
Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line




Abstract
:1. Introduction
2. Materials and Methods
2.1. Certified Reference Materials
2.2. Sample Preparation for PP2A Inhibition Assay from CRM-DSP-Mus-c and CRM-Zero-Mus
2.3. PP2A Inhibition Assay
2.4. Neuro-2a Cell Culture, Okadaic Acids Exposure, and Cell Viability Assay
3. Results
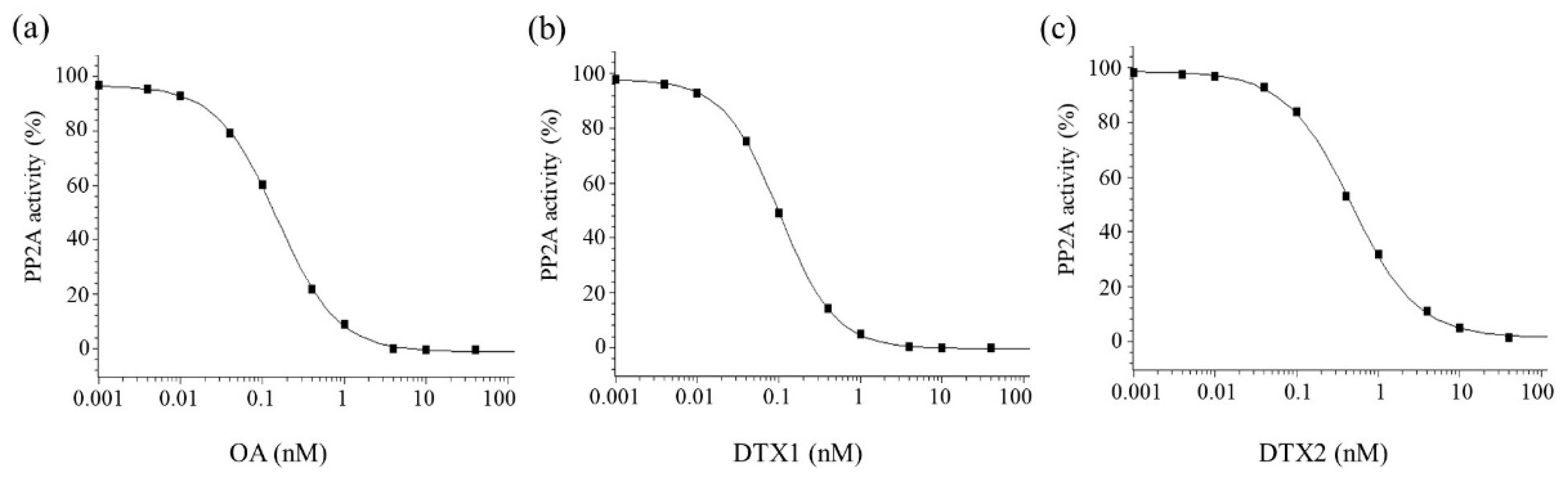
3.1. Inhibitory Potency on PP2A Activity and Neuro-2a Cytotoxicity of OA, DTX1, and DTX2
3.2. Validation of OA Equivalents Obtained through PP2A Inhibition Assay Using Reference Shellfish Tissue
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murakami, Y.; Oshima, Y.; Yasumoto, T. Identification of okadaic acid as a toxic component of a marine dinoflagellate Prorocentrum lima. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 69–72. [Google Scholar] [CrossRef]
- Hu, T.; Doyle, J.; Jackson, D.; Marr, J.; Nixon, E.; Pleasance, S.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C. Isolation of a new diarrhetic shellfish poison from irish mussels. J. Chem. Soc. Chem. Commun. 1992, 1, 39–41. [Google Scholar] [CrossRef]
- Lee, J.-S.; Igarashi, T.; Fraga, S.; Dahl, E.; Hovgaard, P.; Yasumoto, T. Determination of diarrhetic shellfish toxins in various dinoflagellate species. J. Appl. Phycol. 1989, 1, 147–152. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish. 1980, 46, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Hamano, Y.; Kinoshita, Y.; Yasumoto, T. Toxic dinoflagellates. In Proceedings of the Third International Conference on Toxic Dinoflagellates, St. Andrews, NB, Canada, 8–12 June 1985; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier: New York, NY, USA, 1985; pp. 383–388. [Google Scholar]
- Fujiki, H.; Suganuma, M.; Suguri, H.; Yoshizawa, S.; Ojika, M.; Wakamatsu, K.; Yamada, K.; Sugimura, T. Induction of ornithine decarboxylase activity in mouse skin by a possible tumor promoter, okadaic acid. Proc. Jpn. Acad. Ser. B 1987, 63, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Soliño, L.; Sureda, F.X.; Diogène, J. Evaluation of okadaic acid, dinophysistoxin-1 and dinophysistoxin-2 toxicity on neuro-2a, ng108-15 and mcf-7 cell lines. Toxicol. Vitr. 2015, 29, 59–62. [Google Scholar] [CrossRef]
- Smienk, H.; Domínguez, E.; Rodríguez-Velasco, M.L.; Clarke, D.; Kapp, K.; Katikou, P.; Cabado, A.G.; Otero, A.; Vieites, J.M.; Razquin, P.; et al. Quantitative determination of the okadaic acid toxins group by a colorimetric phosphatase inhibition assay: Interlaboratory study. J. AOAC Int. 2019, 96, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Garibo, D.; Dàmaso, E.; Eixarch, H.; de la Iglesia, P.; Fernández-Tejedor, M.; Diogène, J.; Pazos, Y.; Campàs, M. Protein phosphatase inhibition assays for okadaic acid detection in shellfish: Matrix effects, applicability and comparison with lc–ms/ms analysis. Harmful Algae 2012, 19, 68–75. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Commission regulation no. 15/2011 of january 10, 2011. Off. J. Eur. Communities 2011, 3–6. [Google Scholar]
- Codex Alimentarius International Food Standards. In Standard for Live and Raw Bivalve Molluscs Codex Standard 292-2008; Adopted in 2008. Amendment: 2013. Revision: 2014 and 2015; Food and Agriculture Organization of the United Nations: Rome, Italy; World Helth Organization: Geneva, Switzerland, 2008.
- European Commission. Regulation (ec) no 853/2004. Off. J. Eur. Union L139 2004, 47, 55–105. [Google Scholar]
- European Food Safety Authority. Marine biotoxins in shellfish - okadaic acid and analogues-scientific opinion of the panel on contaminants in the food chain. EFSA J. 2008, 6, 589. [Google Scholar] [CrossRef]
- FAO; WHO. Technical Paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; FAO: Rome, Italy, 2016; p. 47. [Google Scholar]
- Abal, P.; Louzao, M.C.; Suzuki, T.; Watanabe, R.; Vilariño, N.; Carrera, C.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Toxic action reevaluation of okadaic acid, dinophysistoxin-1 and dinophysistoxin-2: Toxicity equivalency factors based on the oral toxicity study. Cell. Physiol. Biochem. 2018, 49, 743–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Shimatani, M.; Sugitani, H.; Oshima, Y.; Yasumoto, T. Isolation and structural elucidation of the causative toxin of the diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef]
- Takai, A.; Mieskes, G. Inhibitory effect of okadaic acid on the p-nitrophenyl phosphate phosphatase activity of protein phosphatases. Biochem. J. 1991, 275, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Tubaro, A.; Florio, C.; Luxich, E.; Sosa, S.; Loggia, R.D.; Yasumoto, T. A protein phosphatase 2a inhibition assay for a fast and sensitive assessment of okadaic acid contamination in mussels. Toxicon Off. J. Int. Soc. Toxinol. 1996, 34, 743–752. [Google Scholar] [CrossRef]
- Simon, J.F.; Vernoux, J.P. Highly sensitive assay of okadaic acid using protein phosphatase and paranitrophenyl phosphate. Nat. Toxins 1994, 2, 293–301. [Google Scholar] [CrossRef]
- Ikehara, T.; Nakashima, S.; Nakashima, J.; Kinoshita, T.; Yasumoto, T. Efficient production of recombinant pp2a at a low temperature using a baculovirus expression system. Biotechnol. Rep. 2016, 11, 86–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikehara, T.; Shinjo, F.; Ikehara, S.; Imamura, S.; Yasumoto, T. Baculovirus expression, purification, and characterization of human protein phosphatase 2a catalytic subunits α and β. Protein Expr. Purif. 2006, 45, 150–156. [Google Scholar] [CrossRef]
- Ikehara, T.; Imamura, S.; Yoshino, A.; Yasumoto, T. Pp2a inhibition assay using recombinant enzyme for rapid detection of okadaic acid and its analogs in shellfish. Toxins 2010, 2, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Ikehara, T.; Kinoshita, T.; Kurokawa, A.; Nakashima, S.; Maekawa, K.; Ohshiro, N.; Yasumoto, T. Evaluation of protein phosphatase 2A (PP2A) inhibition assay for rapid detection of dsp toxins in scallop. NIPPON SUISAN GAKKAISHI 2017, 83, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Bodero, M.; Bovee, T.F.H.; Wang, S.; Hoogenboom, R.L.A.P.; Klijnstra, M.D.; Portier, L.; Hendriksen, P.J.M.; Gerssen, A. Screening for the presence of lipophilic marine biotoxins in shellfish samples using the neuro-2a bioassay. Food Addit. Contam. Part A 2018, 35, 351–365. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Yamaguchi, H.; Tanabe, Y.; Sano, T.; Yoshimoto, M.; Oshiro, N.; Nakashima, S.; Yasumoto-Hirose, M. First report of microcystis strains producing mc-fr and -wr toxins in japan. Toxins 2019, 11, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeds, J.R.; Stutts, W.L.; Celiz, M.D.; MacLeod, J.; Hamilton, A.E.; Lewis, B.J.; Miller, D.W.; Kanwit, K.; Smith, J.L.; Kulis, D.M.; et al. Dihydrodinophysistoxin-1 produced by dinophysis norvegica in the gulf of maine, USA and its accumulation in shellfish. Toxins 2020, 12, 533. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Wu, P.; Wei, J.; Luo, J.; Ding, P.; Wei, H.; Yang, F. A mini-review on detection methods of microcystins. Toxins 2020, 12, 641. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxins | PP2A Inhibition | Cytotoxicity | Mouse Lethality | |||
|---|---|---|---|---|---|---|
| IC50 (nM) | OApp2a a | EC50 (nM) | OAn2a b | OAip c | OAor d | |
| OA | 0.14 | 1 | 21.6 | 1 | 1 | 1 |
| DTX1 | 0.09 | 1.6 | 14.1 | 1.5 | 1 | 1.5 |
| DTX2 | 0.45 | 0.3 | 41.0 | 0.5 | 0.5 | 0.3 |
| Matrix | Mussels | |
|---|---|---|
| Unhydrolyzed | Hydrolyzed | |
| Mean a (μg/g) | 0.0225 | 0.0499 |
| SD | ±0.0054 | ±0.0049 |
| LOD b (μg/g) | 0.0387 | 0.0646 |
| LOQ c (μg/g) | 0.0765 | 0.0989 |
| Sample no. | OA Equivalents (μg/g) | Accuracy (%)(a)/(b)×100 | ||
|---|---|---|---|---|
| PP2A Inhibition Assay Result in the Present Study (a) | Expected Equivalent * according to the OApp2a (b) | |||
| Mean | RSD (%) | |||
| 1 | 0.5487 | 5.6 | 0.6080 | 90.3 |
| 2 | 0.3282 | 3.6 | 0.3040 | 108.0 |
| 3 | 0.1733 | 7.8 | 0.1507 | 115.0 |
| 4 | 0.5293 | 2.4 | 0.4820 | 109.8 |
| 5 | 0.2798 | 6.4 | 0.2410 | 116.1 |
| 6 | 0.1332 | 1.7 | 0.1213 | 109.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikehara, T.; Chikanishi, K.; Oshiro, N. Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line. J. Mar. Sci. Eng. 2021, 9, 1140. https://doi.org/10.3390/jmse9101140
Ikehara T, Chikanishi K, Oshiro N. Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line. Journal of Marine Science and Engineering. 2021; 9(10):1140. https://doi.org/10.3390/jmse9101140
Chicago/Turabian StyleIkehara, Tsuyoshi, Kazuya Chikanishi, and Naomasa Oshiro. 2021. "Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line" Journal of Marine Science and Engineering 9, no. 10: 1140. https://doi.org/10.3390/jmse9101140
APA StyleIkehara, T., Chikanishi, K., & Oshiro, N. (2021). Specification of the Okadaic Acid Equivalent for Okadaic Acid, Dinophysistoxin-1, and Dinophysistoxin-2 Based on Protein Phosphatase 2A Inhibition and Cytotoxicity Assays Using Neuro 2A Cell Line. Journal of Marine Science and Engineering, 9(10), 1140. https://doi.org/10.3390/jmse9101140