
Novel Biocement/Honey Composites for Bone Regenerative Medicine

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Cement Powder Mixture
2.2. Formation of Cement Pastes and Characterization of Honeys
2.2.1. Preparation of Cement Pastes
2.2.2. Analysis of Proteins in Honeys
2.2.3. Determination of Total Phenols, Flavonoids, and Sugars in Honeys and Cement Extracts during Release to SBF
2.2.4. Free Radical Scavenging Activity and Total Antioxidant Content
2.2.5. Characterization of Microstructures, Setting Time, and Mechanical Properties
2.3. Preparation of Cement Extracts and In Vitro Cytotoxicity Testing of Extracts
2.3.1. Analysis of Osteogenic Gene Expression in Differentiated Rat MSCs
2.3.2. Antibacterial Activity of Cements and Honey Samples
3. Results
3.1. XRD and FTIR Analysis of Cements
3.2. Microstructures of Cements and Morphology of HAP Particles in Cements, Setting Time, and Compressive Strength of Cements
3.3. Changes in pH and Release of Ca and Phosphate Ions during Soaking in Aqueous Solution
3.4. Analysis of Proteins, Release of Polyphenols and Sugars from Honey Cements, and Antioxidant Capacity of Cement Extracts
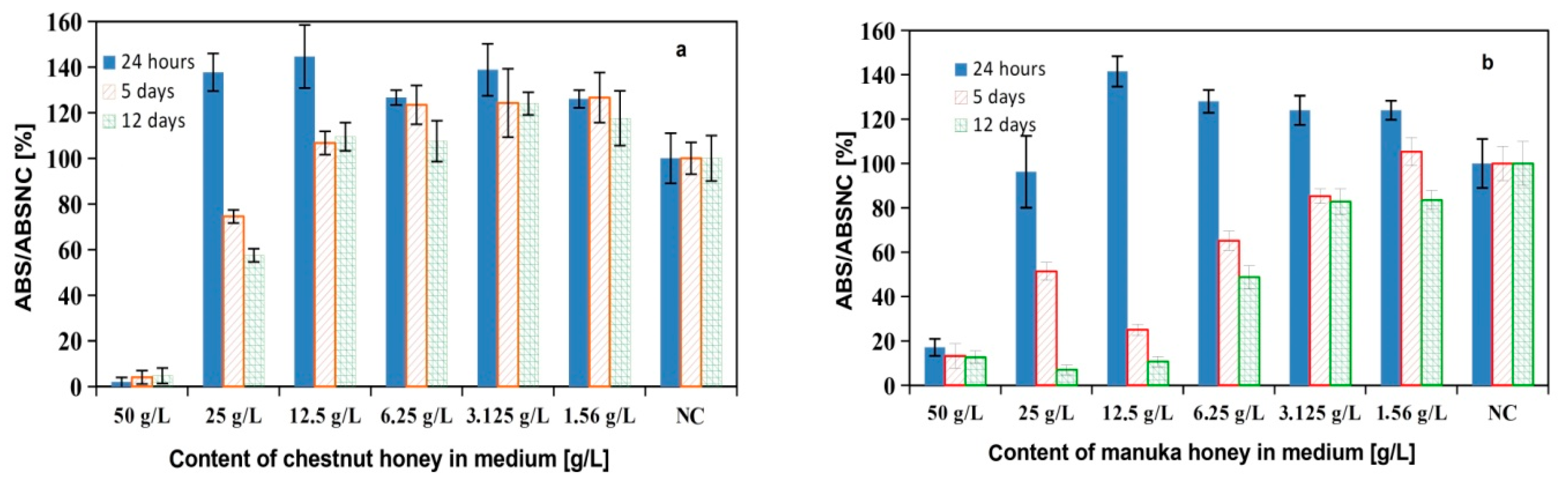
3.5. In Vitro Cytotoxicity Testing of Cements and Cement Extracts, Live/Dead Staining, and Gene Expression of Cell in Cement Extracts
3.6. Antibacterial Activity of Cements
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kageyama, T.; Akieda, H.; Sonoyama, Y.; Sato, K.; Yoshikawa, H.; Isono, H.; Hirota, M.; Kitajima, H.; Chun, J.S.; Maruo, S.; et al. Bone beads enveloped with vascular endothelial cells for bone regenerative medicine. Acta Biomater. 2023, 165, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wang, Y.; Sun, Y.; Gong, L.; Dai, X.; Meng, H.; Xu, W.; Ma, J.; Hu, Q.; Ma, X.; et al. Biodegradable silk fibroin scaffold doped with mineralized collagen induces bone regeneration in rat cranial defects. Int. J. Biol. Macromol. 2023, 235, 123861. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Martini, L.; Giavaresi, G.; Fini, M. Bone regenerative medicine: Metatarsus defects in sheep to evaluate new therapeutic strategies for human long bone defect. A systematic review. Injury 2020, 51, 1457–1467. [Google Scholar] [CrossRef]
- Carey, L.E.; Xu, H.H.K.; Simon, C.G.; Takagi, S.; Chow, L.C. Premixed rapid-setting calcium phosphate composites for bone repair. Biomaterials 2005, 26, 5002–5014. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Chow, L.C.; Markovic, M.; Friedman, C.D.; Costantino, P.D. Morphological and phase characterizations of retrieved calcium phosphate cement implants. J. Biomed. Mater. Res. (Appl. Biomater.) 2001, 58, 36–41. [Google Scholar] [CrossRef]
- Medvecky, L.; Giretova, M.; Stulajterova, R.; Luptakova, L.; Sopcak, T.; Girman, V. Osteogenic potential and properties of injectable silk fibroin/tetracalcium phosphate/monetite composite powder biocement systems. J. Biomed. Mater. Res. 2022, 110, 668–678. [Google Scholar] [CrossRef]
- Nie, Y.; Wang, T.; Wu, M.; Wang, C.; Wang, J.; Han, Z. Enhanced bioactivity and antimicrobial properties of α-tricalcium phosphate cement via PDA@Ag coating. Mater. Lett. 2023, 330, 133230. [Google Scholar] [CrossRef]
- Kouassi, M.; Michaïlesco, P.; Lacoste-Armynot, A.; Boudeville, P. Antibacterial effect of a hydraulic calcium phosphate cement for dental applications. J. Endod. 2003, 29, 100–103. [Google Scholar] [CrossRef]
- Valverde, S.; Ares, A.M.; Elmore, J.S.; Bernal, J. Recent trends in the analysis of honey constituents. Food Chem. 2022, 387, 132920. [Google Scholar] [CrossRef]
- Abd-El Aal, A.M.; El-Hadidy, M.R.; El-Mashad, N.B.; El-Sebaie, A.H. Antimicrobial effect of bee honey in comparison to antibiotics on organisms isolated from infected burns. Ann. Burn. Fire Disasters 2007, 20, 83–88. [Google Scholar]
- Deng, J.; Liu, R.; Lu, Q.; Hao, P.; Xu, A.; Zhang, J.; Tan, J. Biochemical properties, antibacterial and cellular antioxidant activities of buckwheat honey in comparison to manuka honey. Food Chem. 2018, 252, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.L.; Lim, L.Y.; Hammer, K.; Hettiarachchi, D.; Locher, C. A review of commonly used methodologies for assessing the antibacterial activity of honey and honey products. Antibiotics 2022, 11, 975. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E. Honey, wound repair and regenerative medicine. J. Funct. Biomater. 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Yupanqui Mieles, J.; Vyas, C.; Aslan, E.; Humphreys, G.; Diver, C.; Bartolo, P. Honey: An advanced antimicrobial and wound healing biomaterial for tissue engineering applications. Pharmaceutics 2022, 14, 1663. [Google Scholar] [CrossRef] [PubMed]
- Kamaruzzaman, M.A.; Chin, K.Y.; Ramli, E.S.M. A review of potential beneficial effects of honey on bone health. Evid. Based Complement. Alternat. Med. 2019, 2019, 8543618. [Google Scholar] [CrossRef]
- Martiniakova, M.; Kovacova, V.; Mondockov, V.; Zemanova, N.; Babikova, M.; Biro, R.; Ciernikova, S.; Omelka, R. Honey: A promising therapeutic supplement for the prevention and management of osteoporosis and breast cancer. Antioxidants 2023, 12, 567. [Google Scholar] [CrossRef]
- Zor, T.; Selinger, Z. Linearization of the bradford protein assay increases its sensitivity: Theoretical and experimental studies. Anal. Biochem. 1996, 236, 302–308. [Google Scholar] [CrossRef]
- Csepregi, K.; Kocsis, M.; Hideg, E. On the spectrophotometric determination of total phenolic and flavonoid contents. Acta Biol. Hung. 2013, 64, 500–509. [Google Scholar] [CrossRef]
- Otmani, A.; Amessis-Ouchemoukh, N.; Birinci, C.; Yahiaoui, S.; Kolayli, S.; Rodríguez-Flores, M.S.; Escuredo, O.; Seijo, M.C.; Ouchemoukh, S. Phenolic compounds and antioxidant and antibacterial activities of Algerian honeys. Food Biosci. 2021, 42, 101070. [Google Scholar] [CrossRef]
- Sadeer, N.B.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—Chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- ISO 10993-12; Biological Evaluation of Medical Devices—Part 12: Sample Preparation and Reference Materials. International Organization for Standardization: Geneva, Switzerland, 2012.
- ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Giretova, M.; Medvecky, L.; Petrovova, E.; Cizkova, D.; Danko, J.; Mudronova, D.; Slovinska, L.; Bures, R. Poly-hydroxybutyrate/chitosan 3D scaffolds promote in vitro and in vivo chondrogenesis. Appl. Biochem. Biotechnol. 2019, 189, 556–575. [Google Scholar] [CrossRef] [PubMed]
- Grässel, S.; Ahmed, N.; Göttl, C.; Grifka, J. Gene and protein expression profile of naive and osteo-chondrogenically differentiated rat bone marrow-derived mesenchymal progenitor cells. Int. J. Mol. Med. 2009, 23, 745–755. [Google Scholar] [CrossRef]
- Yang, J.; Chen, X.; Yuan, T.; Yang, X.; Fan, Y.; Zhang, X. Regulation of the secretion of immunoregulatory factors of mesenchymal stem cells (MSCs) by collagen-based scaffolds during chondrogenesis. Mater. Sci. Eng. C 2017, 70, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Yusop, N.; Battersby, P.; Alraies, A.; Sloan, A.J.; Moseley, R.; Waddington, R.J. Isolation and characterisation of mesenchymal stem cells from rat bone marrow and the endosteal niche: A comparative study. Stem Cells Int. 2018, 2018, 6869128. [Google Scholar] [CrossRef]
- Karaoz, E.; Aksoy, A.; Ayhan, S.; Sarıboyaci, A.E.; Kaymaz, F.; Kasap, M. Characterization of mesenchymal stem cells from rat bone marrow: Ultra-structural properties, differentiation potential and immunophenotypicmarkers. Histochem. Cell Biol. 2009, 132, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Su, W.; Ma, X.; Zhang, H.; Sun, Z.; Li, X. Comparison of the osteogenic capability of rat bone mesenchymal stem cells on collagen, collagen/hydroxyapatite, hydroxyapatite and biphasic calcium phosphate. Regen. Biomater. 2018, 5, 93–103. [Google Scholar] [CrossRef]
- Ren, F.; Ding, Y.; Leng, Y. Infrared spectroscopic characterization of carbonated apatite: A combined experimental and computational study. J. Biomed. Mater. Res. Part A 2014, 102A, 496–505. [Google Scholar] [CrossRef]
- Dżugan, M.; Miłek, M.; Sidor, E.; Buczkowicz, J.; Heclik, J.; Bocian, A. The application of SDS-PAGE protein and HPTLC amino acid profiling for verification of declared variety and geographical origin of honey. Food Anal. Methods 2023, 16, 1157–1171. [Google Scholar] [CrossRef]
- Lewkowski, O.; Mures, C.I.; Dobritzsch, D.; Fuszard, M.; Erler, S. The effect of diet on the composition and stability of proteins secreted by honey bees in honey. Insects 2019, 10, 282. [Google Scholar] [CrossRef]
- Dalas, E.; Koutsoukos, P.G. The effect of glucose on the crystallization of hydroxyapatite in aqueous solutions. J. Chem. Soc. Faraday Trans. 1989, 85, 2465–2472. [Google Scholar] [CrossRef]
- Walsh, D.; Kingstone, J.L.; Heywood, B.R.; Mann, S. Influence of monosaccharides and related molecules on the morphology of hydroxyapatite. J. Cryst. Growth 1993, 133, 1–12. [Google Scholar] [CrossRef]
- Murillo, L.M.; Iessi, I.L.; Quintino, M.P.; Damasceno, D.C.; Rodrigues, C.G. Glucose is an active chemical agent on degradation of hydroxyapatite nanostructure. Mater. Chem. Phys. 2020, 240, 122166. [Google Scholar]
- Palierse, E.; Masse, S.; Laurent, G.; Le Griel, P.; Mosser, G.; Coradin, T.; Jolivalt, C. Synthesis of hybrid polyphenol/hydroxyapatite nanomaterials with anti-radical properties. Nanomaterials 2022, 12, 3588. [Google Scholar] [CrossRef] [PubMed]
- Jerdioui, S.; Elansari, L.L.; Jaradat, N.; Jodeh, S.; Azzaoui, K.; Hammouti, B.; Lakrat, M.; Tahani, A.; Jama, C.; Bentiss, F. Effects of gallic acid on the nanocrystalline hydroxyapatite formation using the neutralization process. J. Trace Elem. Miner. 2022, 2, 100009. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Silvertooth, J.C. Inhibition of hydroxyapatite precipitation in the presence of fulvic, humic, and tannic acids. Soil Sci. Soc. Am. J. 1988, 52, 941–946. [Google Scholar] [CrossRef]
- Liu, X.; Lin, K.; Qian, R.; Chen, L.; Zhuo, S.; Chang, J. Growth of highly oriented hydroxyapatite arrays tuned by quercetin. Chem. Eur. J. 2012, 18, 5519–5523. [Google Scholar] [CrossRef]
- Jaouia, W.; Hachimia, M.B.; Koutita, T.; Lacout, J.L.; Ferhata, M. Effects of calcium phosphate apatites on the reaction of reducing sugars in an alkaline medium. Mater. Res. Bull. 2000, 35, 1419–1427. [Google Scholar] [CrossRef]
- Patria, R.D.; Islam, M.K.; Luo, L.; Leu, S.Y.; Varjani, S.; Xu, Y.; Wong, J.W.C.; Zhao, J. Hydroxyapatite-based catalysts derived from food waste digestate for efficient glucose isomerization to fructose. Green Synth. Catal. 2021, 2, 356–361. [Google Scholar] [CrossRef]
- Lachman, J.; Orsak, M.; Hejtmankova, A.; Kovarova, E. Evaluation of antioxidant activity and total phenolics of selected Czech honeys. LWT Food Sci. Technol. 2010, 43, 52–58. [Google Scholar] [CrossRef]
- Piljac-Žegarac, J.; Stipčević, T.; Belščak, A. Antioxidant properties and phenolic content of different floral origin honeys. J. ApiProduct ApiMedical Sci. 2009, 1, 43–50. [Google Scholar] [CrossRef]
- Ullah, I.; Zhang, W.; Yang, L.; Ullah, M.W.; Atta, O.M.; Khan, S.; Wu, B.; Wu, T.; Zhang, X. Impact of structural features of Sr/Fe co-doped HAp on the osteoblast proliferation and osteogenic differentiation for its application as a bone substitute. Mater. Sci. Eng. C 2020, 110, 110633. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, C.; Maoc, Y. Bismuth-doped injectable calcium phosphate cement with improved radiopacity and potent antimicrobial activity for root canal filling. Acta Biomater. 2010, 6, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Fadeeva, I.V.; Goldberg, M.A.; Preobrazhensky, I.I.; Mamin, G.V.; Davidova, G.A.; Agafonova, N.V.; Fosca, M.; Russo, F.; Barinov, S.M.; Cavalu, S.; et al. Improved cytocompatibility and antibacterial properties of zinc-substituted brushite bone cement based on β-tricalcium phosphate. J. Mater. Sci. Mater. Med. 2021, 32, 99. [Google Scholar] [CrossRef]
- Iglesias-Fernandez, M.; Buxadera-Palomero, J.; Sadowska, J.M.; Espanol, M.; Ginebra, M.P. Implementation of bactericidal topographies on biomimetic calcium phosphates and the potential effect of its reactivity. Biomater. Adv. 2022, 136, 212797. [Google Scholar] [CrossRef] [PubMed]
- Gbureck, U.; Knappe, O.; Hofmann, N.; Barralet, J.E. Antimicrobial properties of nanocrystalline tetracalcium phosphate cements. J. Biomed. Mater. Res. Part B Appl. Biomater. 2007, 83, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Podder, S.; Ghosh, C.K.; Bhattacharya, M.; Ghosh, J.; Mallik, A.K.; Dey, A.; Mukhopadhyay, A.K. ROS mediated high anti-bacterial efficacy of strain tolerant layered phase pure nano-calcium hydroxide. J. Mech. Behav. Biomed. Mater. 2017, 72, 110–128. [Google Scholar] [CrossRef]
- Gbureck, U.; Knappe, O.; Grover, L.M.; Barralet, J.E. Antimicrobial potency of alkali ion substituted calcium phosphate cements. Biomaterials 2005, 26, 6880–6886. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Gasparrini, M.; Forbes-Hernández, T.Y.; Mazzoni, L.; Giampieri, F. The composition and biological activity of honey: A focus on manuka honey. Foods 2014, 3, 420–432. [Google Scholar] [CrossRef]
- Bazaid, A.S.; Alamri, A.; Almashjary, M.N.; Qanash, H.; Almishaal, A.A.; Amin, J.; Binsaleh, N.K.; Kraiem, J.; Aldarhami, A.; Alafnan, A. Antioxidant, anticancer, antibacterial, antibiofilm properties and gas chromatography and mass spectrometry analysis of manuka honey: A nature’s bioactive honey. Appl. Sci. 2022, 12, 9928. [Google Scholar] [CrossRef]
- Schuhladen, K.; Mukoo, P.; Liverani, L.; Nescakova, Z.; Boccaccini, A.R. Manuka honey and bioactive glass impart methylcellulose foams with antibacterial effects for wound-healing applications. Biomed. Mater. 2020, 15, 065002. [Google Scholar] [CrossRef] [PubMed]
- Arango-Ospina, M.; Lasch, K.; Weidinger, J.; Boccaccini, A.R. Manuka honey and zein coatings impart bioactive glass bone tissue scaffolds antibacterial properties and superior mechanical properties. Front. Mater. 2021, 7, 610889. [Google Scholar] [CrossRef]
- Bondarenko, A.; Angrisani, N.; Meyer-Lindenberg, A.; Seitz, J.M.; Waizy, H.; Reifenrath, J. Magnesium-based bone implants: Immunohistochemical analysis of peri-implant osteogenesis by evaluation of osteopontin and osteocalcin expression. J. Biomed. Mater. Res. Part A 2014, 102, 1449–1457. [Google Scholar] [CrossRef]
- McKee, M.D.; Pedraza, C.E.; Kaartinen, M.T. Osteopontin and wound healing in bone. Cells Tissues Organs 2011, 194, 313–319. [Google Scholar] [CrossRef]
- Termine, J.D.; Kleinman, H.K.; Whitson, S.W.; Conn, K.M.; McGarvey, M.L.; Martin, G.R. Osteonectin, a bone-specific protein linking mineral to collagen. Cell 1981, 26, 99–105. [Google Scholar] [CrossRef]
- Komori, T. Functions of osteocalcin in bone, pancreas, testis, and muscle. Int. J. Mol. Sci. 2020, 21, 7513. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.F.; Shao, J.; Shi, C.J.; Li, Z.P.; Fu, W.M.; Zhang, J.F. Apigenin promotes osteogenic differentiation of mesenchymal stem cells and accelerates bone fracture healing via activating Wnt/b-catenin signaling. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E760–E771. [Google Scholar] [CrossRef]
- Huo, J.F.; Zhang, M.L.; Wang, X.X.; Zou, D.H. Chrysin induces osteogenic differentiation of human dental pulp stem cells. Exp. Cell Res. 2021, 400, 112466. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | References |
|---|---|---|
| B-actin rat | F: GTAGCCATCCAGGCTGTGTT R: CCCTCATAGATGGGCAGAGT | [25] |
| Type I collagen rat | F: CCAGCTGACCTTCCTGCGCC R: CGGTGTGACTCGTGCAGCCA | [26] |
| Osteocalcin rat | F: ACAGACAAGTCCCACACAGCAACT R: CCTGCTTGGACATGAAGGCTTTGT | [27] |
| Osteopontin rat | F: CCGATGAATCTGATGAGTCCTT R: TCCAGCTGACTTGACTCATG | [28] |
| Osteonectin rat | F: GGAAGCTGCAGAAGAGATGG R: TGCACACCTTTTCAAACTCG | [28] |
| Alkaline Phosphatase rat | F: AACCTGACTGACCCTTCCCTCT R: TCAATCCTGCCTCCTTCCACTA | [29] |
| Polyphenols | Chestnut [µg/g of Honey] | Manuka [µg/g of Honey] |
|---|---|---|
| Luteolin | 8.2 ± 1 | 3.9 ± 1 |
| Chrysin | 55.6 ± 6 | 26.1 ± 3 |
| Gallic acid | 44.8 ± 7 | 46.4 ± 6 |
| Kaempferol | 18.1 ± 2 | 6.9 ± 1 |
| 3,4,5-trimethoxybenzoic acid | 26.3 ± 3 | 10.4 ± 1 |
| Chlorogenic acid | 162.1 ± 11 | ND |
| Rutin | 27.5 ± 5 | ND |
| Quercetin | 7.5 ± 1 | 4.3 ± 1 |
| Ferulic acid | 26.8 ± 3 | 12.9 ± 2 |
| Coumaric acid | 76.0 ± 6 | 32.4 ± 2 |
| Syringic acid | 37.9 ± 5 | 16.6 ± 3 |
| Methylsyringate | 37.0 ± 4 | 134.3 ± 12 |
| Caffeic acid | 68.2 ± 8 | 31.8 ± 4 |
| Protocatechuic acid | 36.7 ± 3 | 100.3 ± 11 |
| Sugars | [wt%] | [wt%] |
| Fructose | 50.1 ± 2 | 40.5 ± 2 |
| Glucose | 27.5 ± 1 | 34.0 ± 2 |
| Saccharose | 3.8 ± 0.5 | 2.3 ± 0.3 |
| Maltose | 5.7 ± 0.8 | 4.8 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medvecky, L.; Giretova, M.; Stulajterova, R.; Sopcak, T.; Jevinova, P.; Luptakova, L. Novel Biocement/Honey Composites for Bone Regenerative Medicine. J. Funct. Biomater. 2023, 14, 457. https://doi.org/10.3390/jfb14090457
Medvecky L, Giretova M, Stulajterova R, Sopcak T, Jevinova P, Luptakova L. Novel Biocement/Honey Composites for Bone Regenerative Medicine. Journal of Functional Biomaterials. 2023; 14(9):457. https://doi.org/10.3390/jfb14090457
Chicago/Turabian StyleMedvecky, Lubomir, Maria Giretova, Radoslava Stulajterova, Tibor Sopcak, Pavlina Jevinova, and Lenka Luptakova. 2023. "Novel Biocement/Honey Composites for Bone Regenerative Medicine" Journal of Functional Biomaterials 14, no. 9: 457. https://doi.org/10.3390/jfb14090457
APA StyleMedvecky, L., Giretova, M., Stulajterova, R., Sopcak, T., Jevinova, P., & Luptakova, L. (2023). Novel Biocement/Honey Composites for Bone Regenerative Medicine. Journal of Functional Biomaterials, 14(9), 457. https://doi.org/10.3390/jfb14090457