Cell-Penetrating Peptides as a Tool for the Cellular Uptake of a Genetically Modified Nitroreductase for use in Directed Enzyme Prodrug Therapy

 , ,
, ,
Abstract
:1. Introduction
2. Results
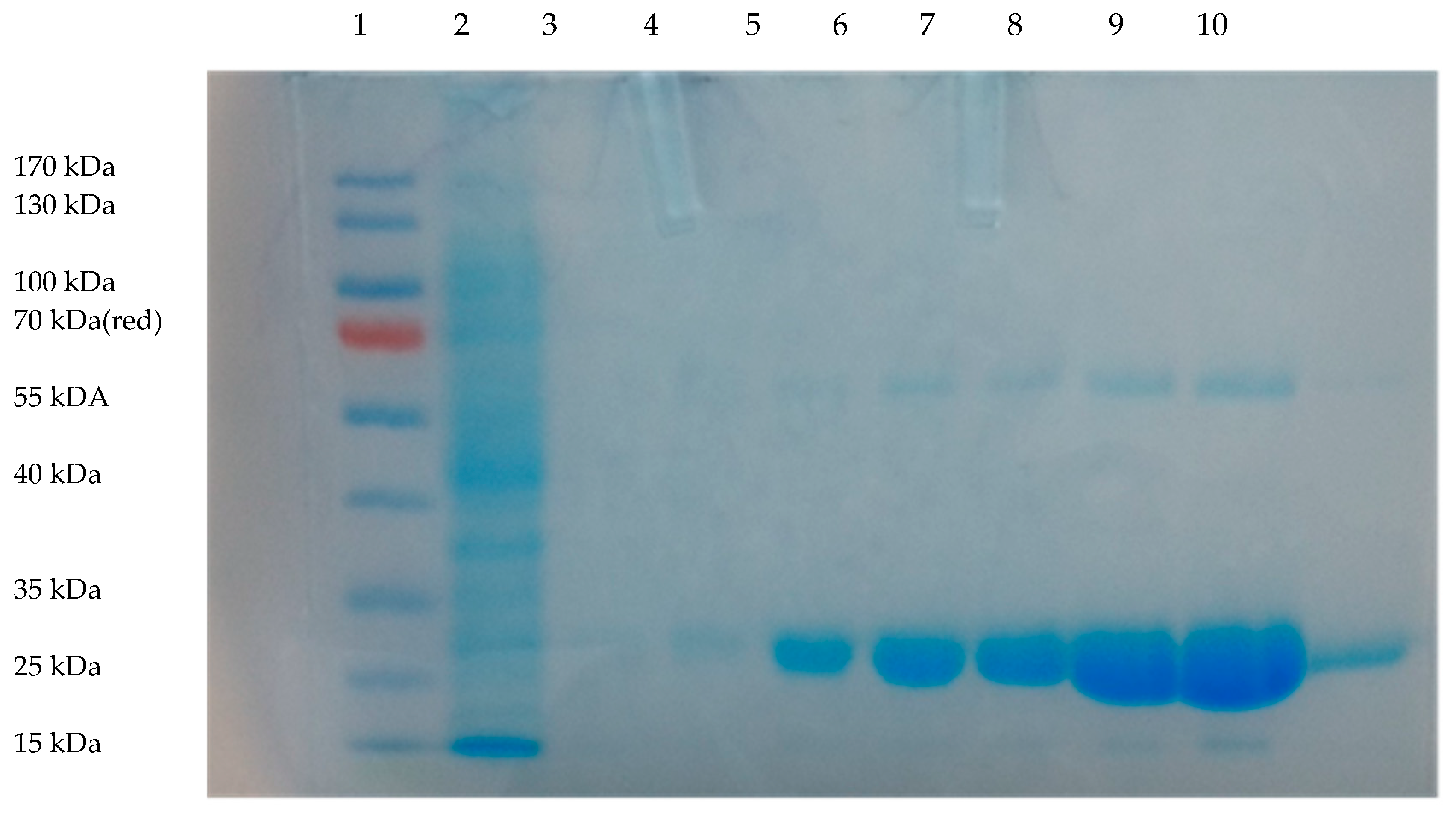
2.1. NfnB-Cys Expression and Purification
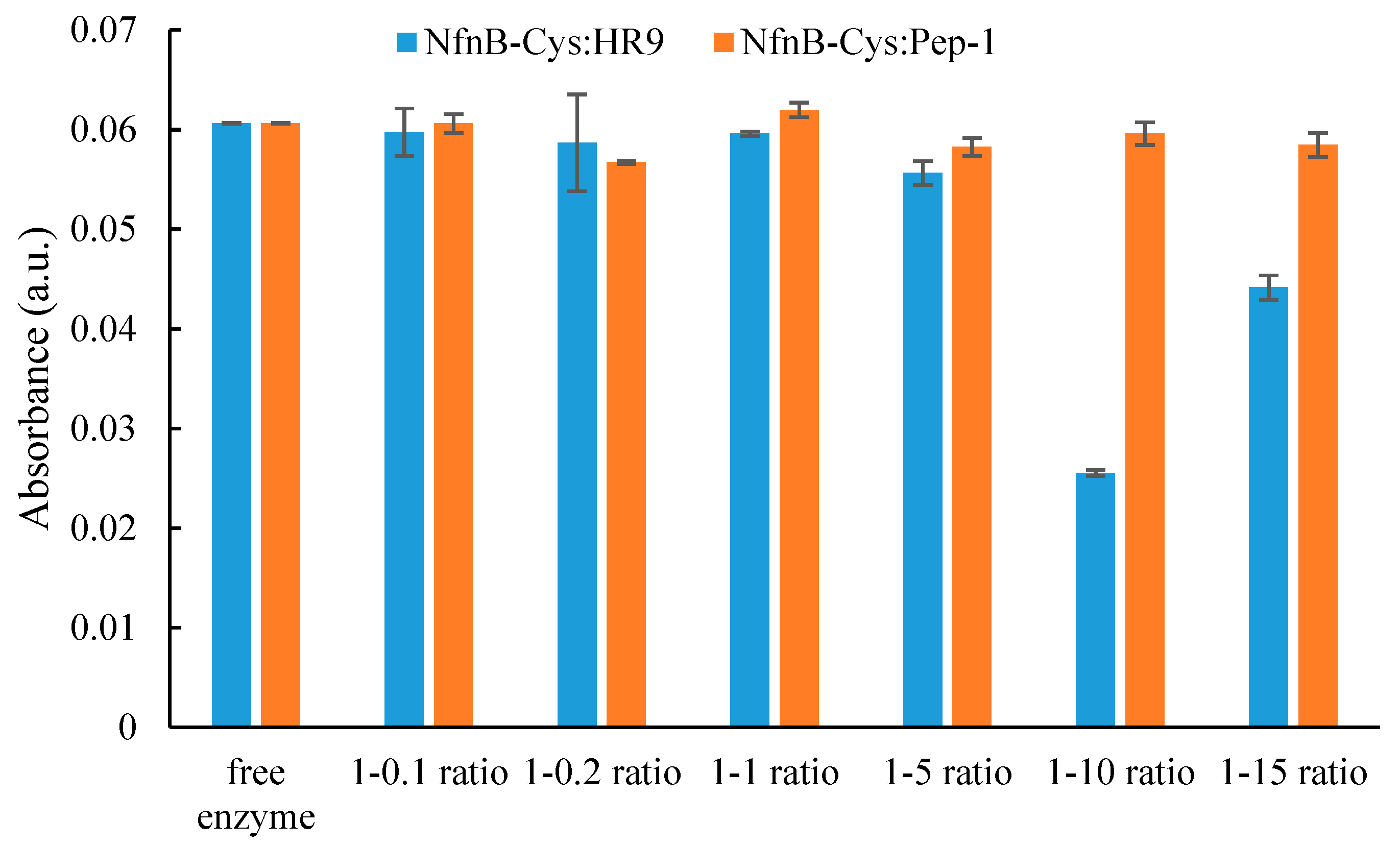
2.2. Conjugation of HR9 and Pep-1 to NfnB-Cys
2.3. NfnB-Cys and NfnB-Cys:CPP Conjugate Reactivity to CB1954
2.4. HPLC Profiles of NfnB-Cys and NfnB-Cyse:CPP Reaction Products
2.5. Effect of NfnB-Cys and NfnB-Cys:CPP Conjugate on Cell Viability
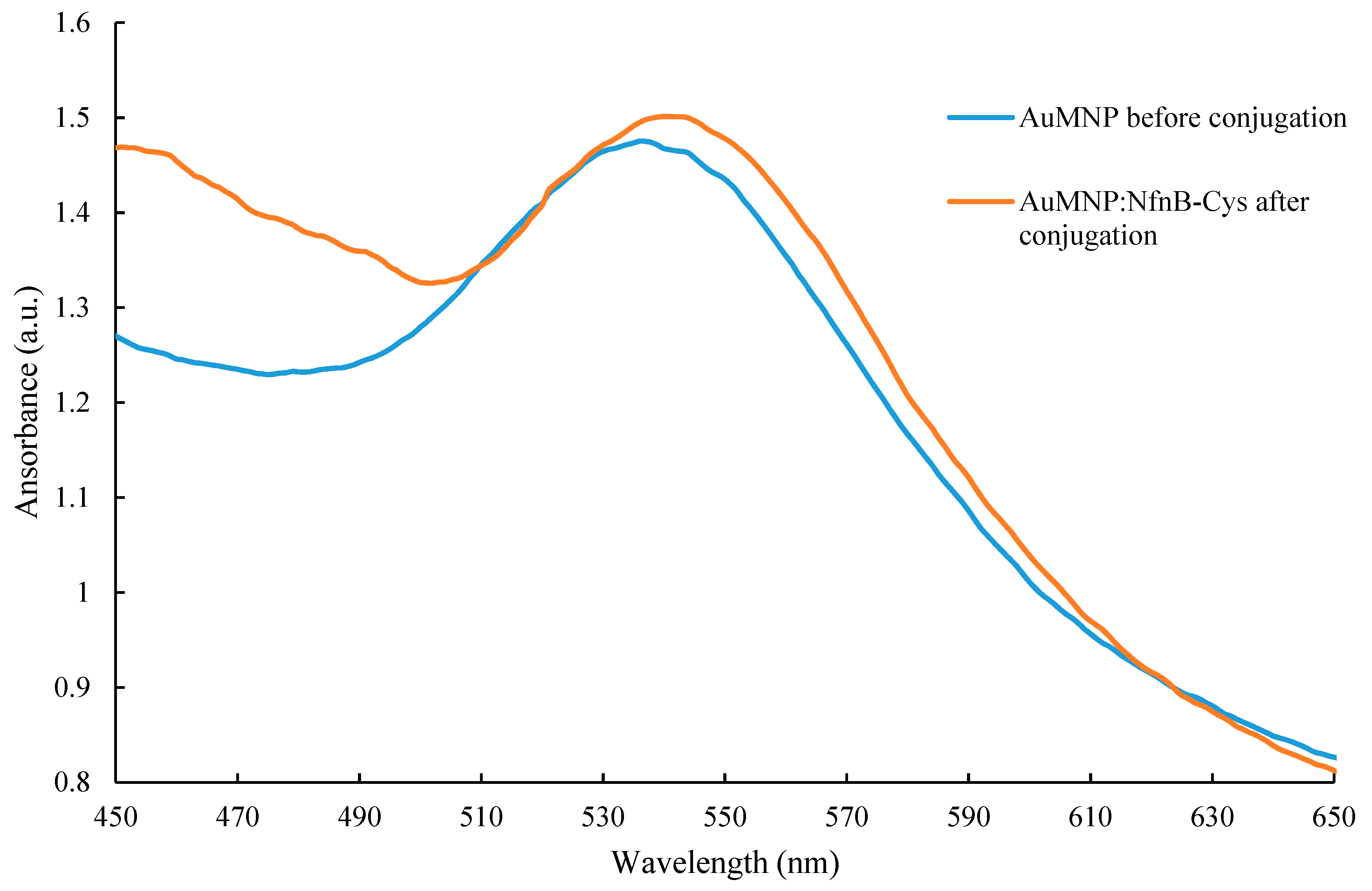
2.6. Effect of AuMNPs and AuMNP Conjugates on Cell Viability
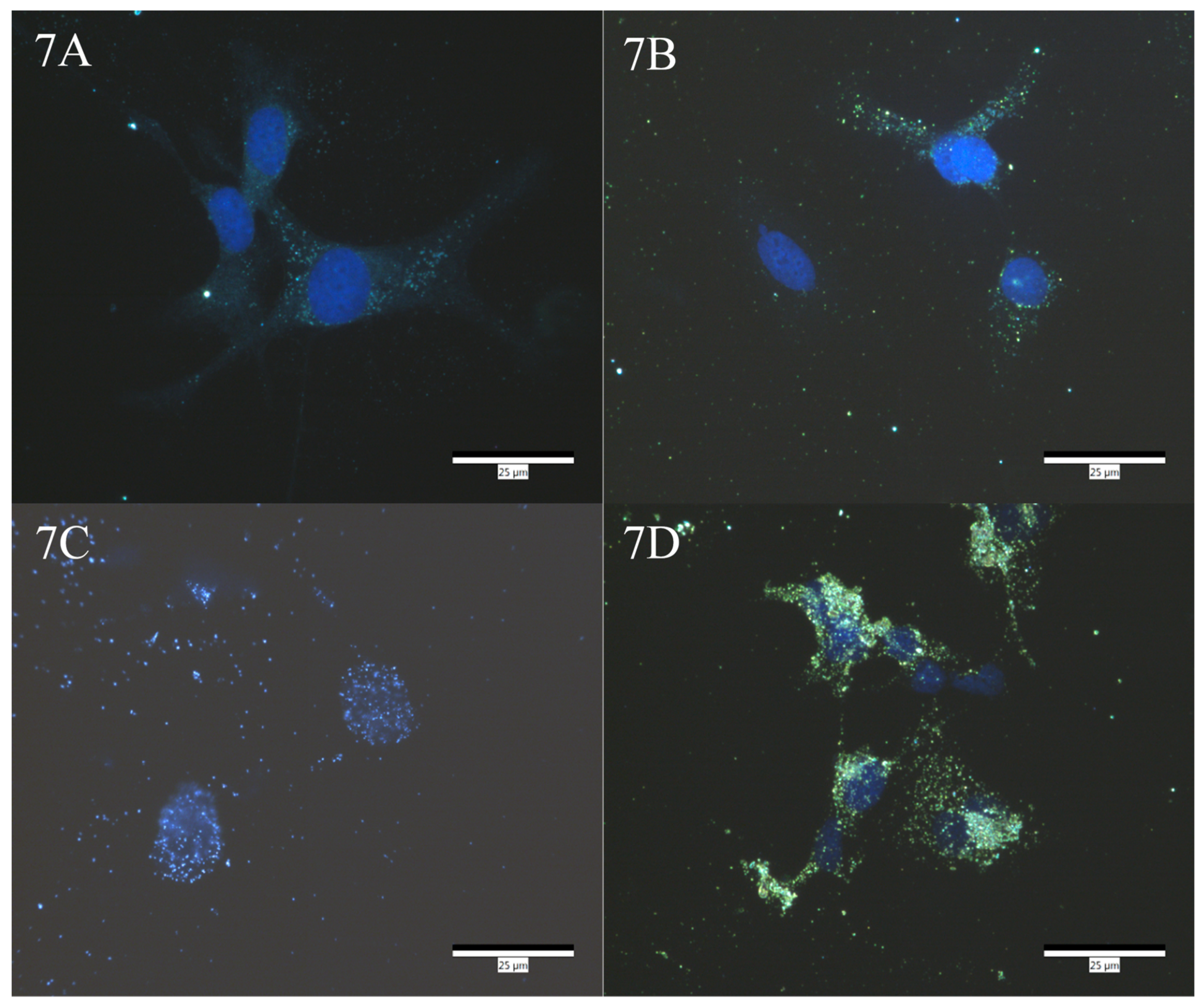
2.7. Darkfield Imaging
3. Discussion
4. Materials and Methods
4.1. Expression and Purification
4.2. CPP Conjugation to NfnB-Cys
4.3. Confirmation of CPP Conjugation to NfnB-Cys
4.4. NfnB-Cys and NfnB-Cys:CPP Conjugate Activity to CB1954
4.5. CB1954 Kinetics
4.6. HPLC
4.7. Cell Viability Assays
4.8. Preparation of AuMNP:NfnB-Cys and AuMNP:NfnB-Cys:HR9 Conjugates for Cell Culture and Darkfield Imaging
4.9. AuMNP Cell Viability Assays
4.10. Darkfield Imaging
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guo, P.; Coban, O.; Snead, N.M.; Trebley, J.; Hoeprich, S.; Guo, S.; Shu, Y. Engineering rna for targeted sirna delivery and medical application. Adv. Drug Deliv. Rev. 2010, 62, 650–666. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [PubMed]
- Luzio, J.P.; Poupon, V.; Lindsay, M.R.; Mullock, B.M.; Piper, R.C.; Pryor, P.R. Membrane dynamics and the biogenesis of lysosomes. Mol. Membr. Biol. 2003, 20, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Ghosh, R.N.; Maxfield, F.R. Endocytosis. Physiol. Rev. 1997, 77, 759–803. [Google Scholar] [CrossRef] [PubMed]
- Trabulo, S.; Cardoso, A.L.; Mano, M.; de Lima, M.C.P. Cell-penetrating peptides-mechanisms of cellular uptake and generation of delivery systems. Pharmaceuticals 2010, 3, 961–993. [Google Scholar] [CrossRef] [PubMed]
- Madani, F.; Lindberg, S.; Langel, Ü.; Futaki, S.; Gräslund, A. Mechanisms of cellular uptake of cell-penetrating peptides. J. Biophys. 2011, 2011, 1–10. [Google Scholar] [CrossRef]
- Langel, Ü. Handbook of Cell Penetrating Peptides, 2nd ed.; Taylor & Francis Group, LLC: Oxford, UK, 2006; Volume 28, ISBN 9780849350900. [Google Scholar]
- Rothbard, J.B.; Jessop, T.C.; Lewis, R.S.; Murray, B.A.; Wender, P.A. Role of Membrane Potential and Hydrogen Bonding in the Mechanism of Translocation of Guanidinium-Rich Peptides into Cells. J. Am. Chem. Soc. 2004, 126, 9506–9507. [Google Scholar] [CrossRef]
- Mitchell, D.J.; Steinman, L.; Kim, D.T.; Fathman, C.G.; Rothbard, J.B. Polyarginine enters cells more efficiently than other polycationic homopolymers. J. Pept. Res. 2000, 56, 318–325. [Google Scholar] [CrossRef]
- Järver, P.; Langel, Ü. Cell-penetrating peptides-A brief introduction. Biochim. Biophys. Acta Biomembr. 2006, 1758, 260–263. [Google Scholar] [CrossRef]
- Kato, T.; Yamashita, H.; Misawa, T.; Nishida, K.; Kurihara, M.; Tanaka, M.; Demizu, Y.; Oba, M. Plasmid DNA delivery by arginine-rich cell-penetrating peptides containing unnatural amino acids. Bioorg. Med. Chem. 2016, 24, 2681–2687. [Google Scholar] [CrossRef]
- Gautam, A.; Nanda, J.S.; Samuel, J.S.; Kumari, M.; Priyanka, P.; Bedi, G.; Nath, S.K.; Mittal, G.; Khatri, N.; Raghava, G.P.S. Topical Delivery of Protein and Peptide Using Novel Cell Penetrating Peptide IMT-P8. Sci. Rep. 2016, 6, 26278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, T.; Tsuzuku, T.; Takahashi, S.; Okamoto, A.; Dewa, T.; Nango, M.; Hyodo, K.; Ishihara, H.; Kikuchi, H.; Oku, N. Cell-penetrating peptide-conjugated lipid nanoparticles for siRNA delivery. Biochem. Biophys. Res. Commun. 2014, 444, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Gwenin, V.V.; Gwenin, C.D.; Kalaji, M. Colloidal gold modified with a genetically engineered nitroreductase: Toward a novel enzyme delivery system for cancer prodrug therapy. Langmuir 2011, 27, 14300–14307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kale, V.; Chen, M. Gene-Directed Enzyme Prodrug Therapy. AAPS J. 2015, 17, 102–110. [Google Scholar] [CrossRef]
- Knox, R.J.; Friedlos, F.; Boland, M.P. The bioactivation of CB 1954 and its use as a prodrug in antibody-directed enzyme prodrug therapy (ADEPT). Cancer Metastasis Rev. 1993, 12, 195–212. [Google Scholar] [CrossRef]
- Williams, E.M.; Little, R.F.; Mowday, A.M.; Rich, M.H.; Chan-Hyams, J.V.E.; Copp, J.N.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Nitroreductase gene-directed enzyme prodrug therapy: Insights and advances toward clinical utility. Biochem. J. 2015, 471, 131–153. [Google Scholar] [CrossRef]
- Race, P.R.; Lovering, A.L.; White, S.A.; Grove, J.I.; Searle, P.F.; Wrighton, C.W.; Hyde, E. Kinetic and Structural Characterisation of Escherichia coli Nitroreductase Mutants Showing Improved Efficacy for the Prodrug Substrate CB1954. J. Mol. Biol. 2007, 368, 481–492. [Google Scholar] [CrossRef]
- Knox, R.J.; Friedlos, F.; Sherwood, R.F.; Melton, R.G.; Anlezark, G.M. The bioactivation of 5-(aziridin-1-yl)-2,4-dinitrobenzamide (CB1954)-II. A comparison of an Escherichia coli nitroreductase and Walker DT diaphorase. Biochem. Pharmacol. 1992, 44, 2297–2301. [Google Scholar] [CrossRef]
- Helsby, N.A.; Ferry, D.M.; Patterson, A.V.; Pullen, S.M.; Wilson, W.R. 2-Amino metabolites are key mediators of CB 1954 and SN 23862 bystander effects in nitroreductase GDEPT. Br. J. Cancer 2004, 90, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Young, J.G.; Mautner, V.; Ashdown, D.; Bonney, S.; Pineda, R.G.; Collins, S.I.; Searle, P.F.; Hull, D.; Peers, E.; et al. A phase I/II clinical trial in localized prostate cancer of an adenovirus expressing nitroreductase with CB1984. Mol. Ther. 2009, 17, 1292–1299. [Google Scholar] [CrossRef]
- Schepelmann, S.; Springer, C.J. Viral vectors for gene-directed enzyme prodrug therapy. Curr. Gene Ther. 2006, 6, 647–670. [Google Scholar] [CrossRef] [PubMed]
- Palmer, D.H.; Mautner, V.; Mirza, D.; Oliff, S.; Gerritsen, W.; Van der Sijp, J.R.M.; Hubscher, S.; Reynolds, G.; Bonney, S.; Rajaratnam, R.; et al. Virus-directed enzyme prodrug therapy: Intratumoral administration of a replication-deficient adenovirus encoding nitroreductase to patients with resectable liver cancer. J. Clin. Oncol. 2004, 22, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Bagshawe, K.D. Antibody-Directed Enzyme Prodrug Therapy Professor. In Prodrugs: Challenges and rewards; Valentino, S., Borchardt, R., Hageman, M., Oliyai, R., Maag, H., Jefferson, T.W., Eds.; Springer: New York, NY, USA, 2005; pp. 526–536. ISBN 978-0-387-49785-3. [Google Scholar]
- Wadhwa, M.S.; Collard, W.T.; Adami, R.C.; McKenzie, D.L.; Rice, K.G. Peptide-mediated gene delivery: Influence of peptide structure on gene expression. Bioconjug. Chem. 1997, 8, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Behr, J.P. Gene Transfer with Synthetic Cationic Amphiphiles: Prospects for Gene Therapy. Bioconjug. Chem. 1994, 5, 382–389. [Google Scholar] [CrossRef]
- Gwenin, V.V.; Gwenin, C.D.; Kalaji, M. Drug Activation Carrier. European Union Patent EP2473197-A2, 2009. [Google Scholar]
- Gwenin, C.D.; Kalaji, M.; Williams, P.A.; Jones, R.M. The orientationally controlled assembly of genetically modified enzymes in an amperometric biosensor. Biosens. Bioelectron. 2007, 22, 2869–2875. [Google Scholar] [CrossRef]
- Ball, P.; Thompson, E.; Anderson, S.; Gwenin, V.; Gwenin, C. Time dependent HPLC analysis of the product ratio of enzymatically reduced prodrug CB1954 by a modified and immobilised nitroreductase. Eur. J. Pharm. Sci. 2019, 127, 217–224. [Google Scholar] [CrossRef]
- Liu, B.R.; Liou, J.S.; Chen, Y.J.; Huang, Y.W.; Lee, H.J. Delivery of Nucleic Acids, Proteins, and Nanoparticles by Arginine-Rich Cell-Penetrating Peptides in Rotifers. Mar. Biotechnol. 2013, 15, 584–595. [Google Scholar] [CrossRef]
- Deshayes, S.; Heitz, A.; Morris, M.C.; Charnet, P.; Divita, G.; Heitz, F. Insight into the Mechanism of Internalization of the Cell-Penetrating Carrier Peptide Pep-1 through Conformational Analysis. Biochemistry 2004, 43, 1449–1457. [Google Scholar] [CrossRef]
- Wang, M.H.; Otteson, D.C. Intracellular Delivery of Proteins into Mouse Muller Glia Cells in vitro and in vivo Using Pep-1 Transfection Reagent. J. Neurosci. 2010, 177, 403–419. [Google Scholar] [CrossRef]
- Gros, E.; Deshayes, S.; Morris, M.C.; Aldrian-Herrada, G.; Depollier, J.; Heitz, F.; Divita, G. A non-covalent peptide-based strategy for protein and peptide nucleic acid transduction. Biochim. Biophys. Acta Biomembr. 2006, 1758, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Koshman, Y.E.; Waters, S.B.; Walker, L.A.; Los, T.; de Tombe, P.; Goldspink, P.H.; Russell, B. Delivery and visualization of proteins conjugated to quantum dots in cardiac myocytes. J. Mol. Cell. Cardiol. 2008, 45, 853–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurzawa, L.; Pellerano, M.; Morris, M.C. PEP and CADY-mediated delivery of fluorescent peptides and proteins into living cells. Biochim. Biophys. Acta Biomembr. 2010, 1798, 2274–2285. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.R.; Huang, Y.W.; Winiarz, J.G.; Chiang, H.J.; Lee, H.J. Intracellular delivery of quantum dots mediated by a histidine- and arginine-rich HR9 cell-penetrating peptide through the direct membrane translocation mechanism. Biomaterials 2011, 32, 3520–3537. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.R.; Der Lin, M.; Chiang, H.J.; Lee, H.J. Arginine-rich cell-penetrating peptides deliver gene into living human cells. Gene 2012, 505, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, J.; Xu, D. Cell-penetrating peptides as noninvasive transmembrane vectors for the development of novel multifunctional drug-delivery systems. J. Control. Release 2016, 229, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwenin, V.V.; Poornima, P.; Halliwell, J.; Ball, P.; Robinson, G.; Gwenin, C.D. Identification of novel nitroreductases from Bacillus cereus and their interaction with the CB1954 prodrug. Biochem. Pharmacol. 2015, 98, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habtemariam, S. Catechols and quercetin reduce MTT through iron ions: A possible artefact in cell viability assays. Phyther. Res. 1995, 9, 603–605. [Google Scholar] [CrossRef]
- Lupu, A.R.; Popescu, T. The noncellular reduction of MTT tetrazolium salt by TiO2 nanoparticles and its implications for cytotoxicity assays. Toxicol. Vitr. 2013, 27, 1445–1450. [Google Scholar] [CrossRef]
- Morris, M.C.; Depollier, J.; Mery, J.; Heitz, F.; Divita, G. A peptide carrier for the delivery of biologically active proteins into mammalian cells. Nat. Biotechnol. 2001, 19, 1173–1176. [Google Scholar] [CrossRef]
- Morris, M.C.; Gros, E.; Aldrian-Herrada, G.; Choob, M.; Archdeacon, J.; Heitz, F.; Divita, G. A non-covalent peptide-based carrier for in vivo delivery of DNA mimics. Nucleic Acids Res. 2007, 35, 2191–2198. [Google Scholar] [CrossRef]
- Morris, M.C.; Deshayes, S.; Heitz, F.; Divita, G. Cell-penetrating peptides: From molecular mechanisms to therapeutics. Biol. Cell 2008, 100, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Arndt, C.; Koristka, S.; Bartsch, H.; Bachmann, M. Native Polyacrylamide Gels. Protein Electrophor. Methods Protoc. 2012, 869, 287–304. [Google Scholar]
- Liu, B.R.; Li, J.-F.; Lu, S.-W.; Leel, H.-J.; Huang, Y.-W.; Shannon, K.B.; Aronstam, R.S. Cellular internalization of quantum dots noncovalently conjugated with arginine-rich cell-penetrating peptides. J. Nanosci. Nanotechnol. 2010, 10, 6534–6543. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Hormesis defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- LeCher, J.C.; Nowak, S.J.; McMurry, J.L. Breaking in and busting out: Cell-penetrating peptides and the endosomal escape problem. Biomol. Concepts 2017, 8, 131–141. [Google Scholar] [CrossRef]
- Bio-Rad Buffer Formulations. Available online: http://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6199.pdf (accessed on 1 September 2019).
- Vass, S.O.; Jarrom, D.; Wilson, W.R.; Hyde, E.I.; Searle, P.F.E. coli NfsA: An alternative nitroreductase for prodrug activation gene therapy in combination with CB1954. Br. J. Cancer 2009, 100, 1903–1911. [Google Scholar] [CrossRef]
- Swe, P.M.; Copp, J.N.; Green, L.K.; Guise, C.P.; Mowday, A.M.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Targeted mutagenesis of the Vibrio fischeri flavin reductase FRase i to improve activation of the anticancer prodrug CB1954. Biochem. Pharmacol. 2012, 84, 775–783. [Google Scholar] [CrossRef]
- Prosser, G.A.; Patterson, A.V.; Ackerley, D.F. UvrB gene deletion enhances SOS chromotest sensitivity for nitroreductases that preferentially generate the 4-hydroxylamine metabolite of the anti-cancer prodrug CB1954. J. Biotechnol. 2010, 150, 190–194. [Google Scholar] [CrossRef]
- Prosser, G.A.; Copp, J.N.; Syddall, S.P.; Williams, E.M.; Smaill, J.B.; Wilson, W.R.; Patterson, A.V.; Ackerley, D.F. Discovery and evaluation of Escherichia coli nitroreductases that activate the anti-cancer prodrug CB1954. Biochem. Pharmacol. 2010, 79, 678–687. [Google Scholar] [CrossRef]
- Cude, M.P.; Gwenin, C.D. Development of Gold Coated Superparamagnetic Iron Oxide Nanoparticles for Nitroreductase Delivery. ECS Trans. 2011, 33, 79–89. [Google Scholar]
- Haiss, W.; Thanh, N.T.K.; Aveyard, J.; Fernig, D.G. Determination of Size and Concentration of Gold Nanoparticles from UV—Vis Spectra Determination of Size and Concentration of Gold Nanoparticles from UV—Vis Spectra. Anal. Chem 2007, 79, 4215–4221. [Google Scholar] [CrossRef] [PubMed]
- Kocbek, P.; Cegnar, M.; Kos, J.; Kristl, J. Targeting cancer cells using PLGA nanoparticles surface modified with monoclonal antibody. J. Control. Release 2007, 120, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; You, C.; Kim, B.; Turingan, R.S.; Forbes, N.S.; Martin, C.T.; Rotello, V.M. Light-Regulated Release of DNA and Its Delivery to Nuclei by Means of Photolabile Gold Nanoparticles **. Angew. Chem. Int. Ed. 2006, 45, 3165–3169. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Cheng, J.; Lin, C.; Wah, Y.; Han, S.; Wong, W. Nuclear penetration of surface functionalized gold nanoparticles. Toxicol. Appl. Pharmacol. 2009, 237, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, A.; Qiao, Y.; Zhang, P.; Ma, L.; Su, M. Cation surface modification of gold nanoparticles for enhanced cellular uptake and X-Ray radiation therapy. J. Mater. Chem. B 2015, 3, 7372–7376. [Google Scholar] [CrossRef]
- Arnida, M.A.; Ghanderhari, H. Cellular uptake and toxicity of gold nanoparticles in prostate cancer cells: A comparative study of rods and spheres. J. Appl. Toxicol. 2010, 30, 212–217. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
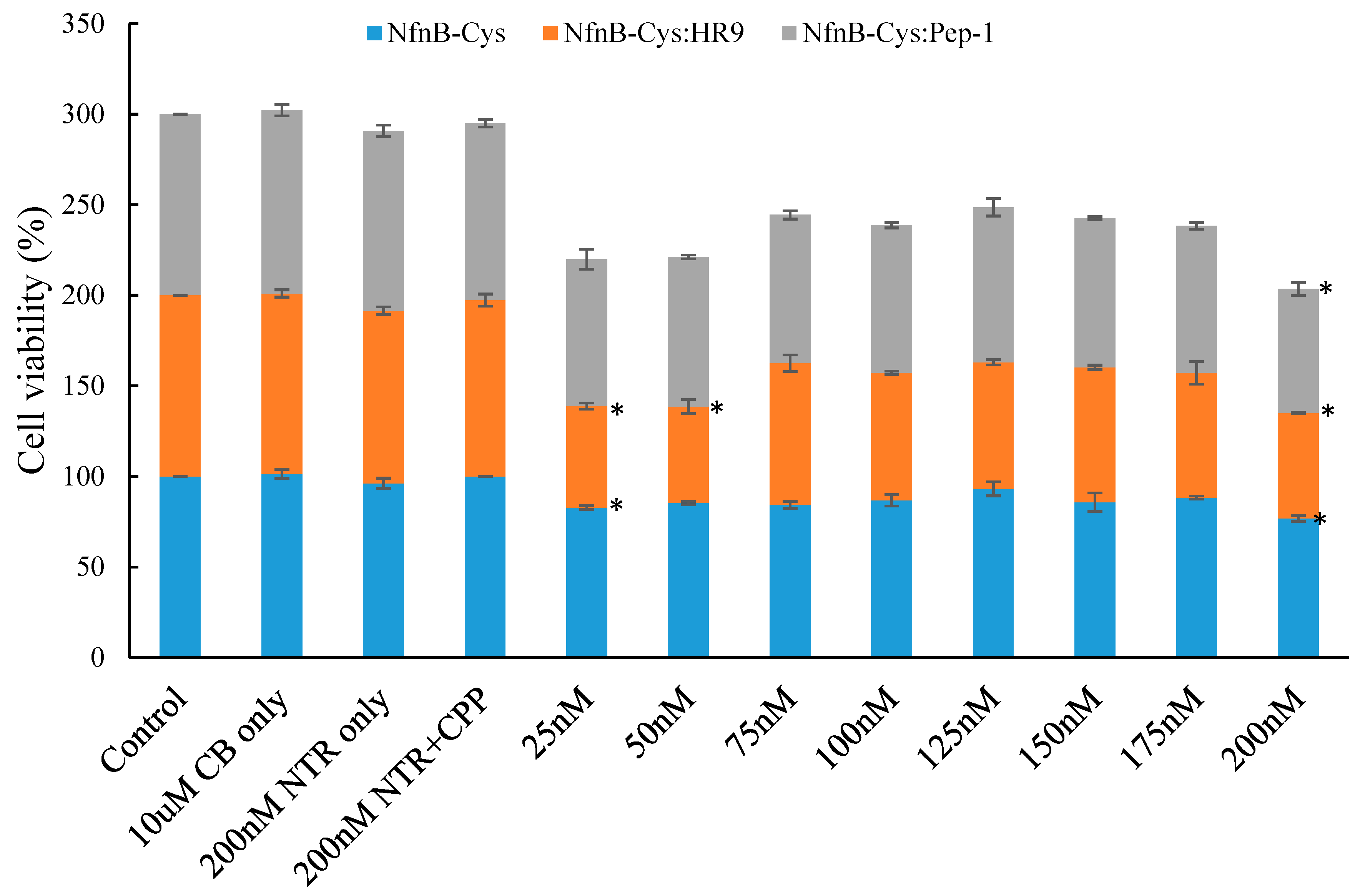{kind=link}
{kind=link}
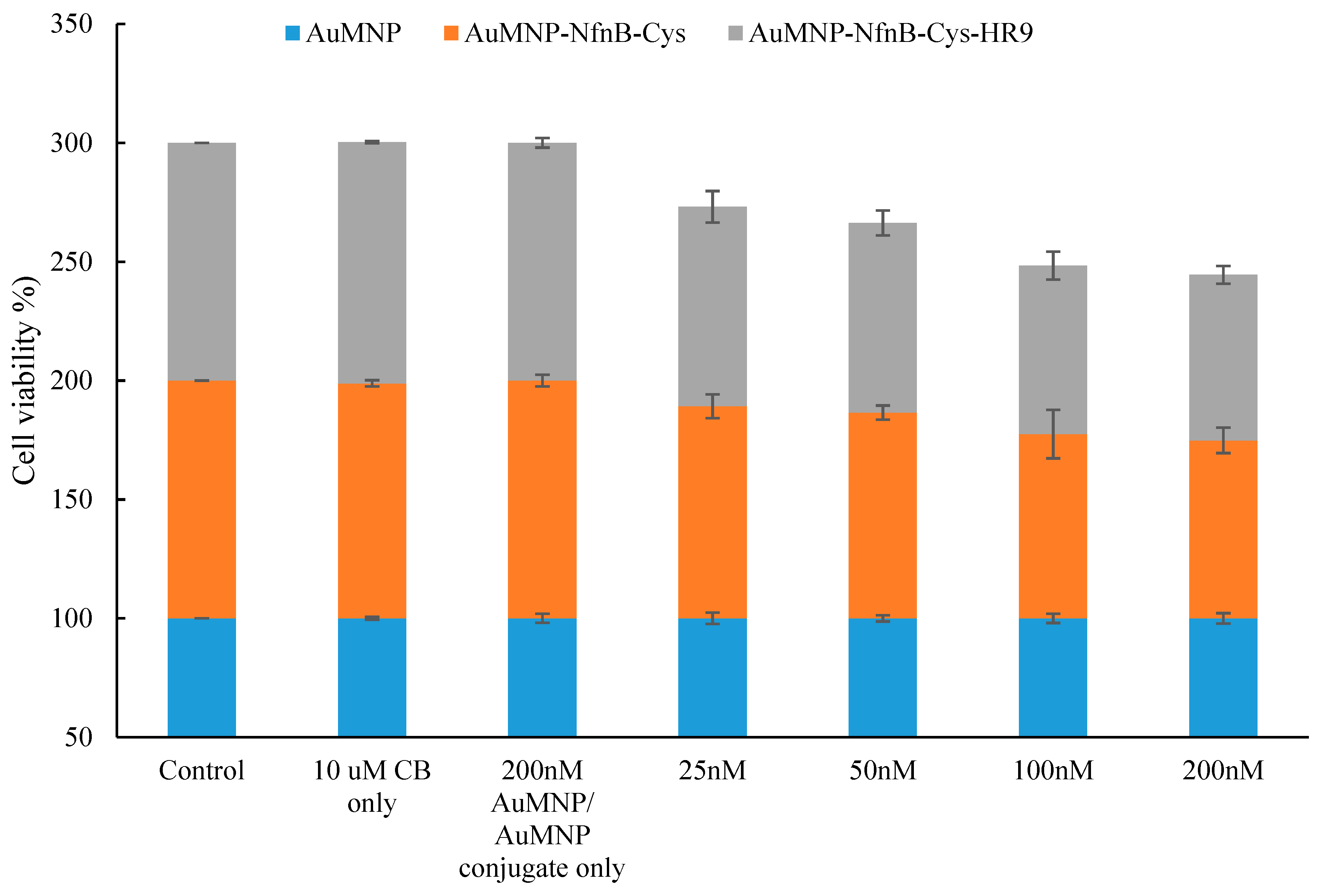{kind=link}
{kind=link}
| Conjugate | Vmax (µMs−1) | Kcat (s−1) | Km (µM) | Kcat/Km (µM−1s−1) |
|---|---|---|---|---|
| NfnB-Cys:HR9 | 7.98 ± 1.39 | 9.06 ± 0.82 | 3443 ± 916 | 0.00263 ± 4.6 × 104 |
| NfnB-Cys:Pep-1 | 7.43 ± 1.25 | 8.44 ± 0.73 | 2381 ± 695 | 0.00354 ± 6.5 × 104 |
| Conjugate | Ratio (2-NHOH:4-NHOH) |
|---|---|
| NfnB-Cys:HR9 | 36:64 |
| NfnB-Cys:Pep-1 | 35:65 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, S.D.; Hobbs, R.J.; Gwenin, V.V.; Ball, P.; Bennie, L.A.; Coulter, J.A.; Gwenin, C.D. Cell-Penetrating Peptides as a Tool for the Cellular Uptake of a Genetically Modified Nitroreductase for use in Directed Enzyme Prodrug Therapy. J. Funct. Biomater. 2019, 10, 45. https://doi.org/10.3390/jfb10040045
Anderson SD, Hobbs RJ, Gwenin VV, Ball P, Bennie LA, Coulter JA, Gwenin CD. Cell-Penetrating Peptides as a Tool for the Cellular Uptake of a Genetically Modified Nitroreductase for use in Directed Enzyme Prodrug Therapy. Journal of Functional Biomaterials. 2019; 10(4):45. https://doi.org/10.3390/jfb10040045
Chicago/Turabian StyleAnderson, Simon D., Robert J. Hobbs, Vanessa V. Gwenin, Patrick Ball, Lindsey A. Bennie, Jonathan A. Coulter, and Chris D. Gwenin. 2019. "Cell-Penetrating Peptides as a Tool for the Cellular Uptake of a Genetically Modified Nitroreductase for use in Directed Enzyme Prodrug Therapy" Journal of Functional Biomaterials 10, no. 4: 45. https://doi.org/10.3390/jfb10040045
APA StyleAnderson, S. D., Hobbs, R. J., Gwenin, V. V., Ball, P., Bennie, L. A., Coulter, J. A., & Gwenin, C. D. (2019). Cell-Penetrating Peptides as a Tool for the Cellular Uptake of a Genetically Modified Nitroreductase for use in Directed Enzyme Prodrug Therapy. Journal of Functional Biomaterials, 10(4), 45. https://doi.org/10.3390/jfb10040045