
Effects of Zinc Oxide Nanoparticles on Physiological and Anatomical Indices in Spring Barley Tissues

 ,
,  ,
, 
 ,
,  , and
, and 
Abstract
:
1. Introduction
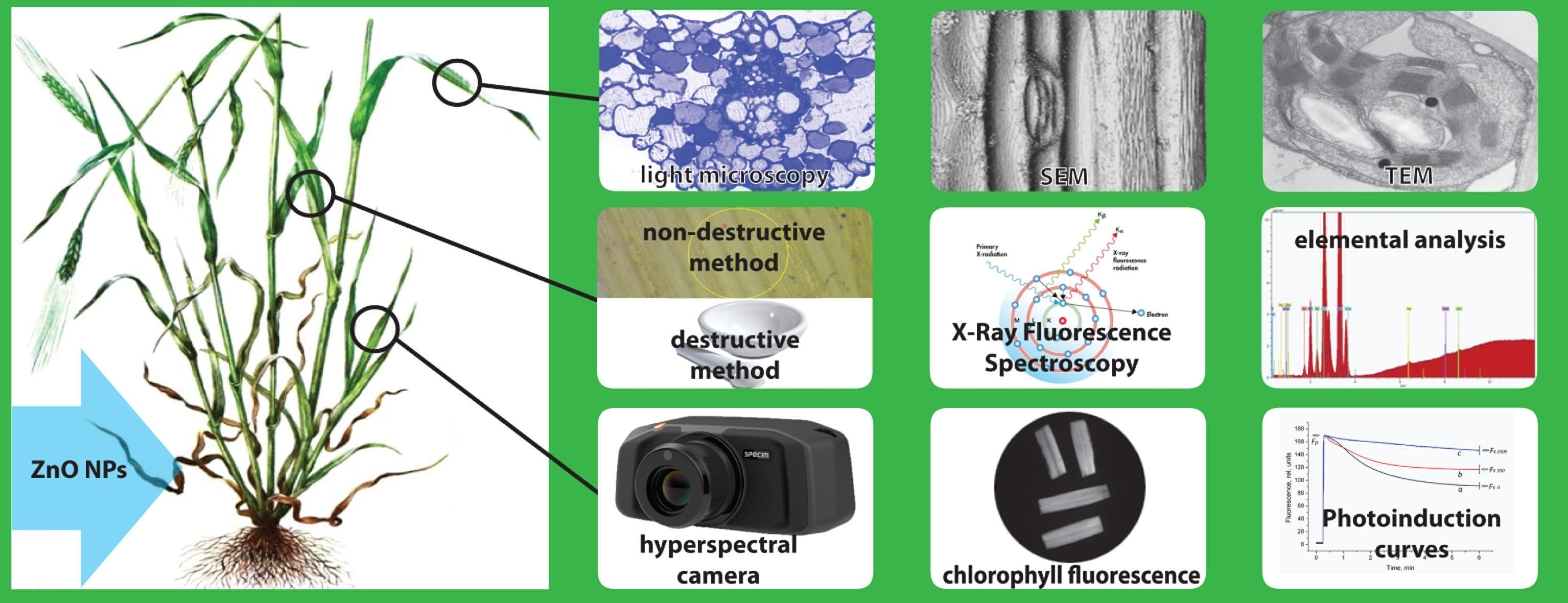
2. Materials and Methods
2.1. Experimental Set-Up
2.1.1. Nanoparticle Preparation and Characterization
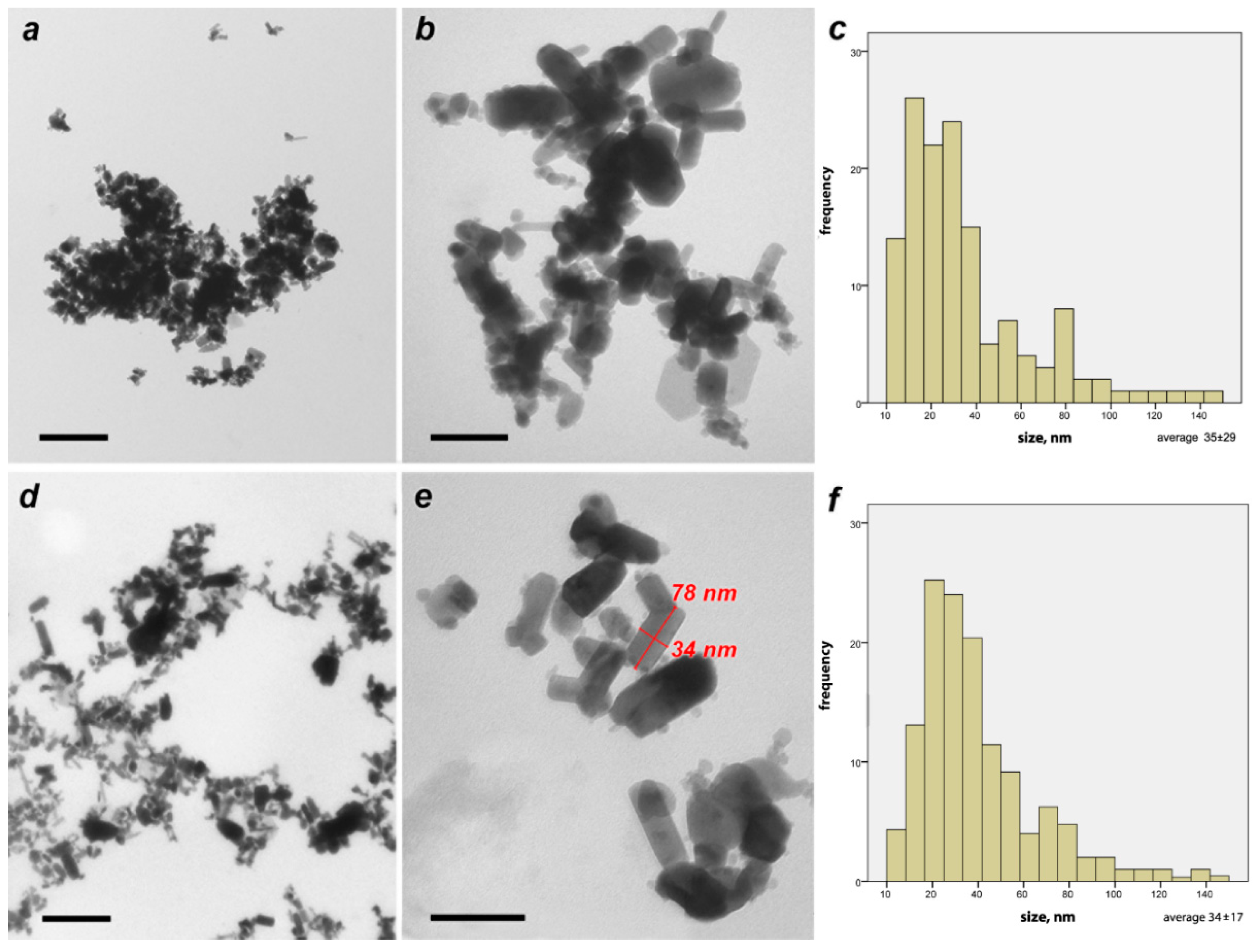
Characterization of ZnO Nanoparticles by Transmission Electron Microscopy (TEM)
Powder X-ray Diffraction of ZnO Nanoparticles
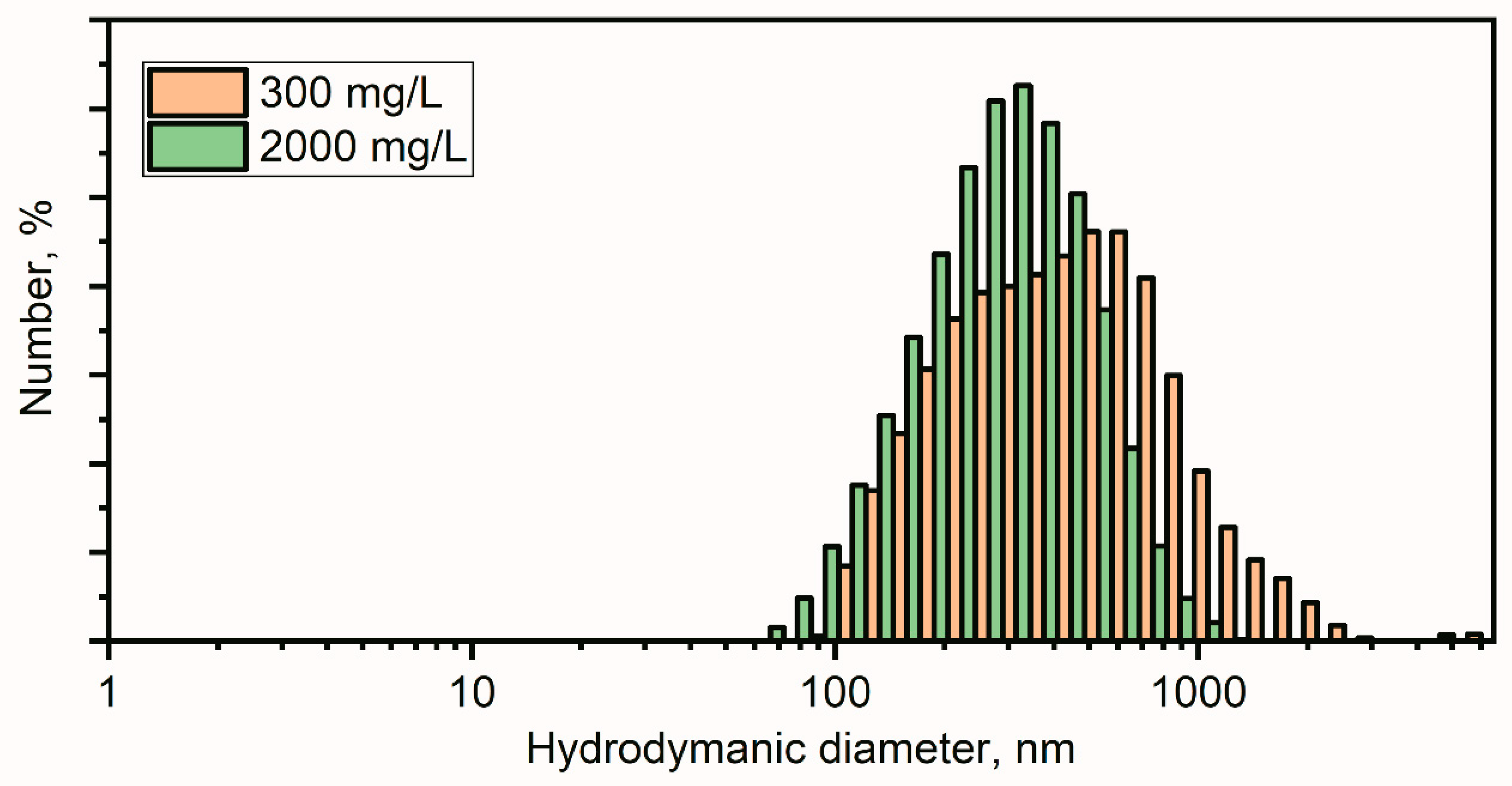
Hydrodynamic Dynamic Light Scattering (DLS) and ζ-Potential Analysis of ZnO Nanoparticles
2.1.2. Plant Growth and Performance
2.2. Elemental Analysis in Plant Tissues
2.2.1. Non-Destructive Method
2.2.2. Destructive Method
2.3. Physiological Indices
2.3.1. Photosynthetic Effectiveness Evaluating by Fluorescence Kinetic Parameters
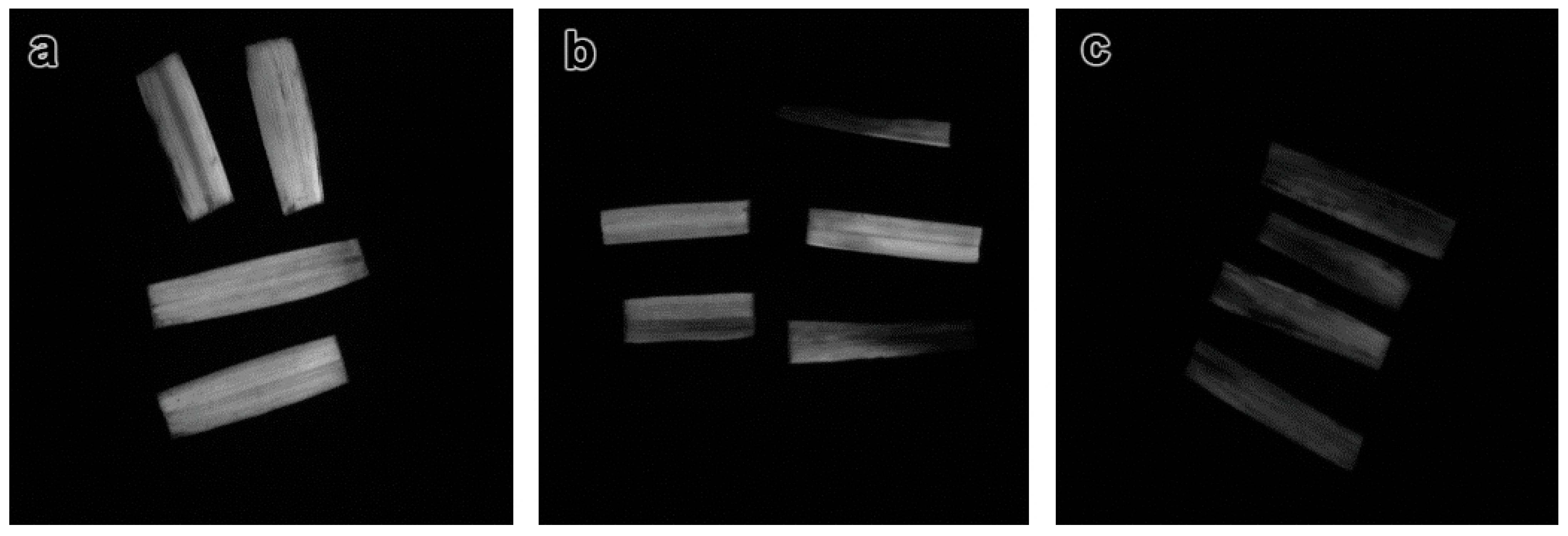
2.3.2. Measurement of Stomatal Aperture and Trichome Morphology
2.4. Anatomical Indices
2.4.1. Cellular and Sub-Cellular Structural and Ultrastructural Observations
2.4.2. Cytomorphometric Quantification
2.5. Statistical Analysis
3. Results
3.1. Characterization of ZnO Nanoparticles, Crystalline Sizes, Hydrodynamic Sizes of the Particles and Their Stability in Colloidal Solutation
3.2. Zn Accumulation in Above-Ground Tissues of H. sativum
3.3. Effects of ZnO NPs on Physiological Indices
3.3.1. Chlorophyll Fluorescence Kinetics in H. sativum Leaves from Plants Treated with Different Concentrations of ZnO NPs
3.3.2. Effects of ZnO NPs on Stomata and Trichome Morphology
3.3.3. Effect of ZnO NPs on Leaf Cellular and Sub-Cellular Organelles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraceto, L.F.; Grillo, R.; de Medeiros, G.A.; Scognamiglio, V.; Rea, G.; Bartolucci, C. Nanotechnology in Agriculture: Which Innovation Potential Does It Have? Front. Environ. Sci. 2016, 4, 20. [Google Scholar] [CrossRef]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.u.; Ashraf, I.; Sanaullah, M. Nanotechnology in Agriculture: Current Status, Challenges and Future Opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef]
- Servin, A.D.; White, J.C. Nanotechnology in Agriculture: Next Steps for Understanding Engineered Nanoparticle Exposure and Risk. NanoImpact 2016, 1, 9–12. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of Zinc-oxide Nanoparticles on Soil, Plants, Animals and Soil Organisms: A Review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, Zinc Nanoparticles and Plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.; Minkina, T.; Mazarji, M.; Shende, S.; Sushkova, S.; Mandzhieva, S.; Burachevskaya, M.; Chaplygin, V.; Singh, A.; Jatav, H. Accumulation of Nanoparticles in the Soil-plant Systems and Their Effects on Human Health. Ann. Agric. Sci. 2020, 65, 137–143. [Google Scholar] [CrossRef]
- Carrouel, F.; Viennot, S.; Ottolenghi, L.; Gaillard, C.; Bourgeois, D. Nanoparticles as Anti-Microbial, Anti-Inflammatory, and Remineralizing Agents in Oral Care Cosmetics: A Review of the Current Situation. Nanomaterials 2020, 10, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [Green Version]
- Shende, S.; Rajput, V.; Gade, A.; Minkina, T.; Sushkova, S.N.; Mandzhieva, S.S.; Boldyreva, V.E. Metal-based Green Synthesized Nanoparticles: Boon for Sustainable Agriculture and Food Security. IEEE Trans. NanoBiosci. 2021, 1. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Sushkova, S.; Behal, A.; Maksimov, A.; Blicharska, E.; Ghazaryan, K.; Movsesyan, H.; Barsova, N. ZnO and CuO Nanoparticles: A Threat to Soil Organisms, Plants, and Human Health. Environ. Geochem. Health 2019, 42, 147–158. [Google Scholar] [CrossRef]
- Jin, M.; Li, N.; Sheng, W.; Ji, X.; Liang, X.; Kong, B.; Yin, P.; Li, Y.; Zhang, X.; Liu, K. Toxicity of Different Zinc Oxide Nanomaterials and Dose-dependent Onset and Development of Parkinson’s Disease-like Symptoms Induced by Zinc Oxide Nanorods. Environ. Int. 2021, 146, 106179. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Mir, A.R.; Hayat, S. Role of Zinc Oxide Nanoparticles in Countering Negative Effects Generated by Cadmium in Lycopersicon esculentum. J. Plant Growth Regul. 2020, 40, 101–115. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, H.; Tu, C.; Hu, X.; Li, L.; Luo, Y.; Christie, P. Phytotoxicity of ZnO Nanoparticles and the Released Zn(II) Ion to Corn (Zea mays L.) and Cucumber (Cucumis sativus L.) During Germination. Environ. Sci. Pollut. Res. Int. 2015, 22, 11109–11117. [Google Scholar] [CrossRef]
- Javed, R.; Usman, M.; Yucesan, B.; Zia, M.; Gurel, E. Effect of Zinc Oxide (ZnO) Nanoparticles on Physiology and Steviol Glycosides Production in Micropropagated Shoots of Stevia rebaudiana Bertoni. Plant Physiol. Biochem. 2017, 110, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Kumari, A.; Harish; Singh, V.K.; Verma, K.K.; Mandzhieva, S.; Sushkova, S.; Srivastava, S.; Keswani, C. Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles. Plants 2021, 10, 1221. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO Nanoparticle Biosynthesis and its Effect on Phosphorous-Mobilizing Enzyme Secretion and Gum Contents in Clusterbean (Cyamopsis tetragonoloba L.). Agric. Res. 2013, 2, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Zinc Oxide Nanoparticles (ZnONPs) as a Novel Nanofertilizer: Influence on Seed Yield and Antioxidant Defense System in Soil Grown Soybean (Glycine max cv. Kowsar). Sci. Total Environ. 2020, 738, 140240. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc Oxide Nanoparticle-mediated Changes in Photosynthetic Efficiency and Antioxidant System of Tomato Plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Elmer, W.H.; White, J.C. The Use of Metallic Oxide Nanoparticles to Enhance Growth of Tomatoes and Eggplants in Disease Infested Soil or Soilless Medium. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Ramesh, M.; Palanisamy, K.; Babu, K.; Sharma, N.K. Effects of Bulk & Nano-Titanium Dioxide and Zinc Oxide on Physio-Morphological Changes in Triticum aestivum Linn. J. Glob. Biosci. 2014, 3, 415–422. [Google Scholar]
- Wang, X.; Yang, X.; Chen, S.; Li, Q.; Wang, W.; Hou, C.; Gao, X.; Wang, L.; Wang, S. Zinc Oxide Nanoparticles Affect Biomass Accumulation and Photosynthesis in Arabidopsis. Front. Plant Sci. 2016, 6, 1243. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Kumar, A. Investigating Long-term Effect of Nanoparticles on Growth of Raphanus sativus Plants: A Trans-generational Study. Ecotoxicology 2018, 27, 23–31. [Google Scholar] [CrossRef]
- Singh, D.; Kumar, A. Impact of Irrigation Using Water Containing CuO and ZnO Nanoparticles on Spinach oleracea Grown in Soil Media. Bull. Environ. Contam. Toxicol. 2016, 97, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.M.; Chung, I.M. Regulation of Morphological, Molecular and Nutrient Status in Arabidopsis thaliana Seedlings in Response to ZnO Nanoparticles and Zn Ion Exposure. Sci. Total Environ. 2017, 575, 187–198. [Google Scholar] [CrossRef]
- Wan, J.; Wang, R.; Wang, R.; Ju, Q.; Wang, Y.; Xu, J. Comparative Physiological and Transcriptomic Analyses Reveal the Toxic Effects of ZnO Nanoparticles on Plant Growth. Environ. Sci. Technol. 2019, 53, 4235–4244. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Shweta; Singh, S.; Singh, S.; Pandey, R.; Singh, V.P.; Sharma, N.C.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. An Overview on Manufactured Nanoparticles in Plants: Uptake, Translocation, Accumulation and Phytotoxicity. Plant Physiol. Biochem. 2017, 110, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of Nanoparticles with Edible Plants and Their Possible Implications in the Food Chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, V.D.; Minkina, T.; Sushkova, S.; Mandzhieva, S.; Fedorenko, A.; Lysenko, V.; Bederska-Błaszczyk, M.; Olchowik, J.; Tsitsuashvili, V.; Chaplygin, V. Structural and Ultrastructural Changes in Nanoparticle Exposed Plants. In Nanoscience for Sustainable Agriculture; Pudake, R.N., Chauhan, N., Kole, C., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 281–295. [Google Scholar]
- Elsheery, N.I.; Sunoj, V.S.J.; Wen, Y.; Zhu, J.J.; Muralidharan, G.; Cao, K.F. Foliar Application of Nanoparticles Mitigates the Chilling Effect on Photosynthesis and Photoprotection in Sugarcane. Plant Physiol. Biochem. 2020, 149, 50–60. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant Nanobionics Approach to Augment Photosynthesis and Biochemical Sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barhoumi, L.; Oukarroum, A.; Taher, L.B.; Smiri, L.S.; Abdelmelek, H.; Dewez, D. Effects of Superparamagnetic Iron Oxide Nanoparticles on Photosynthesis and Growth of the Aquatic Plant Lemna gibba. Arch. Environ. Contam. Toxicol. 2015, 68, 510–520. [Google Scholar] [CrossRef]
- Wang, X.P.; Li, Q.Q.; Pei, Z.M.; Wang, S.C. Effects of Zinc Oxide Nanoparticles on the Growth, Photosynthetic Traits, and Antioxidative Enzymes in Tomato Plants. Biol. Plant. 2018, 62, 801–808. [Google Scholar] [CrossRef]
- Bradfield, S.J.; Kumar, P.; White, J.C.; Ebbs, S.D. Zinc, Copper, or Cerium Accumulation from Metal Oxide Nanoparticles or Ions in Sweet Potato: Yield Effects and Projected Dietary Intake from Consumption. Plant Physiol. Biochem. 2017, 110, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Radi, A.A.; Farghaly, F.A.; Al-Kahtany, F.A.; Hamada, A.M. Zinc Oxide Nanoparticles-mediated Changes in Ultrastructure and Macromolecules of Pomegranate Callus Cells. Plant Cell Tissue Organ Cult. 2018, 135, 247–261. [Google Scholar] [CrossRef]
- Ghosh, M.; Jana, A.; Sinha, S.; Jothiramajayam, M.; Nag, A.; Chakraborty, A.; Mukherjee, A.; Mukherjee, A. Effects of ZnO Nanoparticles in Plants: Cytotoxicity, Genotoxicity, Deregulation of Antioxidant Defenses, and Cell-cycle Arrest. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 807, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Phull, A.-R.; Zia, M. Elemental Zinc to Zinc Nanoparticles: Is ZnO NPs Crucial for Life? Synthesis, Toxicological, and Environmental Concerns. Nanotechnol. Rev. 2018, 7, 413–441. [Google Scholar] [CrossRef]
- Peng, C.; Tong, H.; Shen, C.; Sun, L.; Yuan, P.; He, M.; Shi, J. Bioavailability and Translocation of Metal Oxide Nanoparticles in the Soil-rice Plant System. Sci. Total Environ. 2020, 713, 136662. [Google Scholar] [CrossRef]
- Zoufan, P.; Baroonian, M.; Zargar, B. ZnO Nanoparticles-induced Oxidative Stress in Chenopodium murale L, Zn Uptake, and Accumulation Under Hydroponic Culture. Environ. Sci. Pollut. Res. Int. 2020, 27, 11066–11078. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, V.S.; Varduny, T.V.; Kosenko, P.O.; Kosenko, Y.V.; Chugueva, O.I.; Semin, L.V.; Gorlachev, I.A.; Tarasov, E.K.; Guskova, O.S. Video Registration as a Method for Studying Kinetic Parameters of Chlorophyll Fluorescence in Ficus benjamina Leaves. Russ. J. Plant Physiol. 2014, 61, 419–425. [Google Scholar] [CrossRef]
- Fedorenko, G.M.; Fedorenko, A.G.; Minkina, T.M.; Mandzhieva, S.S.; Rajput, V.D.; Usatov, A.V.; Sushkova, S.N. Method for Hydrophytic Plant Sample Preparation for Light and Electron Microscopy (studies on Phragmites australis Cav.). MethodsX 2018, 5, 1213–1220. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in Plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- AlShrouf, A. Hydroponics, Aeroponic and Aquaponic as Compared with Conventional Farming. Am. Sci. Res. J. Eng. Technol. Sci. 2017, 27, 247–255. [Google Scholar]
- Harwood, W.A. An Introduction to Barley: The Crop and the Model. In Barley: Methods and Protocols; Harwood, W.A., Ed.; Springer New York: New York, NY, USA, 2019; pp. 1–5. [Google Scholar]
- Tymoszuk, A.; Wojnarowicz, J. Zinc Oxide and Zinc Oxide Nanoparticles Impact on In Vitro Germination and Seedling Growth in Allium cepa L. Materials 2020, 13, 2784. [Google Scholar] [CrossRef] [PubMed]
- Nemček, L.; Šebesta, M.; Urík, M.; Bujdoš, M.; Dobročka, E.; Vávra, I. Impact of Bulk ZnO, ZnO Nanoparticles and Dissolved Zn on Early Growth Stages of Barley—A Pot Experiment. Plants 2020, 9, 1365. [Google Scholar] [CrossRef]
- Da Cruz, T.N.M.; Savassa, S.M.; Montanha, G.S.; Ishida, J.K.; de Almeida, E.; Tsai, S.M.; Lavres Junior, J.; Pereira de Carvalho, H.W. A New Glance on Root-to-shoot in Vivo Zinc Transport and Time-dependent Physiological Effects of ZnSO(4) and ZnO Nanoparticles on Plants. Sci. Rep. 2019, 9, 10416. [Google Scholar] [CrossRef] [Green Version]
- López-Moreno, M.L.; de la Rosa, G.; Hernández-Viezcas, J.Á.; Castillo-Michel, H.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Evidence of the Differential Biotransformation and Genotoxicity of ZnO and CeO2 Nanoparticles on Soybean (Glycine max) Plants. Environ. Sci. Technol. 2010, 44, 7315–7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evens, N.P. The Regulatory Mechanisms Controlling Zinc Content in Wheat. Ph.D. Thesis, University of Southampton, Southampton, UK, January 2017; p. 257. [Google Scholar]
- Mousavi Kouhi, S.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Long-term Exposure of Rapeseed (Brassica napus L.) to ZnO Nanoparticles: Anatomical and Ultrastructural Responses. Environ. Sci. Pollut. Res. Int. 2015, 22, 10733–10743. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Shekhawat, G.S. Toxicity of ZnO Engineered Nanoparticles and Evaluation of Their Effect on Growth, Metabolism and Tissue Specific Accumulation in Brassica juncea. J. Environ. Chem. Eng. 2014, 2, 105–114. [Google Scholar] [CrossRef]
- Asli, S.; Neumann, P.M. Colloidal Suspensions of Clay or Titanium Dioxide Nanoparticles Can Inhibit Leaf Growth and Transpiration via Physical Effects on Root Water Transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F.; Langsdorf, G. Chlorophyll Fluorescence Imaging of Photosynthetic Activity in Sun and Shade Leaves of Trees. Photosynth. Res. 2007, 93, 235–244. [Google Scholar] [CrossRef]
- Rajput, V.D.; Chen, Y.; Ayup, M. Effects of High Salinity on Physiological and Anatomical Indices in the Early Stages of Populus euphratica Growth. Russ. J. Plant Physiol. 2015, 62, 229–236. [Google Scholar] [CrossRef]
- Caldelas, C.; Poitrasson, F.; Viers, J.; Araus, J.L. Stable Zn Isotopes Reveal the Uptake and Toxicity of Zinc Oxide Engineered Nanomaterials in Phragmites australis. Environ. Sci. Nano 2020, 7, 1927–1941. [Google Scholar] [CrossRef]
- Song, U.; Lee, S. Phytotoxicity and Accumulation of Zinc Oxide Nanoparticles on the Aquatic Plants Hydrilla verticillata and Phragmites australis: Leaf-type-dependent Responses. Environ. Sci. Pollut. Res. Int. 2016, 23, 8539–8545. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; O’Halloran, J.; Jansen, M.A. The Toxicity of Zinc Oxide Nanoparticles to Lemna minor (L.) is Predominantly Caused by Dissolved Zn. Aquat. Toxicol. 2016, 174, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.V.J.; Sharma, P.K. Effect of Copper Oxide Nanoparticles on Growth, Morphology, Photosynthesis, and Antioxidant Response in Oryza sativa. Photosynthetica 2015, 54, 110–119. [Google Scholar] [CrossRef]
- Perreault, F.; Samadani, M.; Dewez, D. Effect of Soluble Copper Released from Copper Oxide Nanoparticles Solubilisation on Growth and Photosynthetic Processes of Lemna gibba L. Nanotoxicology 2014, 8, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xing, B. Root Uptake and Phytotoxicity of ZnO Nanoparticles. Environ. Sci. Technol. 2008, 42, 5580–5585. [Google Scholar] [CrossRef]
- Austin, J.R., 2nd; Frost, E.; Vidi, P.A.; Kessler, F.; Staehelin, L.A. Plastoglobules are Lipoprotein Subcompartments of the Chloroplast that are Permanently Coupled to Thylakoid Membranes and Contain Biosynthetic Enzymes. Plant Cell 2006, 18, 1693–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalau, C.M.; Mohedano Rde, A.; Schmidt, E.C.; Bouzon, Z.L.; Ouriques, L.C.; dos Santos, R.W.; da Costa, C.H.; Vicentini, D.S.; Matias, W.G. Toxicological Effects of Copper Oxide Nanoparticles on the Growth Rate, Photosynthetic Pigment Content, and Cell Morphology of the Duckweed Landoltia punctata. Protoplasma 2015, 252, 221–229. [Google Scholar] [CrossRef]
- Skórzyńska-Polit, E.; Baszyński, T. Differences in Sensitivity of the Photosynthetic Apparatus in Cd-stressed Runner Bean Plants in Relation to Their Age. Plant Sci. 1997, 128, 11–21. [Google Scholar] [CrossRef]
- Pietrini, F.; Iannelli, M.A.; Pasqualini, S.; Massacci, A. Interaction of Cadmium with Glutathione and Photosynthesis in Developing Leaves and Chloroplasts of Phragmites australis (Cav.) Trin. ex Steudel. Plant Physiol. 2003, 133, 829. [Google Scholar] [CrossRef] [Green Version]
- Siedlecka, A.; Krupa, Z. Cd/Fe Interaction in Higher Plants—Its Consequences for the Photosynthetic Apparatus. Photosynthetica 1999, 36, 321–331. [Google Scholar] [CrossRef]
- Zhuang, X.; Jiang, L. Chloroplast Degradation: Multiple Routes Into the Vacuole. Front. Plant Sci. 2019, 10, 359. [Google Scholar] [CrossRef] [Green Version]
- Molas, J. Changes of Chloroplast Ultrastructure and Total Chlorophyll Concentration in Cabbage Leaves Caused by Excess of Organic Ni(II) Complexes. Environ. Exp. Bot. 2002, 47, 115–126. [Google Scholar] [CrossRef]
- Maksymiec, W.; BaszzDski, T.; Bednara, J. Responses of Runner Bean Plants to Excess Copper as a Function of Plant Growth Stages: Effects on Morphology and Structure of Primary Leaves and Their Chloroplast Ultrastructure. Photosynthetica 1995, 31, 427–435. [Google Scholar]
- Hakmaoui, A.; Ater, M.; Bóka, K.; Barón, M. Copper and Cadmium Tolerance, Uptake and Effect on Chloroplast Ultrastructure. Studies on Salix purpurea and Phragmites australis. Z. Nat. 2007, 62, 417–426. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Above-Ground Tissues |
|---|---|
| Control | 21.9 ± 2.1 |
| 300 mg/L ZnO NPs | 84.2 ± 6.9 |
| 2000 mg/L ZnO NPs | 224.6 ± 17.3 |
| Treatments | Average Chlorenchyma Cell Area, µm2 | Shape of Chlorenchyma | Ratio of Chlorenchyma Cell Area to Total Leaf Cut Area | Stomatal | Trichomes | ||
|---|---|---|---|---|---|---|---|
| Circular | Round | Solid | |||||
| Control | 131 ± 12 | 0.88 | 0.74 | 0.99 | 0.57 | 32.0 ± 1.2 | 12.4 ± 1.5 |
| 300 mg/L ZnO NPs | 146 ± 14 | 0.77 | 0.65 | 0.95 | 0.62 | 39 * ± 1.7 | 18 * ± 2.6 |
| 2000 mg/L ZnO NPs | 138 ± 15 | 0.69 | 0.58 | 0.89 | 0.64 | 30 ± 1.8 | 10 ± 1.1 |
| Treatments | Size Chloroplasts, µm2 | Number of Chloroplasts per Cell | Plastoglobules per Cell | Starch Granules per Cell |
|---|---|---|---|---|
| Control | 7.42 ± 0.72 | 6 ± 0.6 | 16 ± 3 | n/d |
| 300 mg/L ZnO NPs | 5.25 ± 0.70 | 5 ± 0.7 | 12 ± 2 | 3.0 ± 0.2 |
| 2000 mg/L ZnO NPs | 4.17 ± 0.57 | 4 ± 0.5 | 18 ± 4 | 7.1 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajput, V.D.; Minkina, T.; Fedorenko, A.; Chernikova, N.; Hassan, T.; Mandzhieva, S.; Sushkova, S.; Lysenko, V.; Soldatov, M.A.; Burachevskaya, M. Effects of Zinc Oxide Nanoparticles on Physiological and Anatomical Indices in Spring Barley Tissues. Nanomaterials 2021, 11, 1722. https://doi.org/10.3390/nano11071722
Rajput VD, Minkina T, Fedorenko A, Chernikova N, Hassan T, Mandzhieva S, Sushkova S, Lysenko V, Soldatov MA, Burachevskaya M. Effects of Zinc Oxide Nanoparticles on Physiological and Anatomical Indices in Spring Barley Tissues. Nanomaterials. 2021; 11(7):1722. https://doi.org/10.3390/nano11071722
Chicago/Turabian StyleRajput, Vishnu D., Tatiana Minkina, Aleksei Fedorenko, Natalia Chernikova, Tara Hassan, Saglara Mandzhieva, Svetlana Sushkova, Vladimir Lysenko, Mikhail A. Soldatov, and Marina Burachevskaya. 2021. "Effects of Zinc Oxide Nanoparticles on Physiological and Anatomical Indices in Spring Barley Tissues" Nanomaterials 11, no. 7: 1722. https://doi.org/10.3390/nano11071722
APA StyleRajput, V. D., Minkina, T., Fedorenko, A., Chernikova, N., Hassan, T., Mandzhieva, S., Sushkova, S., Lysenko, V., Soldatov, M. A., & Burachevskaya, M. (2021). Effects of Zinc Oxide Nanoparticles on Physiological and Anatomical Indices in Spring Barley Tissues. Nanomaterials, 11(7), 1722. https://doi.org/10.3390/nano11071722