
The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.1.1. General Description of the Study Area
2.1.2. Mini-BRUV
2.1.3. Sampling Design
- Toothfish behaviour at the bait
- 2.
- Comparison between BRUV data and longline catch
- 3.
- Habitats and distribution of Antarctic toothfish individuals
2.2. Video Analysis
2.3. Abundance Indices
3. Results
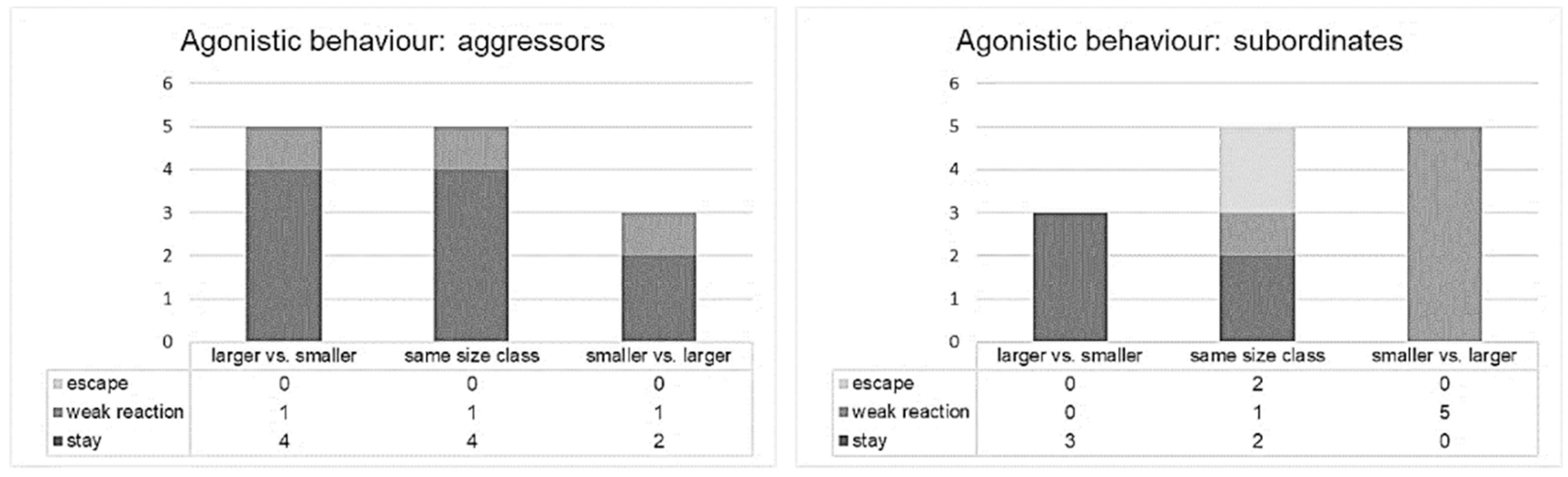
3.1. Fish Behaviour
3.2. BRUV Data and Longline Catch
3.3. Sea Bottom Features, Depth, and Toothfish Abundance
3.4. Other Fish Species Recorded by BRUV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Long, R.D.; Charles, A.; Stephenson, R.L. Key principles of marine ecosystem-based management. Mar. Policy 2015, 57, 53–60. [Google Scholar] [CrossRef]
- Mormede, S.; Dunn, A.; Hanchet, S.M. A stock assessment model of Antarctic toothfish (Dissostichus mawsoni) in the Ross Sea region incorporating multi-year mark-recapture data. CCAMLR Sci. 2014, 21, 39–62. [Google Scholar]
- Hanchet, S.M.; Dunn, A.; Parker, S.J.; Horn, P.L.; Stevens, D.W.; Mormede, S. The Antarctic toothfish (Dissostichus mawsoni): Biology, ecology, and life history in the Ross Sea region. Hydrobiologia 2015, 761, 397–414. [Google Scholar] [CrossRef]
- Hanchet, S.M.; Rickard, G.J.; Fenaughty, J.M.; Dunn, A.; Williams, M.J. A hypothetical life cycle for Antarctic toothfish Dissostichus mawsoni in Antarctic waters of CCAMLR Statistical Area 88. CCAMLR Sci. 2008, 15, 35–54. [Google Scholar]
- Parker, S.J.; Stevens, D.W.; Ghigliotti, L.; La Mesa, M.; Di Blasi, D.; Vacchi, M. Winter spawning of Antarctic toothfish Dissostichus mawsoni in the Ross Sea region. Antarctic Sci. 2019, 31, 243–253. [Google Scholar] [CrossRef]
- Eastman, J.T.; DeVries, A.L. Aspects of body size and gonadal histology in the Antarctic toothfish, Dissostichus mawsoni, from McMurdo Sound, Antarctica. Polar Biol. 2000, 23, 189–195. [Google Scholar] [CrossRef]
- Ramorino, M.C. Rapporto sulla Campagna Antartica Estate Australe 2003–2004; Diciannovesima spedizione. In PNRA Programma Nazionale di Ricerche in Antartide; Progetto Antartide, Final Report; Consortium for the Implementation of National Research Programmes in Antarctica: Roma, Italy, 2004; pp. 71–75. [Google Scholar]
- Ainley, D.G.; Nur, N.; Eastman, J.T.; Ballard, G.; Parkinson, C.L.; Evans, C.W.; DeVries, A.L. Decadal trends in abundance, size and condition of Antarctic toothfish in McMurdo Sound, Antarctica, 1972–2011. Fish Fish 2013, 14, 343–363. [Google Scholar] [CrossRef] [Green Version]
- Parker, S.J.; Mormede, S.; DeVries, A.; Hanchet, S.M.; Eisert, R. Have Antarctic toothfish returned to McMurdo Sound? Antarctic Sci. 2016, 28, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Parker, S.J.; Mormede, S.; Hanchet, S.M.; DeVries, A.; Canese, S.; Ghigliotti, L. Monitoring Antarctic toothfish in McMurdo Sound to evaluate the Ross Sea region Marine Protected Area. Antarctic Sci. 2019, 31, 195–207. [Google Scholar] [CrossRef]
- Mallet, D.; Pelletier, D. Underwater video techniques for observing coastal marine biodiversity: A review of sixty years of publications (1952–2012). Fish. Res. 2014, 154, 44–62. [Google Scholar] [CrossRef]
- Ono, K.; Kotwicki, S.; Dingsør, G.E.; Johnsen, E. Multispecies acoustic dead-zone correction and bias ratio estimates between acoustic and bottom-trawl data. ICES J. Mar. Sci. 2018, 75, 361–373. [Google Scholar] [CrossRef]
- O’Driscoll, R.L.; Ladroit, Y.; Parker, S.J.; Vacchi, M.; Canese, S.; Ghigliotti, L.; Dunford, A.J.; Mormede, S. Acoustic deployments reveal Antarctic silverfish under ice in the Ross Sea. Antarctic Sci. 2018, 30, 345–353. [Google Scholar] [CrossRef]
- Cappo, M.; Harvey, E.; Shortis, M. Counting and Measuring Fish with Baited Video Techniques-An Overview. In Australian Society for Fish Biology Workshop Proceedings; Australian Society for Fish Biology: Tasmania, Australian, 2006; Volume 1, pp. 101–114. [Google Scholar]
- Bassett, D.K.; Montgomery, J.C. Investigating nocturnal fish populations in situ using baited underwater video: With special reference to their olfactory capabilities. J. Exp. Mar. Biol. Ecol. 2011, 409, 194–199. [Google Scholar] [CrossRef]
- Ferrando, S.; Amaroli, A.; Gallus, L.; Di Blasi, D.; Carlig, E.; Rottigni, M.; Vacchi, M.; Parker, S.J.; Ghigliotti, L. Olfaction in the Antarctic toothfish Dissostichus mawsoni: Clues from the morphology and histology of the olfactory rosette and bulb. Polar Biol. 2019, 42, 1081–1091. [Google Scholar] [CrossRef]
- Carlig, E.; Di Blasi, D.; Ghigliotti, L.; Pisano, E.; Faimali, M.; O’Driscoll, R.; Parker, S.; Vacchi, M. Diversification of feeding structures in three adult Antarctic nototheniid fish. Polar Biol. 2018, 41, 1707–1715. [Google Scholar] [CrossRef]
- Bailey, D.M.; King, N.J.; Priede, I.G. Cameras and carcasses: Historical and current methods for using artificial food fall to study deepwater animals. Mar. Ecol. Prog. Ser. 2007, 350, 179–191. [Google Scholar] [CrossRef]
- Dunlop, K.M.; Marian Scott, E.; Parsons, D.; Bailey, D.M. Do agonistic behaviours bias baited remote underwater video surveys of fish? Mar. Ecol. 2015, 36, 810–818. [Google Scholar] [CrossRef]
- Ellis, D.M.; DeMartini, E.E. Evaluation of a video camera technique for indexing the abundance of juvenile pink snapper, Pristipomoides filamentosus, and other Hawaiian insular shelf fishes. Fish B-NOAA 1995, 93, 67–77. [Google Scholar]
- Whitmarsh, S.K.; Fairweather, P.G.; Huveneers, C. What is Big BRUVver up to? Methods and uses of baited underwater video. Rev. Fish Biol. Fisher. 2017, 27, 53–73. [Google Scholar] [CrossRef]
- Devine, B.M.; Wheeland, L.J.; Fisher, J.A. First estimates of Greenland shark (Somniosus microcephalus) local abundances in Arctic waters. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devine, B.M.; Wheeland, L.J.; de Moura Neves, B.; Fisher, J.A. Baited remote underwater video estimates of benthic fish and invertebrate diversity within the eastern Canadian Arctic. Polar Biol. 2019, 42, 1323–1341. [Google Scholar] [CrossRef]
- Yau, C.; Collins, M.A.; Bagely, P.M.; Everson, I.; Nolan, C.P.; Priede, I.G. Estimating the abundance of Patagonian toothfish Dissostichus eleginoides using baited cameras: A preliminary study. Fish. Res. 2001, 51, 403–408. [Google Scholar] [CrossRef]
- Smale, D.A.; Barnes, D.K.A.; Fraser, K.P.P.; Mann, P.J.; Brown, M.P. Scavenging in Antarctica: Intense variation between sites and seasons in shallow benthic necrophagy. J. Exp. Mar. Biol. Ecol. 2007, 349, 405–417. [Google Scholar] [CrossRef]
- Dunlop, K.M.; Barnes, D.K.A.; Bailey, D.M. Variation of scavenger richness and abundance between sites of high and low iceberg scour frequency in Rider Bay, west Antarcic Peninsula. Polar Biol. 2014, 37, 1741–1754. [Google Scholar] [CrossRef] [Green Version]
- Vanney, J.R.; Falconer, R.K.H.; Johnson, G.L. Geomorphology of the Ross Sea and adjacent oceanic provinces. Mar. Geol. 1981, 41, 73–102. [Google Scholar] [CrossRef]
- Langhorne, P.J.; Hughes, K.G.; Gough, A.J.; Smith, I.J.; Williams, M.J.M.; Robinson, N.J.; Stevens, C.L.; Rack, W.; Price, D.; Leonard, G.H.; et al. Observed platelet ice distributions in Antarctic sea ice: An index for ocean-ice shelf heat flux. Geophys. Res. Lett. 2015, 42, 5442–5451. [Google Scholar] [CrossRef] [Green Version]
- Hoppmann, M.; Richter, M.E.; Smith, I.J.; Jendersie, S.; Langhorne, P.J.; Thomas, D.N.; Dieckmann, G.S. Platelet ice: The Southern Ocean’s hidden ice: A review. Ann. Glaciol. 2020, 1–28. [Google Scholar] [CrossRef]
- Malandrino, M.; Abollino, O.; Buoso, S.; Casalino, C.E.; Gasparon, M.; Giacomino, A.; La Gioia, C.; Mentasti, E. Geochemical characterisation of Antarctic soils and lacustrine sediments from Terra Nova Bay. Microchem. J. 2009, 92, 21–31. [Google Scholar] [CrossRef]
- Brooks, E.J.; Sloman, K.A.; Sims, D.W.; Danylchuk, A.J. Validating the use of baited remote underwater video surveys for assessing the diversity, distribution and abundance of sharks in the Bahamas. Endanger. Species Res. 2011, 13, 231–243. [Google Scholar] [CrossRef]
- Barord, G.J.; Dooley, F.; Dunstan, A.; Ilano, A.; Keister, K.N.; Neumeister, H.; Preuss, T.; Schoepfer, S.; Ward, P.D. Comparative population assessments of Nautilus sp. in the Philippines, Australia, Fiji, and American Samoa using baited remote underwater video systems. PLoS ONE 2014, 9, e100799. [Google Scholar] [CrossRef] [Green Version]
- Stobart, B.; Díaz, D.; Álvarez, F.; Alonso, C.; Mallol, S.; Goñi, R. Performance of baited underwater video: Does it underestimate abundance at high population densities? PLoS ONE 2015, 10, e0127559. [Google Scholar] [CrossRef] [PubMed]
- Langlois, T.J.; Fitzpatrick, B.R.; Fairclough, D.V.; Wakefield, C.B.; Hesp, S.A.; McLean, D.L.; Harvey, E.S.; Meeuwig, J.J. Similarities between line fishing and baited stereo-video estimations of length-frequency: Novel application of kernel density estimates. PLoS ONE 2012, 7, e45973. [Google Scholar] [CrossRef]
- Lowry, M.; Folpp, H.; Gregson, M.; Suthers, I. Comparison of baited remote underwater video (BRUV) and underwater visual census (UVC) for assessment of artificial reefs in estuaries. J. Exp. Mar. Biol. Ecol. 2012, 416, 243–253. [Google Scholar] [CrossRef]
- White, J.; Simpfendorfer, C.A.; Tobin, A.J.; Heupel, M.R. Application of baited remote underwater video surveys to quantify spatial distribution of elasmobranchs at an ecosystem scale. J. Exp. Mar. Biol. Ecol. 2013, 448, 281–288. [Google Scholar] [CrossRef]
- Santana-Garcon, J.; Newman, S.J.; Harvey, E.S. Development and validation of a mid-water baited stereo-video technique for investigating pelagic fish assemblages. J. Exp. Mar. Biol. Ecol. 2014, 452, 82–90. [Google Scholar] [CrossRef]
- Udyawer, V.; Cappo, M.; Simpfendorfer, C.A.; Heupel, M.R.; Lukoschek, V. Distribution of sea snakes in the Great Barrier Reef Marine Park: Observations from 10 yrs of baited remote underwater video station (BRUVS) sampling. Coral Reefs 2014, 33, 777–791. [Google Scholar] [CrossRef]
- Schmid, K.; Reis-Filho, J.A.; Harvey, E.; Giarrizzo, T. Baited remote underwater video as a promising nondestructive tool to assess fish assemblages in clearwater Amazonian rivers: Testing the effect of bait and habitat type. Hydrobiologia 2017, 784, 93–109. [Google Scholar] [CrossRef]
- Willis, T.J.; Millar, R.B.; Babcock, R.C. Protection of exploited fish in temperate regions: High density and biomass of snapper Pagrus auratus (Sparidae) in northern New Zealand marine reserves. J. Appl. Ecol. 2003, 40, 214–227. [Google Scholar] [CrossRef]
- Cappo, M.; De’ath, G.; Speare, P. Inter-reef vertebrate communities of the Great Barrier Reef Marine Park determined by baited remote underwater video stations. Mar. Ecol. Prog. Ser. 2007, 350, 209–221. [Google Scholar] [CrossRef]
- Harvey, E.S.; Cappo, M.; Butler, J.J.; Hall, N.; Kendrick, G.A. Bait attraction affects the performance of remote underwater video stations in assessment of demersal fish community structure. Mar. Ecol. Prog. Ser. 2007, 350, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Colton, M.A.; Swearer, S.E. A comparison of two survey methods: Differences between underwater visual census and baited remote underwater video. Mar. Ecol. Prog. Ser. 2010, 400, 19–36. [Google Scholar] [CrossRef] [Green Version]
- Gladstone, W.; Lindfield, S.; Coleman, M.; Kelaher, B. Optimisation of baited remote underwater video sampling designs for estuarine fish assemblages. J. Exp. Mar. Biol. Ecol. 2012, 429, 28–35. [Google Scholar] [CrossRef]
- Campbell, M.D.; Pollack, A.G.; Gledhill, C.T.; Switzer, T.S.; DeVries, D.A. Comparison of relative abundance indices calculated from two methods of generating video count data. Fish. Res. 2015, 170, 125–133. [Google Scholar] [CrossRef]
- Cappo, M.; Harvey, E.; Malcolm, H.; Speare, P. Potential of Video Techniques to Monitor Diversity, Abundance and Size of Fish in Studies of Marine Protected Areas. In Aquatic Protected Areas. What Works Best and How Do We Know? Beumer, J.P., Grant, A., Smith, D.C., Eds.; Australian Society for Fish Biology: Saint Lucia, Australia, 2003; Volume 1, pp. 455–464. [Google Scholar]
- Cappo, M.; Speare, P.; De’ath, G. Comparison of baited remote underwater video stations (BRUVS) and prawn (shrimp) trawls for assessments of fish biodiversity in inter-reefal areas of the Great Barrier Reef Marine Park. J. Exp. Mar. Biol. Ecol. 2004, 302, 123–152. [Google Scholar] [CrossRef]
- Stoner, A.W.; Laurel, B.J.; Hurst, T.P. Using a baited camera to assess relative abundance of juvenile Pacific cod: Field and laboratory trials. J. Exp. Mar. Biol. Ecol. 2008, 354, 202–211. [Google Scholar] [CrossRef]
- Priede, I.G.; Merrett, N.R. Estimation of abundance of abyssal demersal fishes; a comparison of data from trawls and baited cameras. J. Fish Biol. 1996, 49, 207–216. [Google Scholar] [CrossRef]
- Roberts, J.; Xavier, J.C.; Agnew, D.J. The diet of toothfish species Dissostichus eleginoides and Dissostichus mawsoni with overlapping distributions. J. Fish Biol. 2011, 79, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.T.; Licht, S.; Haiat, K.S.; Bonney, J.; Allder, J.; Chaloux, N.; Shomberg, R.; Noyes, T.J. DEEPi: A miniaturized, robust, and economical camera and computer system for deep-sea exploration. Deep. Sea. Res. Pt I 2019, 153, 103136. [Google Scholar] [CrossRef]
- Wraith, J.; Lynch, T.; Minchinton, T.; Broad, A.; Davis, A. Bait type affects fish assemblages and feeding guilds observed at baited remote underwater video stations. Mar. Ecol. Prog. Ser. 2013, 477, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.E.; Griffin, R.A.; Januchowski-Hartley, S.R.; Unsworth, R.K.F. The influence of bait on remote video observations in shallow-water coastal environments associated with the North-Eastern Atlantic. PeerJ 2020, 8, e9744. [Google Scholar] [CrossRef]
- Eastman, J.T.; Barry, J.P. Underwater video observation of the Antarctic toothfish Dissostichus mawsoni (Perciformes: Nototheniidae) in the Ross Sea, Antarctica. Polar Biol. 2002, 25, 391–395. [Google Scholar] [CrossRef]
- Fuiman, L.; Davis, S.; Williams, T. Behavior of midwater fishes under the Antarctic ice: Observations by a predator. Mar. Biol. 2002, 140, 815–822. [Google Scholar] [CrossRef]
- Langlois, T.; Goetze, J.; Bond, T.; Monk, J.; Abesamis, R.A.; Asher, J.; Barrett, N.; Bernard, A.T.F.; Bouchet, P.J.; Birt, M.J.; et al. A field and video annotation guide for baited remote underwater stereo-video surveys of demersal fish assemblages. Methods Ecol. Evol. 2020, 11, 1401–1409. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station # | Fish Recorded | VLL Catch (Mean ± SD) |
|---|---|---|
| 20 | 5 | 13.0 ± 2.6 |
| 28 | 3 | 13.7 ± 5.5 |
| 29 | 0 | 6.3 ± 5.9 |
| Site # | Set Duration (hh:mm:ss) | Depth (m) | Substrate | Benthos | Geomorph. | CPUE | MeanN | TFA (min) |
|---|---|---|---|---|---|---|---|---|
| 01 | 01:19:38 | 258 | mixed | 80% | Ridge | 0.75 | 1 | 8.2 |
| 02 | 05:38:43 | 356 | soft | 40% | Ridge | 0.53 | 0.6 | 238.8 |
| 03 | 06:01:05 | 475 | mixed | 80% | Trench | 0 | 0 | - |
| 04 | 06:29:34 | 303 | mixed | 70% | Ridge | 0 | 0 | - |
| 05 | 01:59:25 | 179 | mixed | 90% | Ridge | 0 | 0 | - |
| 06 | 06:14:34 | 543 | soft | 10% | Trench | 1.92 | 1.4 | 60.3 |
| 07 | 05:27:00 | 390 | soft | 60% | Ridge | 0.37 | 0.4 | 167.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Blasi, D.; Canese, S.; Carlig, E.; Parker, S.J.; Pisano, E.; Vacchi, M.; Ghigliotti, L. The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice. J. Mar. Sci. Eng. 2021, 9, 255. https://doi.org/10.3390/jmse9030255
Di Blasi D, Canese S, Carlig E, Parker SJ, Pisano E, Vacchi M, Ghigliotti L. The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice. Journal of Marine Science and Engineering. 2021; 9(3):255. https://doi.org/10.3390/jmse9030255
Chicago/Turabian StyleDi Blasi, Davide, Simonepietro Canese, Erica Carlig, Steven J. Parker, Eva Pisano, Marino Vacchi, and Laura Ghigliotti. 2021. "The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice" Journal of Marine Science and Engineering 9, no. 3: 255. https://doi.org/10.3390/jmse9030255
APA StyleDi Blasi, D., Canese, S., Carlig, E., Parker, S. J., Pisano, E., Vacchi, M., & Ghigliotti, L. (2021). The Challenge to Observe Antarctic Toothfish (Dissostichus mawsoni) under Fast Ice. Journal of Marine Science and Engineering, 9(3), 255. https://doi.org/10.3390/jmse9030255