Silk-Based Therapeutics Targeting Pseudomonas aeruginosa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
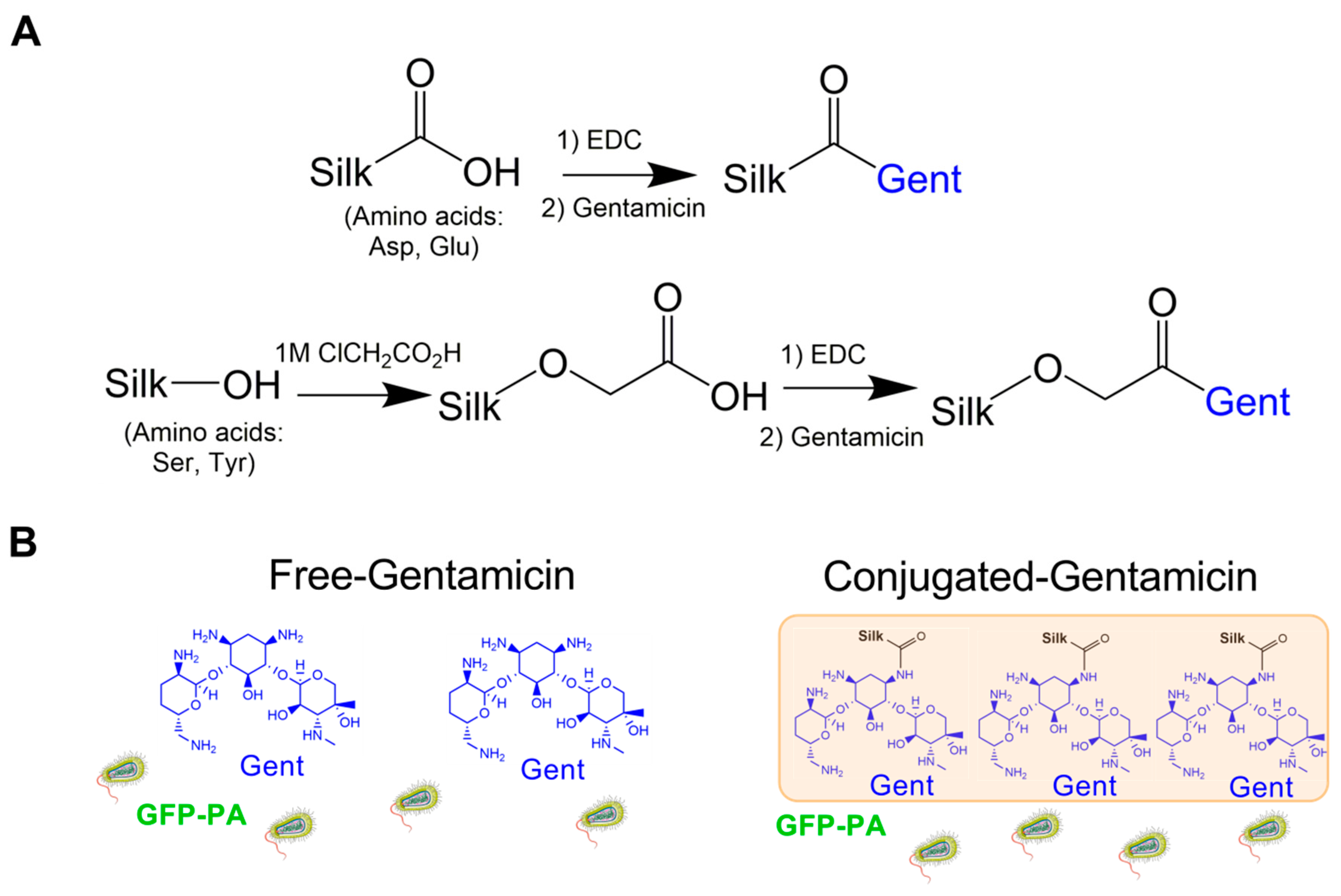
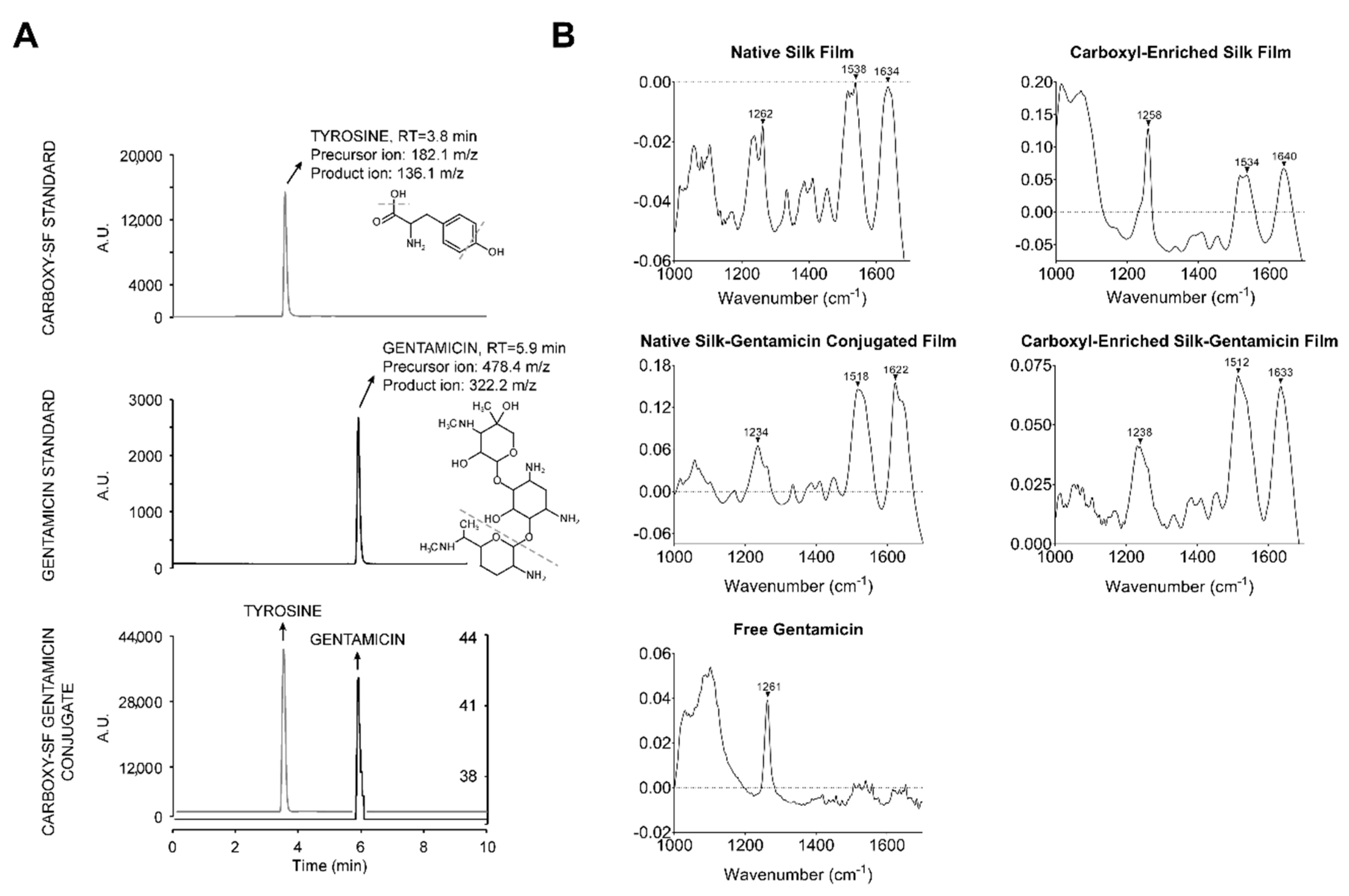
2.1. Design and Characterization of Gentamicin-Conjugated Silk Films
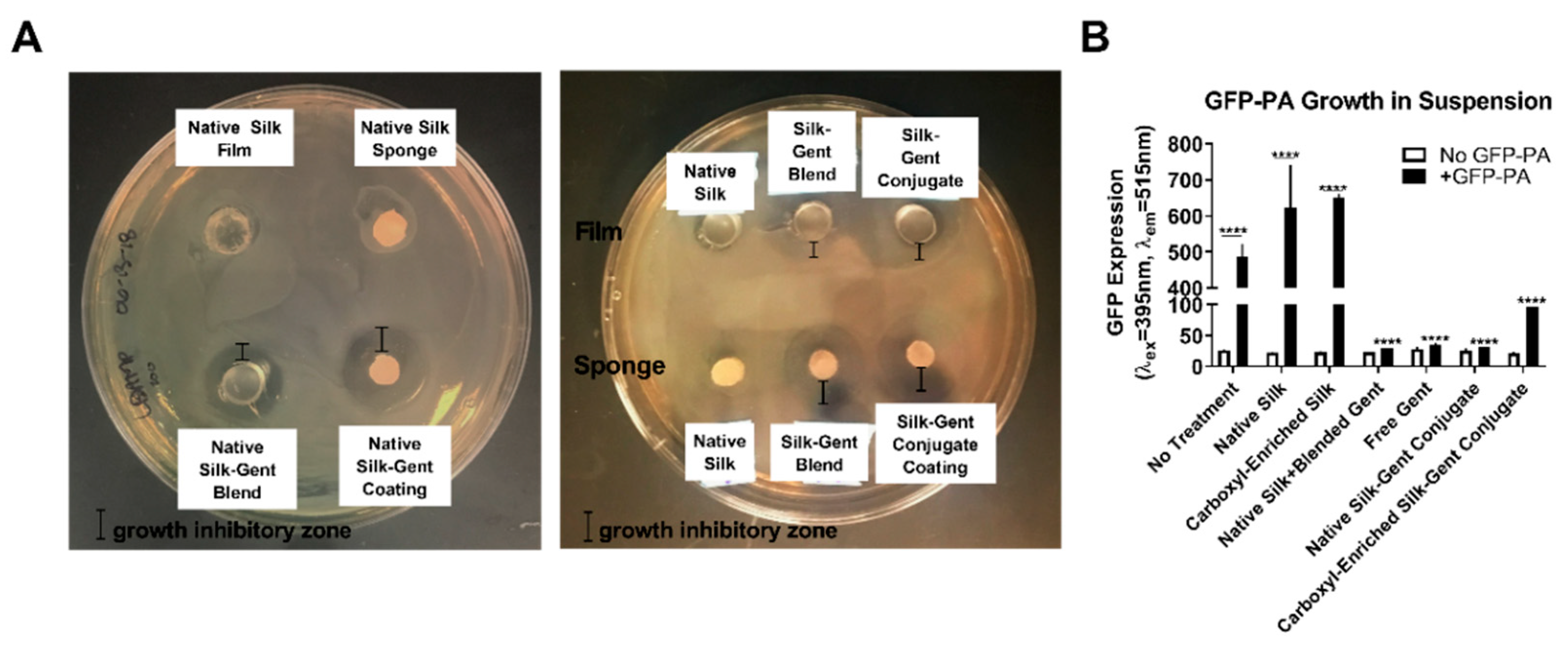
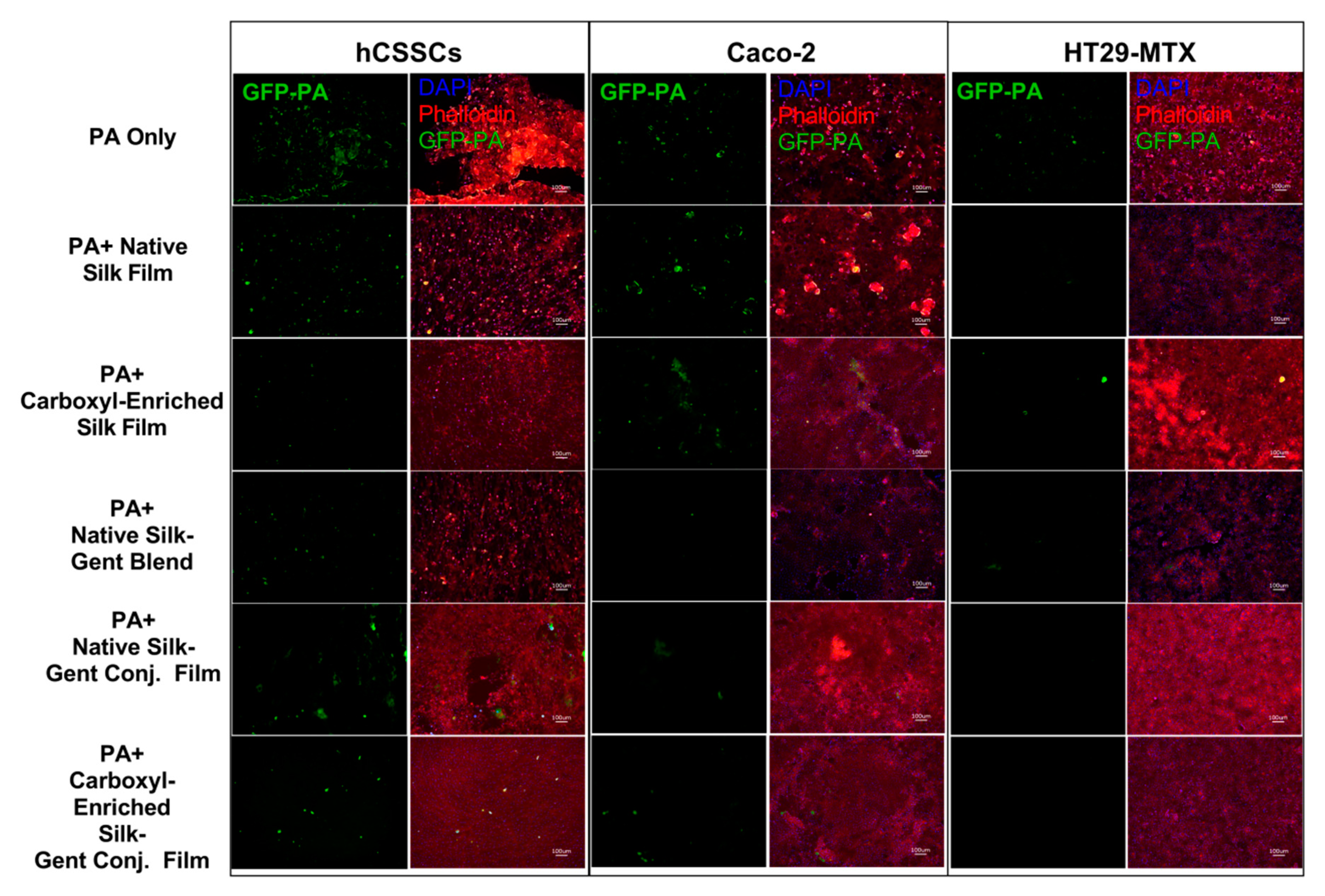
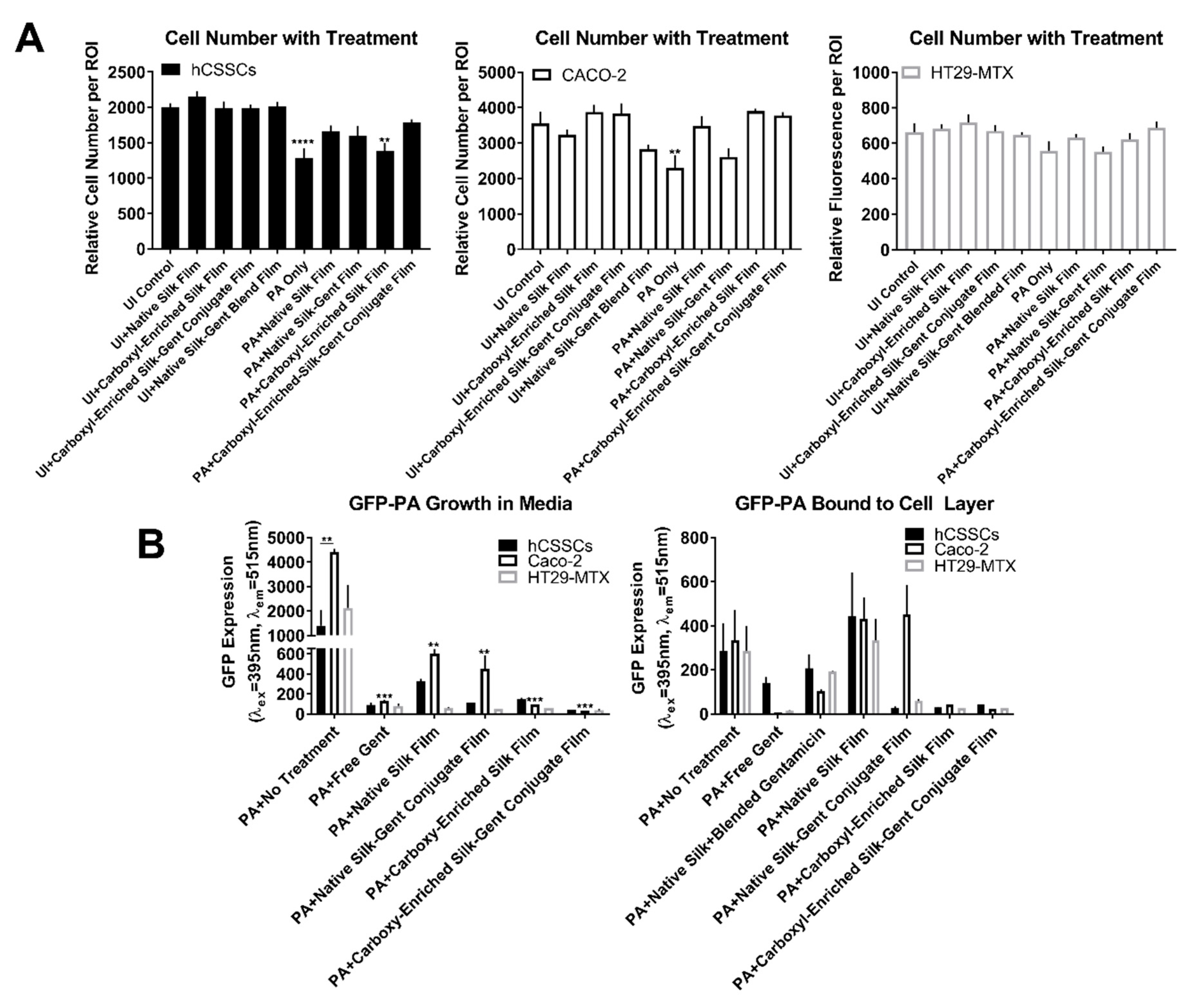
2.2. Efficacy of Gentamicin-Loaded Silk Biomaterials Against Bacterial Infection
3. Materials and Methods
3.1. PDMS Mold Preparation
3.2. Silk Film Preparation
3.3. Silk Sponge Preparation
3.4. Synthesis of Carboxyl-Enriched Silk
3.5. EDC-Mediated Gentamicin–Silk Conjugation
3.6. Infrared Spectroscopy
3.7. Mass Spectrometry
3.8. Ultraviolet (UV)/Visible Spectroscopy
3.9. Cell Culture
3.9.1. Pseudomonas aeruginosa
3.9.2. Bacterial Abundance
3.9.3. Corneal Stromal Cells
3.9.4. Caco-2 and HT29-MTX Cells
3.10. De-Cellularization of Corneal Tissue
3.11. Cell Counting
3.12. Viability Assay
3.13. Immunohistochemistry and Protein Staining
3.14. Fluroescence Microscopy
3.15. Scanning Electron Microscopy
3.16. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- CDC. Antibiotic resistance threats in the United States, 2013 Services, US Department of Health and Human Services, Ed. Available online: https://www.cdc.gov/drugresistance/pdf/ar-threats-2013-508.pdf (accessed on 23 April 2013).
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Nat. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Cantón, R.; Campo, P.; Baquero, F.; Blázquez, J. High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science 2000, 288, 1251–1253. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.D.; Rhinehart, E.; Jackson, M.; Chiarello, L. 2007 guideline for isolation precautions: Preventing transmission of infectious agents in health care settings. Am. J. Inf. Contr. 2007, 35, S65–S164. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, S.; Fourmy, D.; Puglisi, J.D. Structural origins of gentamicin antibiotic action. EMBO J. 1998, 17, 6437–6448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadurugamuwa, J.L.; Clarke, A.J.; Beveridge, T.J. Surface action of gentamicin on Pseudomonas aeruginosa. J. Bacteriol. 1993, 175, 5798–5805. [Google Scholar] [CrossRef] [PubMed]
- Ahangari, A.; Salouti, M.; Heidari, Z.; Kazemizadeh, A.R.; Safari, A.A. Development of gentamicin-gold nanospheres for antimicrobial drug delivery to staphylococcal infected foci. Drug Deliv. 2013, 20, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Burygin, G.; Khlebtsov, B.; Shantrokha, A.; Dykman, L.; Bogatyrev, V.; Khlebtsov, N. On the enhanced antibacterial activity of antibiotics mixed with gold nanoparticles. Nanoscale Res. Lett. 2009, 4, 794. [Google Scholar] [CrossRef]
- Weissleder, R.; Poss, K.; Wilkinson, R.; Zhou, C.; Bogdanov, A. Quantitation of slow drug release from an implantable and degradable gentamicin conjugate by in vivo magnetic resonance imaging. Antimicrob. Agents Chemother. 1995, 39, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Aumsuwan, N.; McConnell, M.S.; Urban, M.W. Tunable antimicrobial polypropylene surfaces: Simultaneous attachment of penicillin (gram+) and gentamicin (gram−). Biomacromolecules 2009, 10, 623–629. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Lam, J.S.; Beveridge, T.J. Interaction of gentamicin with the a band and b band lipopolysaccharides of Pseudomonas aeruginosa and its possible lethal effect. Antimicrob. Agents Chemother. 1993, 37, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Kotra, L.P.; Haddad, J.; Mobashery, S. Aminoglycosides: Perspectives on mechanisms of action and resistance and strategies to counter resistance. Antimicrob. Agents Chemother. 2000, 44, 3249–3256. [Google Scholar] [CrossRef] [PubMed]
- Antoci, V., Jr.; King, S.B.; Jose, B.; Parvizi, J.; Zeiger, A.R.; Wickstrom, E.; Freeman, T.A.; Composto, R.J.; Ducheyne, P.; Shapiro, I.M. Vancomycin covalently bonded to titanium alloy prevents bacterial colonization. J. Orthop. Res. 2007, 25, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Jose, B.; Antoci, V., Jr.; Zeiger, A.R.; Wickstrom, E.; Hickok, N.J. Vancomycin covalently bonded to titanium beads kills staphylococcus aureus. Chem. Biol. 2005, 12, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Lakes, A.L.; Peyyala, R.; Ebersole, J.L.; Puleo, D.A.; Hilt, J.Z.; Dziubla, T.D. Synthesis and characterization of an antibacterial hydrogel containing covalently bound vancomycin. Biomacromolecules 2014, 15, 3009–3018. [Google Scholar] [CrossRef]
- Costa, F.; Carvalho, I.F.; Montelaro, R.C.; Gomes, P.; Martins, M.C.L. Covalent immobilization of antimicrobial peptides (amps) onto biomaterial surfaces. Acta Biomater. 2011, 7, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Haynie, S.L.; Crum, G.A.; Doele, B.A. Antimicrobial activities of amphiphilic peptides covalently bonded to a water-insoluble resin. Antimicrob. Agents Chemother. 1995, 39, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Melo, M.N.; Ferre, R.; Castanho, M.A. Antimicrobial peptides: Linking partition, activity and high membrane-bound concentrations. Nature Rev. Microbiol. 2009, 7, 245. [Google Scholar] [CrossRef]
- Yucel, T.; Lovett, M.L.; Kaplan, D.L. Silk-based biomaterials for sustained drug delivery. J. Control. Release 2014, 190, 381–397. [Google Scholar] [CrossRef] [Green Version]
- Abbott, R.D.; Kimmerling, E.P.; Cairns, D.M.; Kaplan, D.L. Silk as a biomaterial to support long-term three-dimensional tissue cultures. ACS Appl. Mater. Interfaces 2016, 8, 21861–21868. [Google Scholar] [CrossRef]
- Pritchard, E.M.; Kaplan, D.L. Silk fibroin biomaterials for controlled release drug delivery. Expert Opin. Drug Deliv. 2011, 8, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Rockwood, D.N.; Preda, R.C.; Yucel, T.; Wang, X.; Lovett, M.L.; Kaplan, D.L. Materials fabrication from bombyx mori silk fibroin. Nature Protoc. 2011, 6, 1612–1631. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, C.E.; Wang, L.; Behlau, I.; Rnjak-Kovacina, J.; Wang, S.; Goldstein, M.H.; Liu, J.; Marchant, J.K.; Rosenblatt, M.I.; Kaplan, D.L. Degradation of silk films in multipocket corneal stromal rabbit models. J. Appl. Biomater. Funct. Mater. 2016, 14, e266–e276. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.D.; Marchant, J.K.; Pindrus, M.A.; Omenetto, F.G.; Kaplan, D.L. Silk film biomaterials for cornea tissue engineering. Biomaterials 2009, 30, 1299–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Rnjak-Kovacina, J.; Du, Y.; Funderburgh, M.L.; Kaplan, D.L.; Funderburgh, J.L. Corneal stromal bioequivalents secreted on patterned silk substrates. Biomaterials 2014, 35, 3744–3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, E.M.; Valentin, T.; Panilaitis, B.; Omenetto, F.; Kaplan, D.L. Antibiotic-releasing silk biomaterials for infection prevention and treatment. Adv. Funct. Mater. 2013, 23, 854–861. [Google Scholar] [CrossRef]
- Lovett, M.L.; Wang, X.; Yucel, T.; York, L.; Keirstead, M.; Haggerty, L.; Kaplan, D.L. Silk hydrogels for sustained ocular delivery of anti-vascular endothelial growth factor (anti-vegf) therapeutics. Eur. J. Pharm. Biopharm. 2015, 95, 271–278. [Google Scholar] [CrossRef]
- Conway, B.P.; Tabatabay, C.A.; Campochiaro, P.A.; D’amico, D.J.; Hanninen, L.A.; Kenyon, K.R. Gentamicin toxicity in the primate retina. Arch. Ophthalmol. 1989, 107, 107–112. [Google Scholar] [CrossRef]
- Halmagyi, G.M.; Fattore, C.M.; Curthoys, I.S.; Wade, S. Gentamicin vestibulotoxicity. Otolaryngol. Head Neck Surg. 1994, 111, 571–574. [Google Scholar] [CrossRef]
- Mesaros, N.; Nordmann, P.; Plésiat, P.; Roussel-Delvallez, M.; Van Eldere, J.; Glupczynski, Y.; Van Laethem, Y.; Jacobs, F.; Lebecque, P.; Malfroot, A. Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium. Clin. Microbiol. Infect. 2007, 13, 560–578. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Murphy, A.R.; Kaplan, D.L. Biomedical applications of chemically-modified silk fibroin. J. Mater. Chem. 2009, 19, 6443–6450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serban, M.A.; Kaplan, D.L. Ph sensitive ionomeric particles obtained via chemical conjugation of silk with poly-amino acids. Biomacromolecules 2010, 11, 3406–3412. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.; Coburn, J.M.; Partlow, B.P.; Mu, X.; Kaplan, D.L. Molecular and macro-scale analysis of enzyme-crosslinked silk hydrogels for rational biomaterial design. Acta Biomater. 2017, 63, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Löffler, D.; Ternes, T.A. Analytical method for the determination of the aminoglycoside gentamicin in hospital wastewater via liquid chromatography–electrospray-tandem mass spectrometry. J. Chromatogr. A 2003, 1000, 583–588. [Google Scholar] [CrossRef]
- Barbier, M.; Mittar, D. Novel Fluorescent Reporters for Studying Host-Pathogen Interactions; American Type Culture Collection (ATCC): Manassas, VA, USA, 2014. [Google Scholar]
- Hu, X.; Shmelev, K.; Sun, L.; Gil, E.S.; Park, S.H.; Cebe, P.; Kaplan, D.L. Regulation of silk material structure by temperature-controlled water vapor annealing. Biomacromolecules 2011, 12, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Shang, K.; Rnjak-Kovacina, J.; Lin, Y.; Hayden, R.S.; Tao, H.; Kaplan, D.L. Accelerated in vitro degradation of optically clear low beta-sheet silk films by enzyme-mediated pretreatment. Transl. Vis. Sci. Technol. 2013, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Rousset, M. The human colon carcinoma cell lines ht-29 and caco-2: Two in vitro models for the study of intestinal differentiation. Biochimie 1986, 68, 1035–1040. [Google Scholar] [CrossRef]
- Lesuffleur, T.; Porchet, N.; Aubert, J.-P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes muc1 to muc5 in relation to growth and differentiation of different mucus-secreting ht-29 cell subpopulations. J. Cell Sci. 1993, 106, 771–783. [Google Scholar]
- Gil, E.S.; Park, S.H.; Marchant, J.; Omenetto, F.; Kaplan, D.L. Response of human corneal fibroblasts on silk film surface patterns. Macromol. Biosci. 2010, 10, 664–673. [Google Scholar] [CrossRef]
- Basu, S.; Hertsenberg, A.J.; Funderburgh, M.L.; Burrow, M.K.; Mann, M.M.; Du, Y.; Lathrop, K.L.; Syed-Picard, F.N.; Adams, S.M.; Birk, D.E.; et al. Human limbal biopsy-derived stromal stem cells prevent corneal scarring. Sci. Transl. Med. 2014, 6, 266ra172. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nature Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; Sun, L.; Kim, H.J.; Hu, X.; Rice, W.; Kluge, J.; Reis, R.L.; Kaplan, D.L. Aligned silk-based 3-d architectures for contact guidance in tissue engineering. Acta Biomater. 2012, 8, 1530–1542. [Google Scholar] [CrossRef] [PubMed]
- Fleiszig, S.M.; Evans, D.J. Pathogenesis of contact lens-associated microbial keratitis. Optom. Vis. Sci. 2010, 87, 225–232. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKay, T.B.; Parker, R.N.; Hawker, M.J.; McGill, M.; Kaplan, D.L. Silk-Based Therapeutics Targeting Pseudomonas aeruginosa. J. Funct. Biomater. 2019, 10, 41. https://doi.org/10.3390/jfb10030041
McKay TB, Parker RN, Hawker MJ, McGill M, Kaplan DL. Silk-Based Therapeutics Targeting Pseudomonas aeruginosa. Journal of Functional Biomaterials. 2019; 10(3):41. https://doi.org/10.3390/jfb10030041
Chicago/Turabian StyleMcKay, Tina B., Rachael N. Parker, Morgan J. Hawker, Meghan McGill, and David L. Kaplan. 2019. "Silk-Based Therapeutics Targeting Pseudomonas aeruginosa" Journal of Functional Biomaterials 10, no. 3: 41. https://doi.org/10.3390/jfb10030041
APA StyleMcKay, T. B., Parker, R. N., Hawker, M. J., McGill, M., & Kaplan, D. L. (2019). Silk-Based Therapeutics Targeting Pseudomonas aeruginosa. Journal of Functional Biomaterials, 10(3), 41. https://doi.org/10.3390/jfb10030041