligotrophic NW Mediterranean Coast. Journal of Marine Science and Engineering, 8(11), 911. https://doi.org/10.3390/jmse8110911