
Hiding in Fouling Communities: A Native Spider Crab Decorating with a Cryptogenic Bryozoan in a Mediterranean Marina




{kind=link}
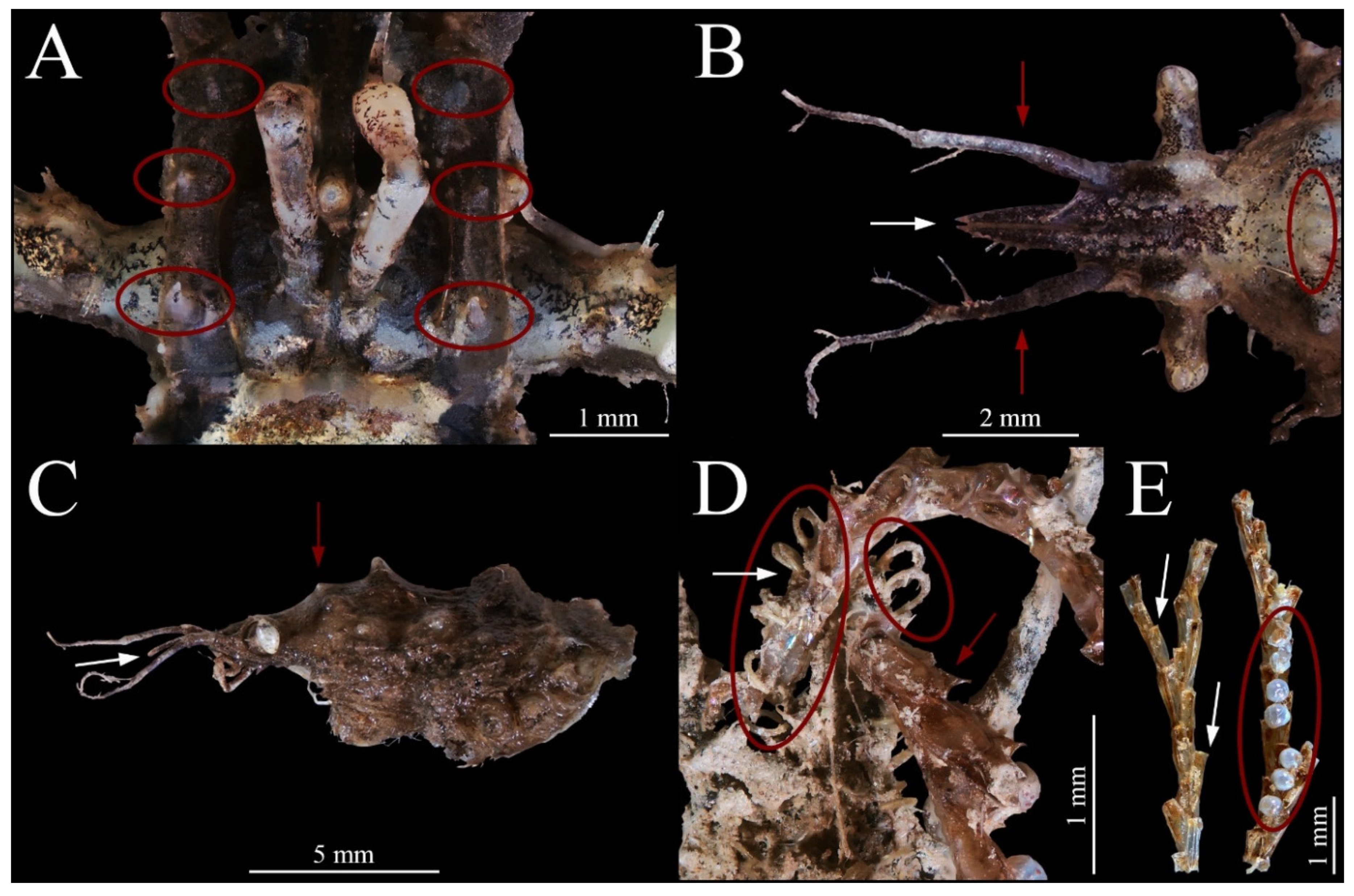{kind=link}
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wicksten, M.K. Camouflage in marine invertebrates. Oceanogr. Mar. Biol. Annu. Rev. 1983, 21, 177–193. [Google Scholar]
- Hultgren, K.; Stachowicz, J.J. Camouflage in decorator crabs: Integrating ecological, behavioural and evolutionary approaches. In Animal Camouflage Mechanisms and Function; Stevens, M., Merlaita, S., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 214–229. [Google Scholar]
- Ruxton, G.D.; Stevens, M. The evolutionary ecology of decorating behaviour. Biol. Lett. 2015, 11, 20150325. [Google Scholar] [CrossRef] [Green Version]
- Guinot, D.; Wicksten, M.K. Camouflage: Carrying behaviour, decoration behaviour, and other modalities of concealment in Brachyura. In Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea; Castro, P., Davie, P., Guinot, D., Schram, F., Klein, C.V., Eds.; Brill Online: Leiden, The Netherlands, 2015; Volume 9, pp. 583–638. [Google Scholar] [CrossRef]
- Stevens, M.; Merilaita, S. Animal Camouflage: Mechanisms and Function.; Cambridge University Press: Cambridge, UK, 2011; p. 357. [Google Scholar]
- Wicksten, M.K. A review and a model of decorating behavior in spider crabs (Decapoda, Brachyura, Majidae). Crustaceana 1993, 64, 314–325. [Google Scholar] [CrossRef]
- Woods, C.M.C.; McLay, C.L. Use of camouflage materials as a food store by the spider crab Notomithrax ursus (Brachyura: Majidae). N. Z. J. Mar. Freshw. Res. 1994, 28, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, J.J.; Hay, M.E. Reducing predation through chemically mediated camouflage: Indirect effect of plant defences on herbivores. Ecology 1999, 80, 2085–2101. [Google Scholar] [CrossRef]
- Thanh, P.D.; Wada, K.; Sato, M.; Shirayama, Y. Decorating behaviour by the majid crab Tiarinia cornigera as protection against predators. J. Mar. Biol. Assoc. UK 2003, 83, 1235–1237. [Google Scholar] [CrossRef]
- Hultgren, K.M.; Stachowicz, J.J. Alternative camouflage strategies mediate predation risk among closely related co-occurring kelp crabs. Oecologia 2008, 155, 519–528. [Google Scholar] [CrossRef]
- Hultgren, K.M.; Stachowicz, J.J. Evolution of decoration in majoid crabs: A comparative phylogenetic analysis of the role of body size and alternative defensive strategies. Am. Nat. 2009, 173, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Maluquer, P. Considerations on the fauna associated with colonies of Schizoporella errata of the Port of Mahon, Minorca, Balearic Islands, Spain. Publ. Dep. Zool. Univ. Barcelona 1985, 11, 23–28. [Google Scholar]
- Izquierdo-Muñoz, A.; Díaz Valdés, M.; Ramos-Esplá, A.A. Recent non-indigenous ascidians in the Mediterranean Sea. Aquat. Invasions 2009, 4, 59–64. [Google Scholar] [CrossRef] [Green Version]
- López-Legentil, S.; Legentil, M.L.; Erwin, P.M.; Turon, X. Harbor networks as introduction gateways: Contrasting distribution patterns of native and introduced ascidians. Biol. Invasions 2015, 17, 1623–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ros, M.; Vázquez-Luis, M.; Guerra-García, J.M. Environmental factors modulating the extent of impact in coastal invasions: The case of a widespread invasive caprellid (Crustacea: Amphipoda) in the Iberian Peninsula. Mar. Poll. Bull. 2015, 98, 247–258. [Google Scholar] [CrossRef]
- Leon, V.M.; Bellido, J.M. Mar Menor: Una laguna singular y sensible. Evaluación científica de su estado; Instituto Español de Oceanografía, Ministerio de Economía y Competitividad: Madrid, Spain, 2016; p. 414. [Google Scholar]
- Zariquey-Álvarez, R. Crustáceos Decápodos Ibéricos; Investigacion Pesquera: Barcelona, Spain, 1968; p. 510. [Google Scholar]
- Falciai, L.; Minervini, R. Guida dei Crostacei Decapodi d’Europa; Franco Muzzio Editore: Padova, Italy, 1992; p. 282. [Google Scholar]
- Forest, J.; Zariquey-Álvarez, R. Le genre Macropodia Leach en Méditerranée. I. Description et étude comparative des espèces (Crustacea Brachyura Majidae). Bull. Mus. Nat. Hist. Nat. Paris 1964, 36, 222–244. [Google Scholar]
- Van Noort, G.J.; Adema, J.P.H.M. The genus Macropodia Leach, 1814 in the North Sea and adjacent waters, with the description of a new species. Zool. Med. Leiden 1985, 59, 363–379. [Google Scholar]
- D’Udekem d’Acoz, C. A propos des trois Décapodes nouveaux pour la faune portugais: Philocheras monacanthus (Holthuis, 1961), Pachygrapsus transversus (Gibbes, 1950) et Macropodia zernjawskii (Brandt, 1880). Arq. Mus. Bocage 1992, 2, 127–136. [Google Scholar]
- Spiridonov, V.A.; Simakova, U.V.; Anosov, S.E.; Zalota, A.K.; Timofeev, V.A. Review of Macropodia in the Black Sea supported by molecular barcoding data; with the redescription of the type material, observations on ecology and epibiosis of Macropodia czernjawskii (Brandt, 1880) and notes on other Atlanto-Mediterranean species of Macropodia Leach, 1814 (Crustacea, Decapoda, Inachidae). Zoosyst. Evol. 2020, 96, 609–635. [Google Scholar] [CrossRef]
- Ryland, J.S.; Hayward, P.J. British Anascan Bryozoans. Synopses of the British Fauna (New Series); Academic Press: London, UK, 1977; p. 188. [Google Scholar]
- Occhipinti-Ambrogi, A.; Sacchi, C.F. Briozoi Lagunari; Consiglio Nazionale delle Ricerche: Roma, Italy, 1981; p. 146. [Google Scholar]
- Zabala, M.; Maluquer, P. Illustrated keys for the classification of Mediterranean Bryozoa; Ajuntament de Barcellona: Barcelona, Spain, 1988; p. 294. [Google Scholar]
- Chimenz Gusso, C.; Nicoletti, L.; Bondanese, C. Briozoi; Società Italiana di Biologia Marina: Genova, Italy, 2014; p. 336. [Google Scholar]
- Igić, L. Some species of Bryozoa from the Adriatic Sea and from Freshwaters, Which Are of Special Importance for Fouling Complex; Kotor Institute of Marine Biology: Kotor, Montenegro, 2007; p. 116. [Google Scholar]
- Ryland, J.S.; Bishop, J.D.D.; De Blauwe, H.; El Nagar, A.; Minchin, D.; Wood, C.A.; Yunnie, A.L.E. Alien species of Bugula (Bryozoa) along the Atlantic coasts of Europe. Aquat. Invasions 2011, 6, 17–31. [Google Scholar] [CrossRef]
- Ramalhosa, P.; Souto, J.; Canning-Clode, J. Diversity of Bugulidae (Bryozoa, Cheilostomata) colonizing artificial substrates in the Madeira Archipelago (NE Atlantic Ocean). Helgol. Mar. Res. 2017, 71, 1. [Google Scholar] [CrossRef] [Green Version]
- Koçak, F.; Bakal, I. Bugulidae Species along the Aegean Coast of Turkey. Thalassas 2019, 35, 663–673. [Google Scholar] [CrossRef]
- Davidson, S.K.; Haygood, M.G. Identification of sibling species of the bryozoan Bugula neritina that produce different anticancer bryostatins and harbor distinct strains of the bacterial symbiont “Candidatus Endobugula sertula”. Biol. Bull. 1999, 196, 273–280. [Google Scholar] [CrossRef] [PubMed]
- McGovern, T.M.; Hellberg, M.E. Cryptic species, cryptic endosymbionts, and geographical variation in chemical defences in the bryozoan Bugula neritina. Mol. Ecol. 2003, 12, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Mackie, J.A.; Keough, M.J.; Christidis, L. Invasion patterns inferred from cytochrome oxidase I sequences in three bryozoans, Bugula neritina, Watersipora subtorquata, and Watersipora arcuata. Mar. Biol. 2006, 149, 285–295. [Google Scholar] [CrossRef]
- Fehlauer-Ale, K.H.; Mackie, J.A.; Lim-Fong, G.E.; Ale, E.; Pie, M.R.; Waeschenbach, A. Cryptic species in the cosmopolitan Bugula neritina complex (Bryozoa, Cheilostomata). Zool. Scr. 2014, 43, 193–205. [Google Scholar] [CrossRef]
- Fehlauer-Ale, K.H.; Winston, J.E.; Tilbrook, K.J.; Nascimento, K.B.; Vieira, L.M. Identifying monophyletic groups within Bugula sensu lato (Bryozoa, Buguloidea). Zool. Scr. 2015, 44, 334–347. [Google Scholar] [CrossRef]
- Shen, X.; Tian, M.; Chu, K.H.; Wang, J.F.; Chen, S.; Liu, H.L.; Zhao, X.H.; Zhao, F.Q. Comparative mitogenomic analyses reveal cryptic diversity of the bryozoan Bugula neritina Linnaeus, 1758, in the Yellow Sea. Mar. Freshw. Res. 2016, 67, 1241–1252. [Google Scholar] [CrossRef]
- Crocetta, F.; Tanduo, V.; Osca, D.; Turolla, E. The Chinese mitten crab Eriocheir sinensis H. Milne Edwards, 1853 (Crustacea: Decapoda: Varunidae) reappears in the northern Adriatic Sea: Another intrusion attempt or the trace of an overlooked population? Mar. Poll. Bull. 2020, 156, 111221. [Google Scholar] [CrossRef] [PubMed]
- Schubart, C.D.; Huber, M.G.J. Genetic comparisons of German populations of the stone crayfish, Austropotamobius torrentium (Crustacea: Astacidae). Bull. Fr. Pêche Piscic. 2006, 380–381, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Tanduo, V.; Osca, D.; Crocetta, F. A bycatch surprise: Scyllarus subarctus Crosnier, 1970 (Decapoda: Achelata: Scyllaridae) in the Mediterranean Sea. J. Crust. Biol. 2021, 41, ruab010. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Graeffe, E. Biologische Notìzen über Seethìere der Adria. I. Ueber die bei den Oxyrhynchen vorkommende Maskirung. Boll. Soc. Adr. Sci. Nat. Trieste 1882, 7, 41–51. [Google Scholar]
- Aurivillius, C.W. Die Maskierung der oxyrhynchen Decapoden. K. Svenska Vetensk. Akad. Handl. 1889, 23, 1–72. [Google Scholar]
- Cruz-Rivera, E. Generality and specificity in the feeding and decoration preferences of three Mediterranean crabs. J. Exp. Mar. Biol. Ecol. 2001, 266, 17–31. [Google Scholar] [CrossRef]
- Furbock, S.; Patzner, R.A. Decoration preferences of Maja crispata Risso 1827 (Brachyura, Majidae). Nat. Croat. 2005, 14, 175. [Google Scholar]
- Martinelli, M.; Calcinai, B.; Bavestrello, G. Use of sponges in the decoration of Inachus phalangium (Decapoda, Majidae) from the Adriatic Sea. Ital. J. Zool. 2006, 73, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, M.; Betti, F.; Di Camillo, C.; Puce, S.; Bavestrello, G. Patterns of epibiont colonisation on the spider crab Inachus communissimus (Decapoda, Inachidae) from the Northern Adriatic Sea (Mediterranean Sea). Ital. J. Zool. 2011, 78, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Rorandelli, R.; Gomei, M.; Vannini, M.; Cannicci, S. Feeding and masking selection in Inachus phalangium (Decapoda, Majidae): Dressing up has never been so complicated. Mar. Ecol. Prog. Ser. 2007, 336, 225–233. [Google Scholar] [CrossRef]
- Woods, C.M.C. Masking in the spider crab Trichoplatus huttoni (Brachyura: Majidae). N. Z. Nat. Sci. 1995, 22, 75–80. [Google Scholar]
- Brooker, R.M.; Muñoz Ruiz, E.C.; Sih, T.L.; Dixson, D.L. Shelter availability mediates decorating in the majoid crab, Camposcia retusa. Behav. Ecol. 2018, 29, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Wicksten, M.K. Decorating behavior in Loxorhynchus crispatus Stimpson and Loxorhynchus grandis Stimpson (Brachyura, Majidae). Crustaceana 1979, 5, 461979. [Google Scholar]
- Wicksten, M.K. Decorator crabs. Sci. Am. 1980, 242, 116–122. [Google Scholar] [CrossRef]
- D’Udekem d’Acoz, C. Inventaire et Distribution des Crustacés Décapodes de L’Atlantique Nord-Oriental, de la Méditerranée et des Eaux Continentales Adjacentes au Nord de 25° N; Collection Patrimoines Naturels: Paris, France, 1999; p. 383. [Google Scholar]
- Melzer, R.R.; Caseña, F.; Buršić, M.; Lehmann, T.; Meyer, R.; Mavrič, B.; Makovec, T.; Pfannkuchen, M.; McHenry, J.; Heß, M. Knights, Ballerinas and Invisibles: The Decapod Crustaceans of the Brijuni Marine Protected Area; Javna ustanova Nacionalni park Brijuni: Pula, Croatia, 2019; p. 121. [Google Scholar]
- Skelhorn, J.; Rowland, H.M.; Speed, M.P.; Ruxton, G.D. Masquerade: Camouflage without crypsis. Science 2010, 327, 51. [Google Scholar] [CrossRef] [PubMed]
- Cutress, C.; Ross, D.M.; Sutton, L. The association of Calliactis tricolor with its pagurid, calappid, and majid partners in the Caribbean. Can. J. Zool. 1970, 48, 371–376. [Google Scholar] [CrossRef]
- Clynick, B.G. Assemblages of fish associated with coastal marinas in north-western Italy. J. Mar. Biol. Ass. UK 2006, 86, 847–852. [Google Scholar] [CrossRef]
- Clynick, B.G. Characteristics of an urban fish assemblage: Distribution of fish associated with coastal marinas. Mar. Environ. Res. 2008, 65, 18–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, K.B.; Ng, C.S.L.; Leong, W.G.K.; Jaafar, Z.; Chou, L.M. Assemblages and diversity of fishes in Singapore’s marinas. Raff. Bull. Zool. 2016, 32, 85–94. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanduo, V.; Virgili, R.; Osca, D.; Crocetta, F. Hiding in Fouling Communities: A Native Spider Crab Decorating with a Cryptogenic Bryozoan in a Mediterranean Marina. J. Mar. Sci. Eng. 2021, 9, 495. https://doi.org/10.3390/jmse9050495
Tanduo V, Virgili R, Osca D, Crocetta F. Hiding in Fouling Communities: A Native Spider Crab Decorating with a Cryptogenic Bryozoan in a Mediterranean Marina. Journal of Marine Science and Engineering. 2021; 9(5):495. https://doi.org/10.3390/jmse9050495
Chicago/Turabian StyleTanduo, Valentina, Riccardo Virgili, David Osca, and Fabio Crocetta. 2021. "Hiding in Fouling Communities: A Native Spider Crab Decorating with a Cryptogenic Bryozoan in a Mediterranean Marina" Journal of Marine Science and Engineering 9, no. 5: 495. https://doi.org/10.3390/jmse9050495
APA StyleTanduo, V., Virgili, R., Osca, D., & Crocetta, F. (2021). Hiding in Fouling Communities: A Native Spider Crab Decorating with a Cryptogenic Bryozoan in a Mediterranean Marina. Journal of Marine Science and Engineering, 9(5), 495. https://doi.org/10.3390/jmse9050495