
Vertical Configuration of a Side Scan Sonar for the Monitoring of Posidonia oceanica Meadows

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
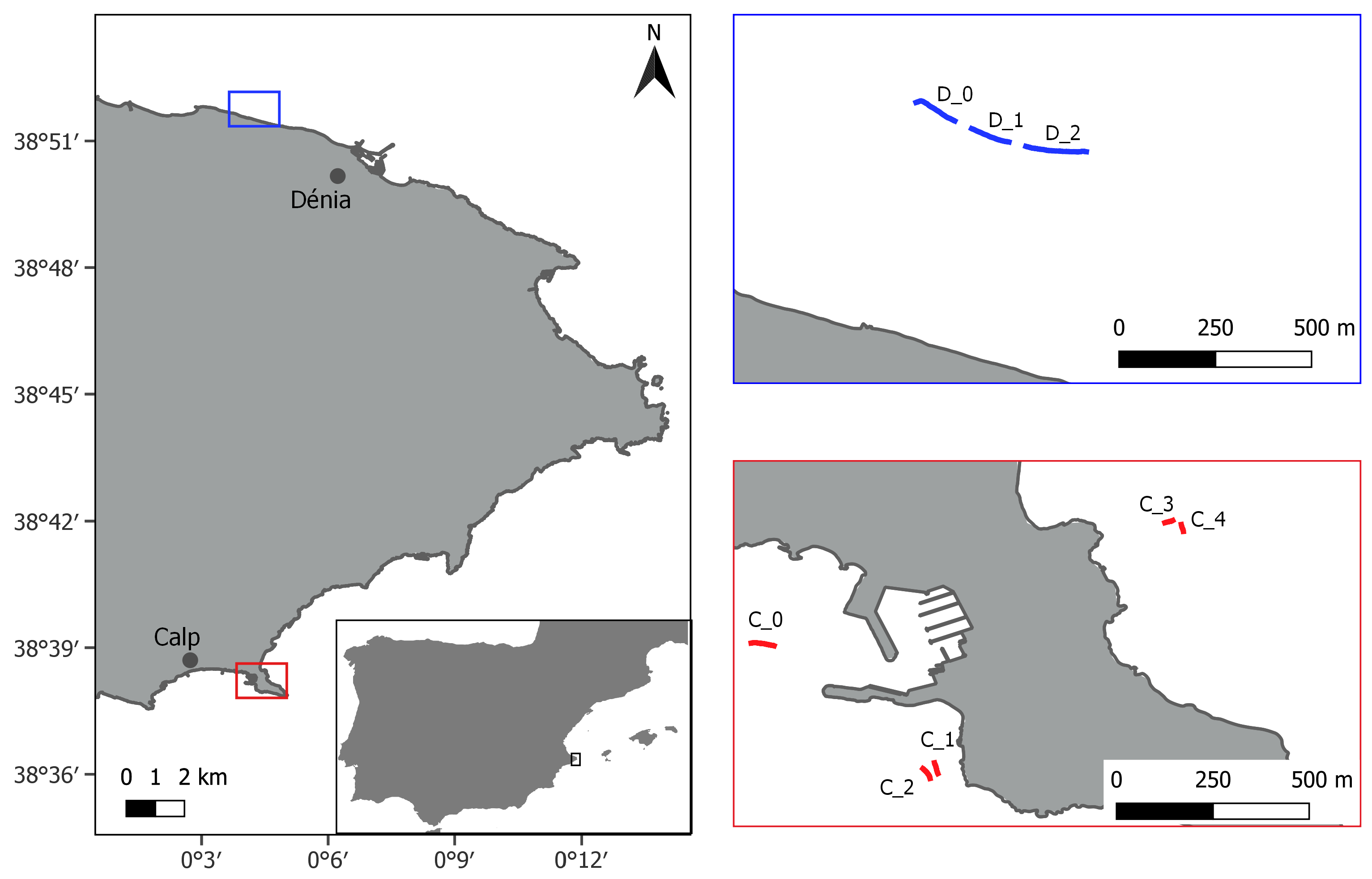
2.1. Study Area
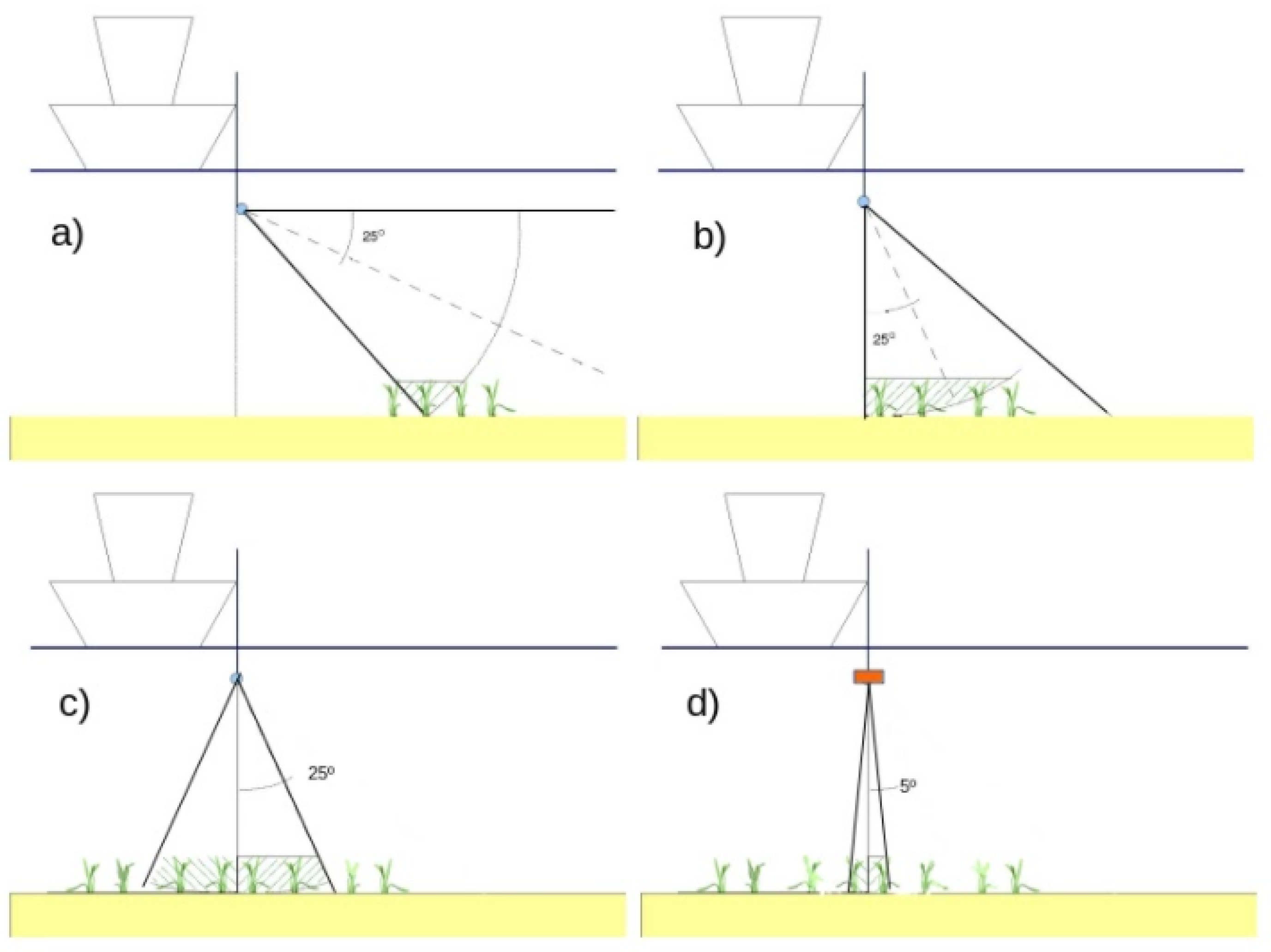
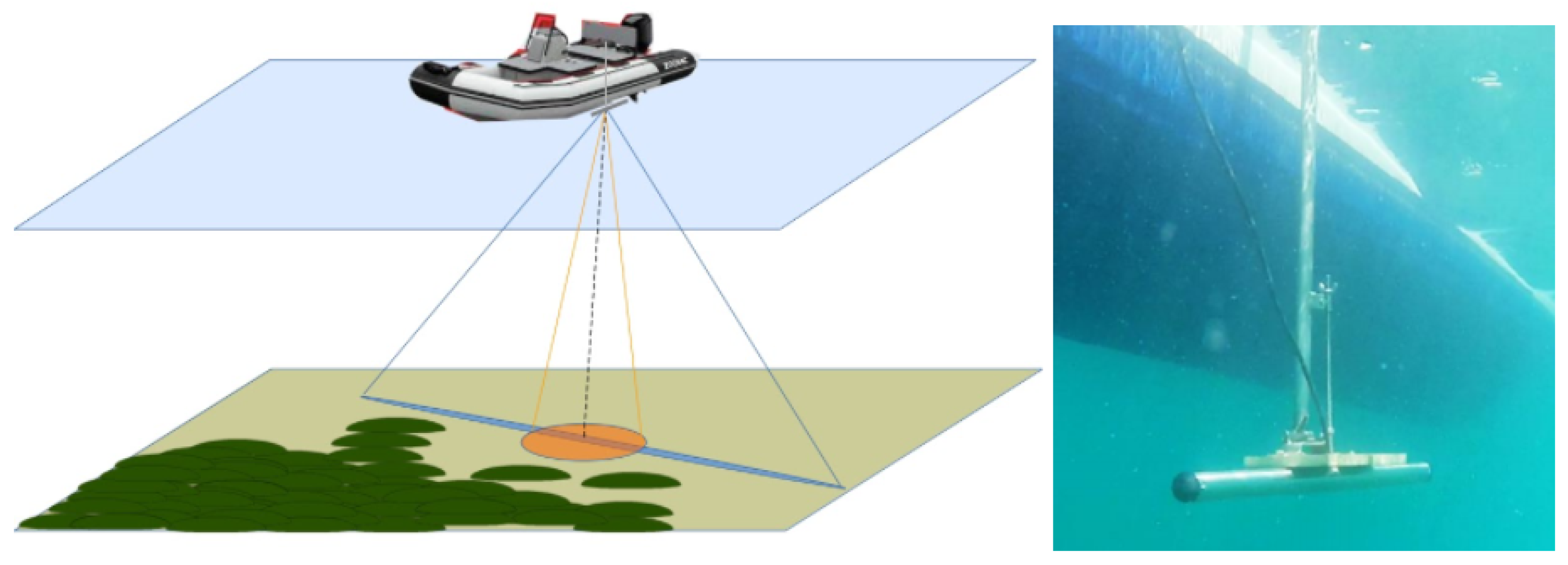
2.2. Acoustic Data Acquisition
2.3. Ground-Truthing
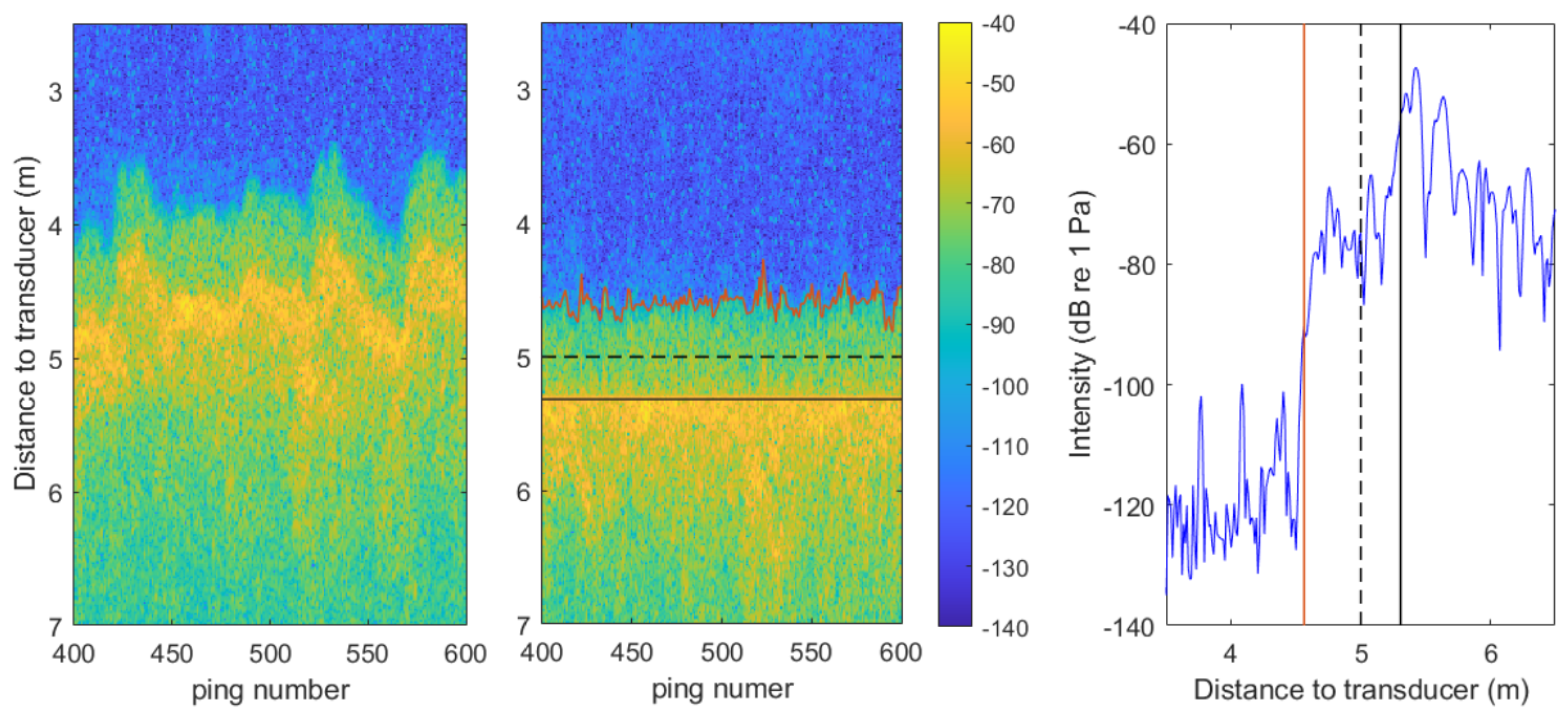
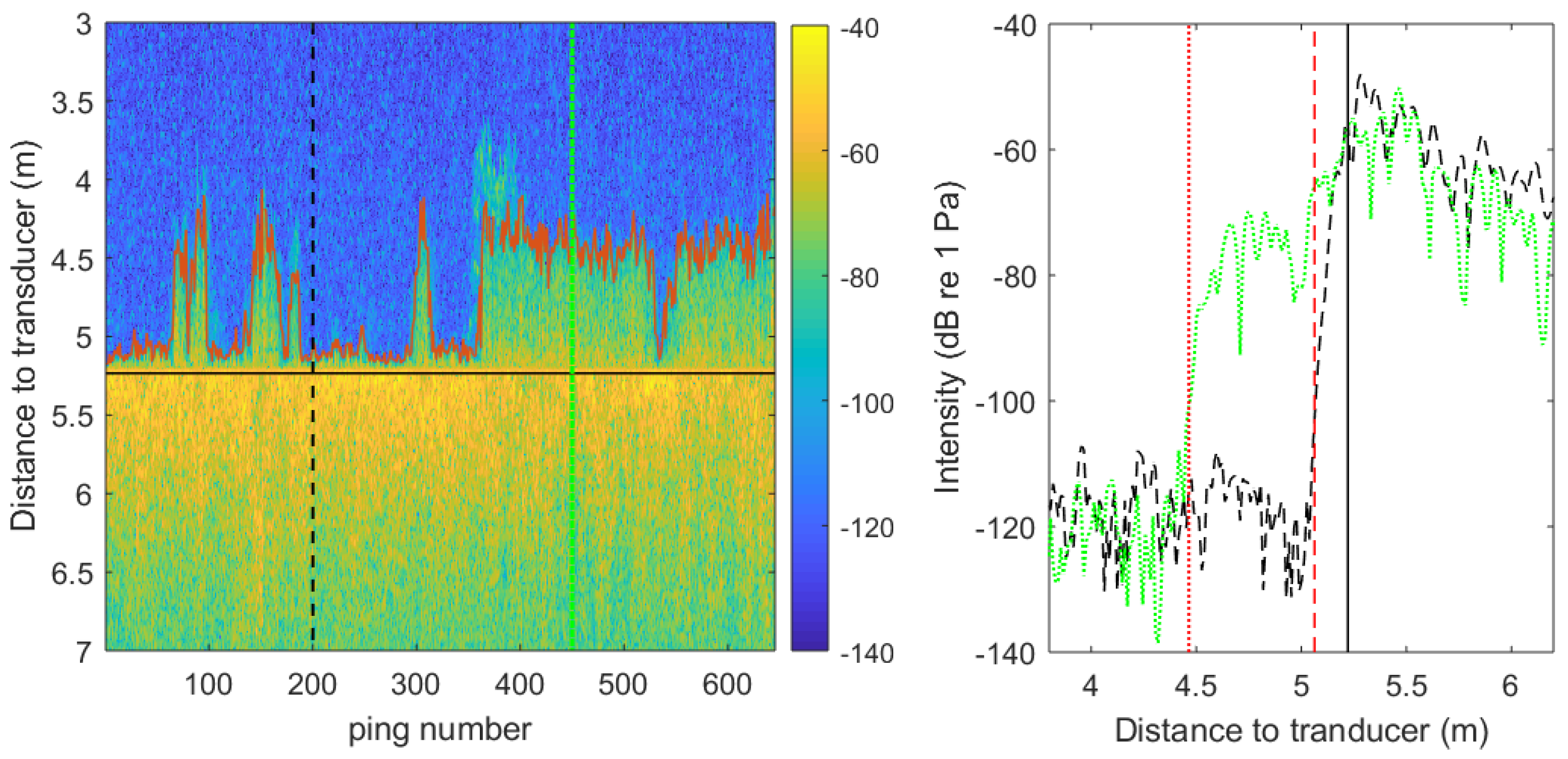
2.4. Acoustic Data Processing
3. Results and Discussion
3.1. Density and Cover Direct Estimation Values
3.2. Acoustic Detection and Characterization of P. oceanica Meadows
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Picard, J. Recherches qualitatives sur les biocoenoses marines des substrats meubles dragables de la région marseillaise. Rec. Trav. Stn. Mar. Endoume Fac. Sci. 2009, 36, 1–60. [Google Scholar]
- Marbà, N.; Duarte, C.M.; Cebrián, J.; Gallegos, M.E.; Olesen, B.; Sand-Jensen, K. Growth and population dynamics of Posidonia oceanica on the Spanish Mediterranean coast: Elucidating seagrass decline. Mar. Ecol. Prog. Ser. 1996, 137, 203–213. [Google Scholar] [CrossRef]
- Montefalcone, M.; Parravicini, V.; Vacchi, M.; Albertelli, G.; Ferrari, M.; Morri, C.; Bianchi, C.N. Human influence on seagrass habitat fragmentation in NW Mediterranean Sea. Estuar. Coast. Shelf Sci. 2010, 86, 292–298. [Google Scholar] [CrossRef]
- Boudouresque, C.F. The Mediterranean Sea: An Overview of its Present State and Plans for Future Protection. In The Erosion of Mediterranean Biodiversity; Rodríguez-Prieto, C., Pardini, G., Eds.; Servei de Publicacions de la Universitat de Girona: Girona, Spain, 2003; pp. 53–112. [Google Scholar]
- Boudouresque, C.; Bernard, G.; Bonhomme, P.; Charbonnel, E.; Diviacco, G.; Meinesz, A.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Tunesi, L. Protection and Conservation of Posidonia Oceanica Meadows; Ramoge and RAC/SPA Publisher: Tunis, Marseille, 2012; p. 202. [Google Scholar]
- Coleman, F.C.; Williams, S.L. Overexploiting marine ecosystem engineers: Potential consequences for biodiversity. Trends Ecol. Evol. 2002, 17, 40–44. [Google Scholar] [CrossRef]
- Personnic, S.; Bodouresque, C.F.; Astruch, P.; Ballesteros, E.; Blouet, E.; Bella-Santini, D.; Bonhomme, P.; Thibault-Botha, D.; Feuteun, E.; Harmelin-Vivien, M.; et al. An Ecosystem-Based Approach to Assess the Status of a Mediterranean Ecosystem, the Posidonia oceanica Seagrass Meadow. PLoS ONE 2014, 6, e98994. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Mayot, N.; Pergent, G. The outstanding traits of the functioning of the Posidonia oceanica seagrass ecosystem. Biol. Mar. Medit. 2006, 13, 109–113. [Google Scholar]
- de los Santos, C.B.; Krause-Jensen, D.; Alcoverro, T.; Marbà, N.; Duarte, C.M.; Van Katwijk, M.M.; Pérez, M.; Romero, J.; Sánchez-Lizaso, J.L.; Roca, G.; et al. Recent trend reversal for declining European seagrass meadows. Nat. Commun. 2019, 10, 3356. [Google Scholar] [CrossRef]
- Jordà, G.; Marbà, M.; Duarte, C.M. Mediterranean seagrass vulnerable to regional climate warming. Nat. Clim. Chang. 2012, 2, 821–824. [Google Scholar] [CrossRef]
- Ballesteros, E.; Cebrián, E.; Alcoverro, T. Mortality of shoots of Posidonia oceanica following meadow invasion by red alga Lophocladia lallemandii. Bot. Mar. 2007, 50, 8–13. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Terrados, J.; Deudero, S.; Pons, A. Antioxidant response of the seagrass Posidonia oceanica when epiphytized by the invasive macroalgae Lophocladia lallemandi. Mar. Environ. Res. 2008, 66, 359–363. [Google Scholar] [CrossRef]
- De Villéle, X.; Verlaque, M. Changes and degradation in a Posidonia oceanica bed invaded by the introduced tropical alga Caulerpa taxifolia in the north western Mediterranean. Bot. Mar. 1995, 38, 79–88. [Google Scholar] [CrossRef]
- Micheli, C.; Paganin, P.; Peirano, A.; Caye, G.; Meinesz, A.; Bianchi, C.N. Genetic variability of Posidonia oceanica (L.) Delile in relation to local factors and biogeographic patterns. Aquat. Bot. 2005, 82, 210–221. [Google Scholar] [CrossRef]
- Schuegraf, M.J. Establishment of Seagrass Decline and Causative Mechanisms in Pearl Lagoon, Nicaragua through Use of traditional Ecological Knowledge, Sediment Coring and Direct Visual Census; FES Outstanding Graduate Student Paper Series; University of New York: New York, NY, USA, 2004; Volume 9. [Google Scholar]
- Newmaster, A.F.; Berg, K.J.; Ragupathy, S.; Palanisamy, M.; Sambandan, K.; Newmaster, S.G. Local knowledge and conservation of seagrasses in the Tamil Nadu State of India. J. Ethnobiol. Ethnomed. 2011, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pergent-Martini, C.; Leoni, V.; Pasqualini, V.; Ardizzone, G.D.; Balestri, E.; Bedini, R.; Belluscio, A.; Belsher, T.; Borg, J.; Boudouresque, C.F.; et al. Descriptors of Posidonia oceanica meadows: Use and application. Ecol. Indic. 2005, 5, 213–230. [Google Scholar] [CrossRef]
- Crawford, C.M.; Mitchell, I.M.; Macleod, C.K.A. Video assessment of environmental impacts of salmon farms. ICES J. Mar. Sci. 2001, 58, 445–452. [Google Scholar] [CrossRef]
- Paul, M.; Lefebvre, A.; Manca, E.; Amos, C.L. An acoustic method for the remote measurement of seagrass metrics. Estuar. Coast. Shelf Sci. 2011, 93, 68–79. [Google Scholar] [CrossRef]
- Traganos, D.; Reinartz, P. Mapping Mediterranean seagrasses with Sentinel-2 imagery. Mar. Pollut. Bull. 2018, 134, 197–209. [Google Scholar] [CrossRef]
- Vis, C.; Hudon, C.; Carignan, R. An evaluation of approaches used to determine the distribution and biomass of emergent and submerged aquatic macrophytes over large spatial scales. Aquat. Bot. 2003, 77, 187–201. [Google Scholar] [CrossRef]
- McCarthy, E.; Sabol, B. Acoustic characterization of submerged aquatic vegetation: Military and environmental monitoring applications. In Proceedings of the Oceans 2000 MTS/IEEE Conference and Exhibition, Providence, RI, USA, 11–14 September 2000; pp. 1957–1961. [Google Scholar]
- Simmonds, E.J.; MacLennan, D.N. Fisheries Acoustics: Theory and Practice; Blackwell Publishing: London, UK, 2005. [Google Scholar]
- Anderson, J.T.; Van Holliday, D.; Kloser, R.; Reid, D.G.; Simard, Y. Acoustic seabed classification: Current practice and future directions. ICES J. Mar. Sci. 2008, 65, 1004–1011. [Google Scholar] [CrossRef]
- Gumusay, M.U.; Bakirman, T.; Kizilkaya, I.T.; Aykut, N.O. A review of seagrass detection, mapping and monitoring applications using acoustic systems. Eur. J. Remote Sens. 2019, 52, 1–29. [Google Scholar] [CrossRef]
- Mamede, R.; Rodrigues, A.M.; Freitas, R.; Quintino, V. Single-beam acoustic variability associated with seabed habitats. J. Sea Res. 2015, 100, 152–159. [Google Scholar] [CrossRef]
- Jordan, A.; Lawler, M.; Halley, V.; Barret, N. Seabed habitat mapping in the Kent Group of islands and its role in Marine protected area planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 51–70. [Google Scholar] [CrossRef]
- Brown, C.J.; Blondel, P. Developments in the application of multibeam sonar backscatter for seafloor habitat mapping. Appl. Acoust. 2009, 70, 1242–1247. [Google Scholar] [CrossRef]
- Komatsu, T.; Igarashi, C.; Tatsukawa, K.; Sultana, S.; Matsuoka, Y.; Harada, S. Use of multi-beam sonar to map seagrass beds in Otsuchi Bay on the Sanriku Coast of Japan. Aquat. Living Resour. 2003, 16, 223–230. [Google Scholar] [CrossRef]
- Di Maida, G.; Tomasello, A.; Luzzu, F.; Scannavino, A.; Pirrotta, M.; Orestano, C.; Calvo, S. Discriminating between Posidonia oceanica meadows and sand substratum using multibeam sonar. ICES J. Mar. Sci. 2011, 68, 12–19. [Google Scholar] [CrossRef]
- Hamana, M.; Komatsu, T. Real-Time Classification of Seagrass Meadows on Flat Bottom with Bathymetric Data Measured by a Narrow Multibeam Sonar System. Remote Sens. 2016, 8, 96. [Google Scholar] [CrossRef]
- Greene, A.; Rahman, A.F.; Kline, R.; Rahman, M.S. Side scan sonar: A cost-efficient alternative method for measuring seagrass cover in shallow environments. Estuar. Coast. Shelf Sci. 2018, 207, 250–258. [Google Scholar] [CrossRef]
- Piazzi, L.; Acunto, S.; Cinelli, F. Mapping of Posidonia oceanica beds around Elba Island (western Mediterranean) with integration of direct and indirect methods. Estuar. Coast. Shelf Sci. 2000, 23, 339–346. [Google Scholar] [CrossRef]
- Sánchez-Carnero, N.; Rodríguez-Pérez, D.; Couñago, E.; Aceña, S.; Freire, J. Using vertical Sidescan Sonar as a tool for seagrass cartography. Estuar. Coast. Shelf Sci. 2012, 115, 334–344. [Google Scholar] [CrossRef]
- Rahnemoonfar, M.; Rahman, A.F.; Kline, R.J.; Greene, A. Automatic seagrass disturbance pattern identification on sonar images. IEEE J. Ocean. Eng. 2018, 44, 132–141. [Google Scholar] [CrossRef]
- Collier, J.S.; Brown, C.J. Correlation of sidescan backscatter with grain size distribution of surficial seabed sediments. Mar. Geol. 2005, 214, 431–449. [Google Scholar] [CrossRef]
- Ahmad-Kamil, E.I.; Ramli, R.; Jaaman, S.A.; Bali, J.; Al-Obaidi, J.R. The effects of water parameters on monthly seagrass percentage cover in Lawas, East Malaysia. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Lurton, X. An Introduction to Underwater Acoustics: Principles and Applications; Springer Science and Business Media: Chichester, UK, 2002. [Google Scholar]
- François, R.E.; Garrison, G.R. Sound absorption based on ocean measurements. Part I: Pure water and magnesium sulfate contributions. JASA 1982, 72, 896–907. [Google Scholar] [CrossRef]
- François, R.E.; Garrison, G.R. Sound absorption based on ocean measurements. Part II: Boric acid contribution and equation for total absorption. JASA 1982, 72, 1879–1890. [Google Scholar] [CrossRef]
- Mackenzie, K.V. Nine-term equation for sound speed in the oceans. JASA 1981, 70, 807–812. [Google Scholar] [CrossRef]
- Balestri, E. Flowering of the seagrass Posidonia oceanica in a north-western Mediterranean coastal area: Temporal and spatial variations. Mar. Biol. 2004, 145, 61–68. [Google Scholar] [CrossRef]
- Ciarolo, G.; Ferreri, G.B.; Loggia, G.L. Flow resistance of Posidonia oceanica in shallow water. J. Hydraul. Res. 2006, 44, 189–202. [Google Scholar] [CrossRef]
- Olesen, B.; Enríquez, S.; Duarte, C.M.; Sand-Jensen, K. Depth-acclimation of photosynthesis, morphology and demography of Posidonia oceanica and Cymodocea nodosa in the Spanish Mediterranean Sea. Mar. Ecol. Prog. Ser. 2002, 236, 89–97. [Google Scholar] [CrossRef]
- Olguner, C.; Mutlu, E. Acoustic Estimates of Leaf Height and Biomass of Posidonia oceanica Meadow in Gulf of Antalya, The Eastern Mediterranean. COMU-JMSF 2020, 3, 79–94. [Google Scholar] [CrossRef]
- Dalla Via, J.; Sturmbauer, C.; Schönweger, G.; Sötz, E.; Mathekowitsch, S.; Stifter, M.; Rieger, R. Light gradients and meadow structure in Posidonia oceanica: Ecomorphological and functional correlates. Mar. Ecol. Prog. Ser. 1998, 163, 267–278. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Middelboe, A.L.; Sand-Jensen, K.; Christensen, P.B. Eelgrass, Zostera marina, growth along depth gradients: Upper boundaries of the variation as a powerful predictive tool. Oikos 2000, 91, 233–244. [Google Scholar] [CrossRef]
- Hamilton, L.J. Acoustic Seabed Classification Systems; Defence Science and Technology Organisation; DSTO Aeronautical and Maritime Research Laboratory: Victoria, Australia, 2001. [Google Scholar]
- Rodríguez-Pérez, D.; Sánchez-Carnero, N.; Freire, J. A pulse-length correction to improve energy-based seabed classification in coastal areas. Cont. Shelf Res. 2014, 77, 1–13. [Google Scholar] [CrossRef]
- Delgado, O.; Ruiz, J.; Pérez, M.; Romero, J.; Ballesteros, E. Effects of fish farming on seagrass (Posidonia oceanica) in a Mediterranean bay: Seagrass decline after organic loading cessation. Oceanol. Acta 1999, 22, 109–117. [Google Scholar] [CrossRef]
- Terrados, J.; Medina-Pons, F.J. Inter-annual variation of shoot density and biomass, nitrogen and phosphorus content of the leaves, and epiphyte load of the seagrass Posidonia oceanica (L.) Delile off Mallorca, western Mediterranean. Sci. Mar. 2011, 75, 61–70. [Google Scholar] [CrossRef]
- Hermand, J.P. Continuous acoustic monitoring of physiological and environmental processes in seagrass prairies with focus on photosynthesis. In Acoustic Sensing Techniques for the Shallow Water Environment; Caiti, A., Chapman, N.R., Hermand, J.P., Jesus, M., Eds.; Springer: Dordrecht, The Nederland, 2006; pp. 183–196. [Google Scholar]
- Felisberto, P.; Jesus, S.M.; Zabel, F.; Santos, R.; Silva, J.; Gobert, S.; Beer, S.; Njörl, M.; Mazzuca, S.; Procaccini, G.; et al. Acoustic monitoring of O2 production of a seagrass meadow. J. Exp. Mar. Biol. Ecol. 2015, 464, 75–87. [Google Scholar] [CrossRef]
- Wilson, C.J.; Wilson, P.S.; Greene, C.A.; Dunton, K.H. Seagrass meadows provide an acoustic refuge for estuarine fish. Mar. Ecol. Prog. Ser. 2013, 472, 117–127. [Google Scholar] [CrossRef]
- Barguet, L.; Romero-García, V.; Jiménez, N.; Garcia-Raffi, L.M.; Sánchez-Morcillo, V.J.; Groby, J.P. Natural sonic crystal absorber constituted of seagrass (Posidonia Oceanica) fibrous spheres. Sci. Rep. 2021, 11, 711. [Google Scholar] [CrossRef]
- Freitas, R.; Sampaio, L.; Rodrigues, A.M.; Quintino, V. Sea-bottom classification across a shallow-water bar channel and near-shore shelf, using single-beam acoustics. Estuar. Coast. Shelf Sci. 2005, 65, 625–632. [Google Scholar] [CrossRef]
- Ferretti, R.; Fumagalli, E.; Caccia, M.; Bruzzone, G. Seabed classification using a single beam echosounder. In Proceedings of the Oceans 2015-Genova, IEEE Conference, Genova, Italy, 18–21 May 2015; pp. 1–5. [Google Scholar]
- Snellen, M.; Siemes, K.; Simons, D.G. Model-based sediment classification using single-beam echosounder signals. JASA 2011, 129, 2878–2888. [Google Scholar] [CrossRef]
- Foote, K.G.; Knudsen, H.P.; Vestnes, G.; MacLennan, D.N.; Simmonds, E.J. Calibration of Acoustic Instruments for Fish Density Estimation: A Practical Guide; Cooperative Research Report 144; International Council for the Exploration of the Sea: Copenhagen, Denmark, 1987; p. 69. [Google Scholar]
- Demer, D.A.; Berger, L.; Bernasconi, M.; Bethke, E.; Boswell, K.; Chu, D.; Domokos, R.; Dunford, A.; Fässler, S.; Gauthier, S.; et al. Calibration of Acoustic Instruments; ICES Cooperative Research Report 326; International Council for the Exploration of the Sea (ICES) Publisher: Copenhagen, Denmark, 2015. [Google Scholar]
- Tseng, Y.T. Recognition and Assessment of Seafloor Vegetation Using a Single Beam Echosounder. Ph.D. Thesis, Curtin University, Bentley, Australia, 2009. [Google Scholar]
- Monpert, C.; Legris, M.; Noel, C.; Zerr, B.; Le Caillec, J.M. Studying and modeling of submerged aquatic vegetation environments seen by a single beam echosounder. Proc. Meet. Acoust. 2012, 17, 070044. [Google Scholar]
- Barrell, J.; Grant, J. Detecting hot and cold spots in a seagrass landscape using local indicators of spatial association. Landsc. Ecol. 2013, 28, 2005–2018. [Google Scholar] [CrossRef]
- Chamberlain, R.H.; Doering, P.H.; Orlando, B.; Sabol, B.M. Comparison of manual and hydroacoustic measurement of seagrass distribution in the Caloosahatchee estuary. Fla. Sci. 2009, 72, 386–405. [Google Scholar]
- Sabol, B.M.; Melton, R.E.; Chamberlain, R.; Doering, P.; Haunert, K. Evaluation of a digital echo sounder system for detection of submersed aquatic vegetation. Estuaries 2002, 25, 133–141. [Google Scholar] [CrossRef]
- Quintino, V.; Freitas, R.; Mamede, R.; Ricardo, F.; Rodrigues, A.M.; Mota, J.; Pérez-Ruzafa, A.; Marcos, C. Remote sensing of underwater vegetation using single-beam acoustics. ICES J. Mar. Sci. 2010, 67, 594–605. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Length (m) | Depth (m) |
|---|---|---|
| D_0 | 110 | 6 |
| D_1 | 110 | 5 |
| D_2 | 162 | 5 |
| C_0 | 60 | 2.5 |
| C_1 | 37 | 9 |
| C_2 | 39 | 12.5 |
| C_3 | 22 | 4.5 |
| C_4 | 24 | 6 |
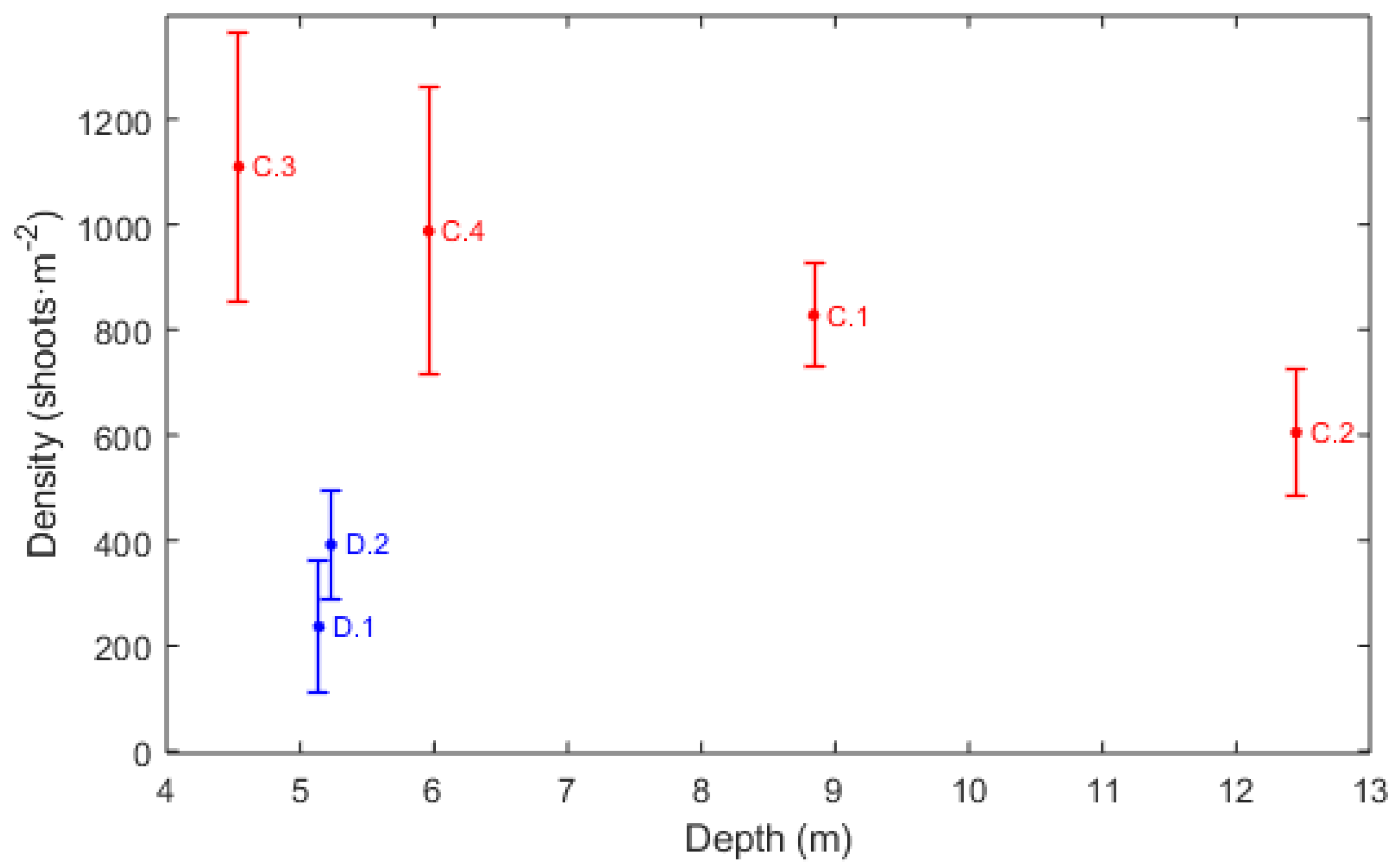
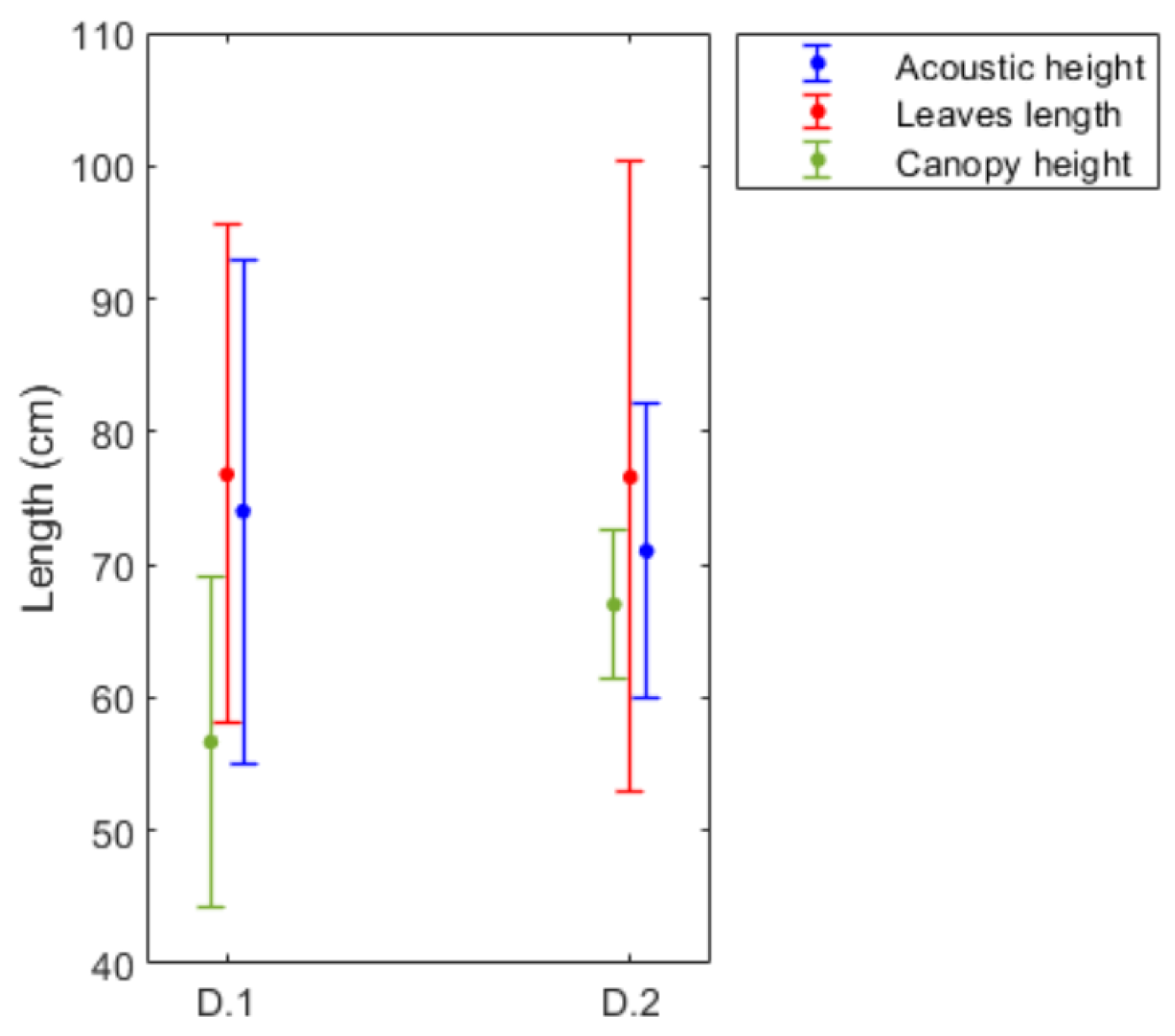
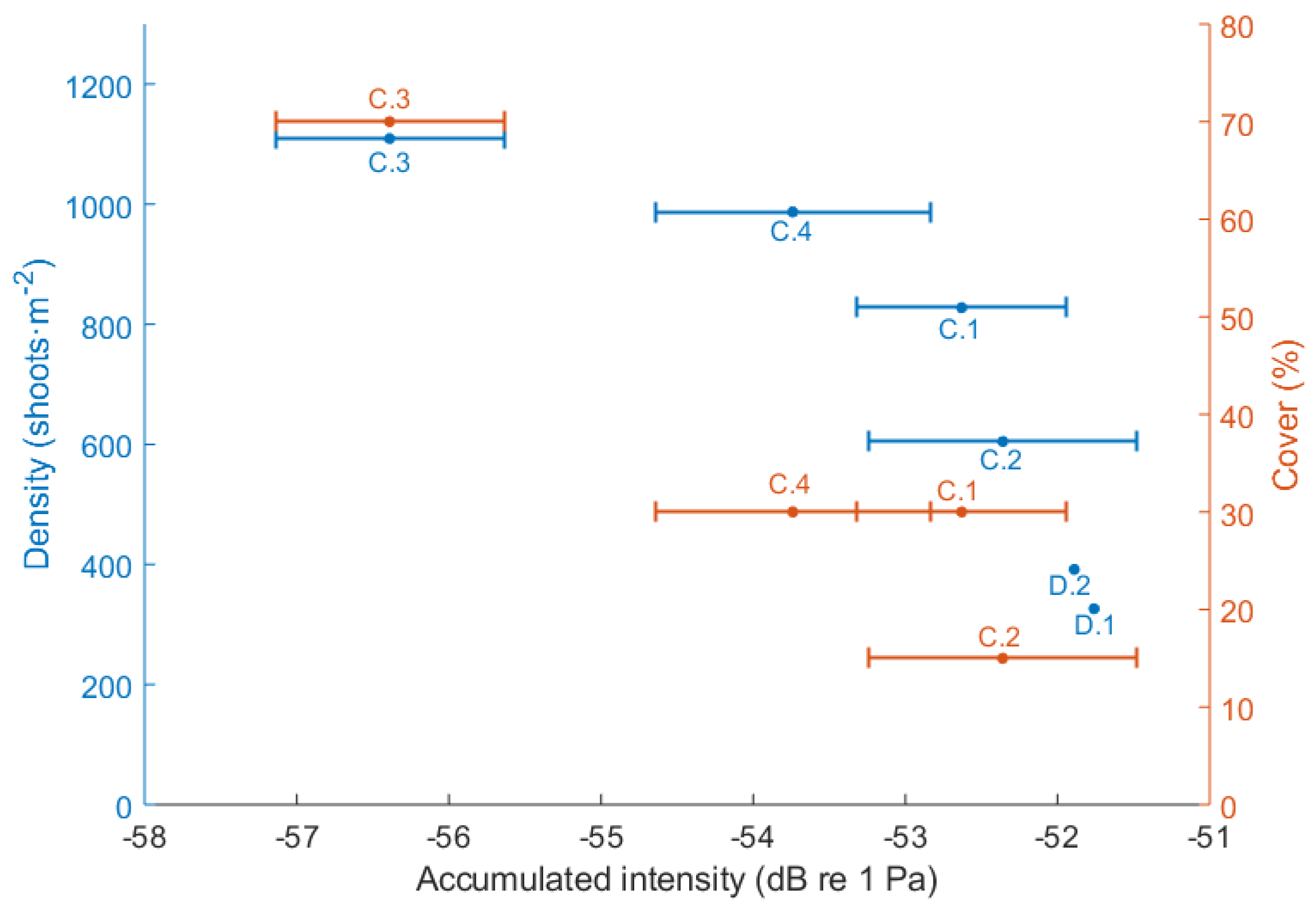
| Stations | Density Stimation (Shoots·m) | Leaf Length (m) | Canopy Height (m) | Cover (%) |
|---|---|---|---|---|
| D_1 | 326.30 ± 125.34 | 76.8 ± 18.73 | 56.68 ± 12.43 | - |
| D_2 | 391.87 ± 104.25 | 76.6 ± 23.67 | 67 ± 5.58 | - |
| C_1 | 827.64 ± 97.61 | - | - | 30 |
| C_2 | 604.80 ± 122.47 | - | - | 15 |
| C_3 | 1109.33 ± 255.18 | - | - | 70 |
| C_4 | 987.20 ± 272.48 | - | - | 30 |
| Stations | Accumulated Intensity (dB) |
|---|---|
| D_1 | −51.76 |
| D_2 | −51.89 |
| C_1 | −52.63 ± 0.69 |
| C_2 | −52.36 ± 0.88 |
| C_3 | −56.32 ± 0.75 |
| C_4 | −53.74 ± 0.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llorens-Escrich, S.; Tamarit, E.; Hernandis, S.; Sánchez-Carnero, N.; Rodilla, M.; Pérez-Arjona, I.; Moszynski, M.; Puig-Pons, V.; Tena-Medialdea, J.; Espinosa, V. Vertical Configuration of a Side Scan Sonar for the Monitoring of Posidonia oceanica Meadows. J. Mar. Sci. Eng. 2021, 9, 1332. https://doi.org/10.3390/jmse9121332
Llorens-Escrich S, Tamarit E, Hernandis S, Sánchez-Carnero N, Rodilla M, Pérez-Arjona I, Moszynski M, Puig-Pons V, Tena-Medialdea J, Espinosa V. Vertical Configuration of a Side Scan Sonar for the Monitoring of Posidonia oceanica Meadows. Journal of Marine Science and Engineering. 2021; 9(12):1332. https://doi.org/10.3390/jmse9121332
Chicago/Turabian StyleLlorens-Escrich, Susana, Elena Tamarit, Sebastián Hernandis, Noela Sánchez-Carnero, Miguel Rodilla, Isabel Pérez-Arjona, Marek Moszynski, Vicente Puig-Pons, José Tena-Medialdea, and Víctor Espinosa. 2021. "Vertical Configuration of a Side Scan Sonar for the Monitoring of Posidonia oceanica Meadows" Journal of Marine Science and Engineering 9, no. 12: 1332. https://doi.org/10.3390/jmse9121332
APA StyleLlorens-Escrich, S., Tamarit, E., Hernandis, S., Sánchez-Carnero, N., Rodilla, M., Pérez-Arjona, I., Moszynski, M., Puig-Pons, V., Tena-Medialdea, J., & Espinosa, V. (2021). Vertical Configuration of a Side Scan Sonar for the Monitoring of Posidonia oceanica Meadows. Journal of Marine Science and Engineering, 9(12), 1332. https://doi.org/10.3390/jmse9121332