


Trophic Transfer of Single-Walled Carbon Nanotubes at the Base of the Food Chain and Toxicological Response

Abstract
:1. Introduction
2. Experimental Section
2.1. Preparation and Characterization of SWCNT Suspension
2.2. Organisms
2.3. Interaction of Agglomerated SWCNTS with the Marine Green Algae, T. Suecica under Exposure Conditions
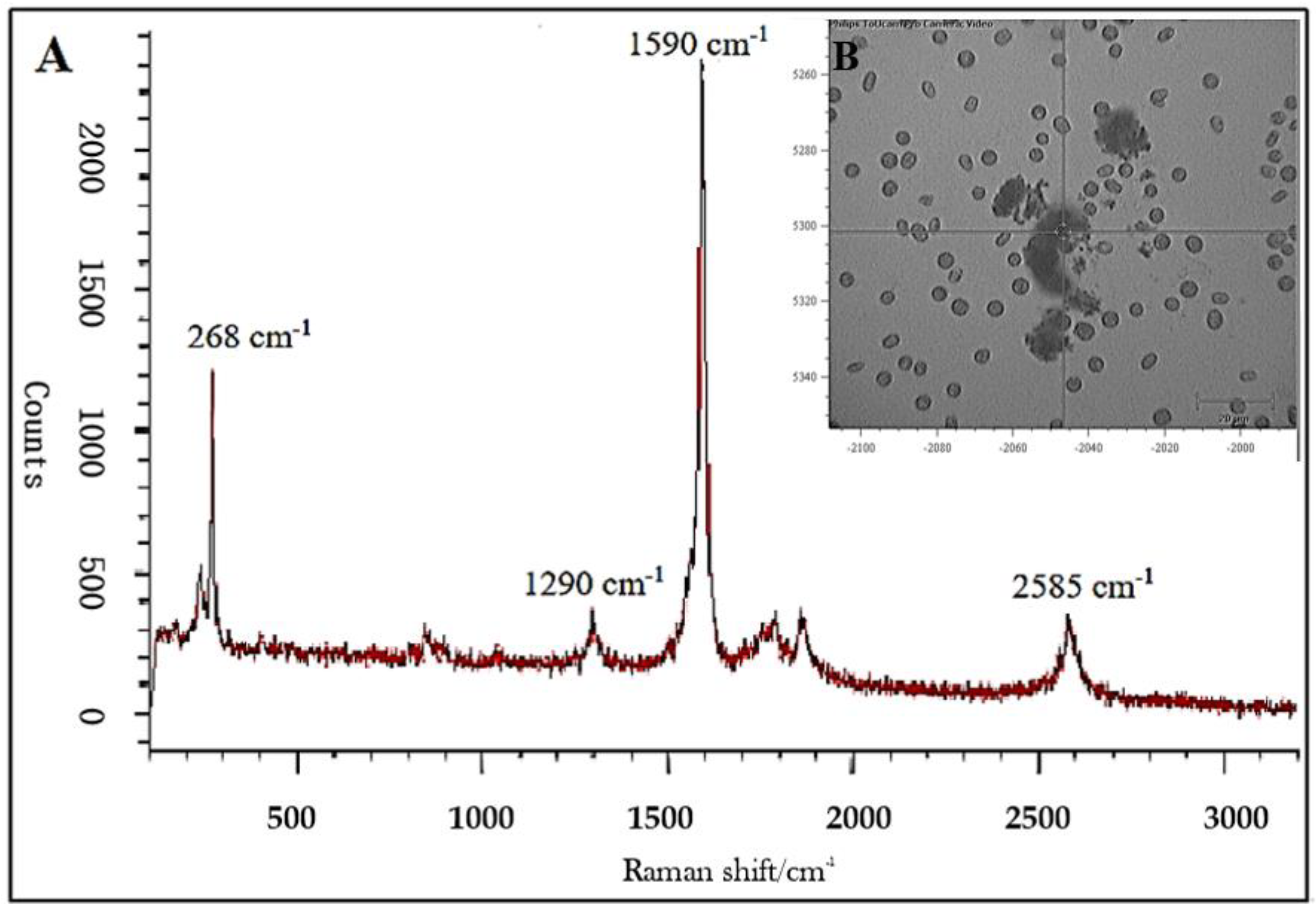
2.4. Raman Microspectroscopy
2.5. Scanning Electron Microscope (SEM)
2.6. Transmission Electron Microscopy (TEM)
2.7. Algal Growth Rates and Abundance
2.8. Cell Viability
2.9. Pseudofaecal Algal Cells and SWCNTs
2.10. Trophic Transfer of SWCNTs from Algae to Mussels
2.11. Toxicology
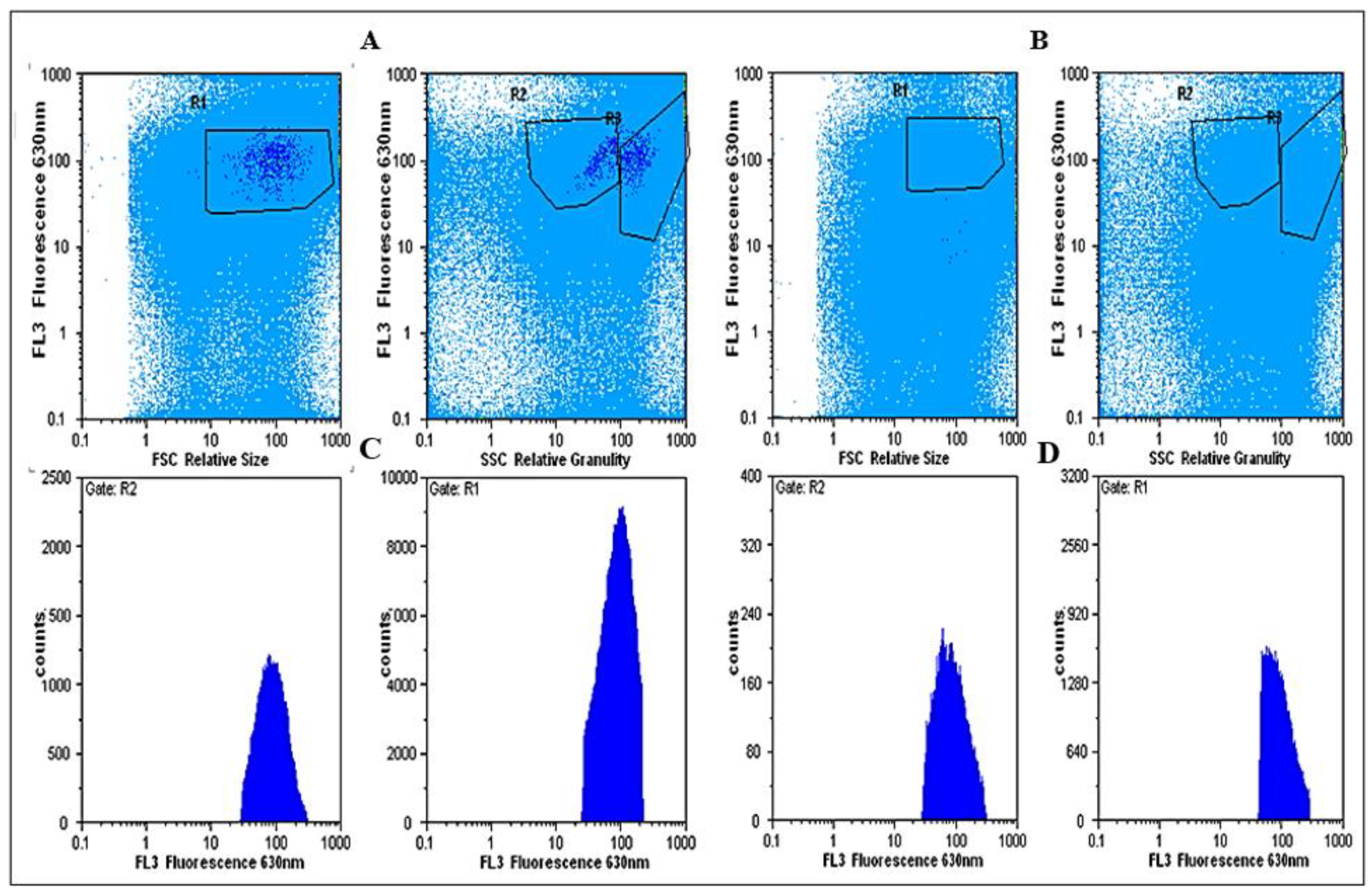
2.11.1. Cell Isolation
2.11.2. Comet Assay
2.11.3. Oxidative Stress
2.11.4. Data Analysis
3. Results and Discussions
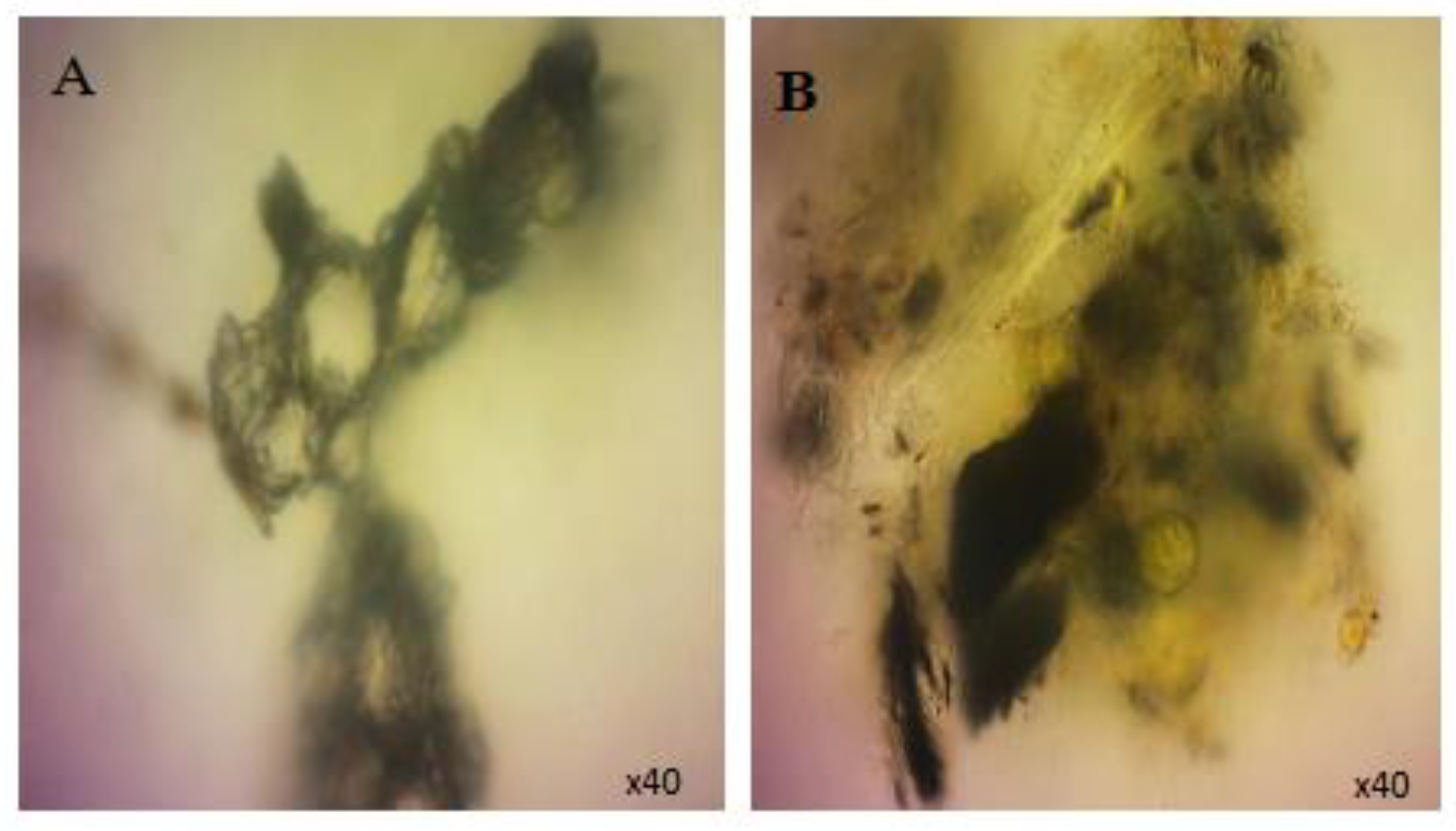
3.1. SWCNT Interaction with Algae
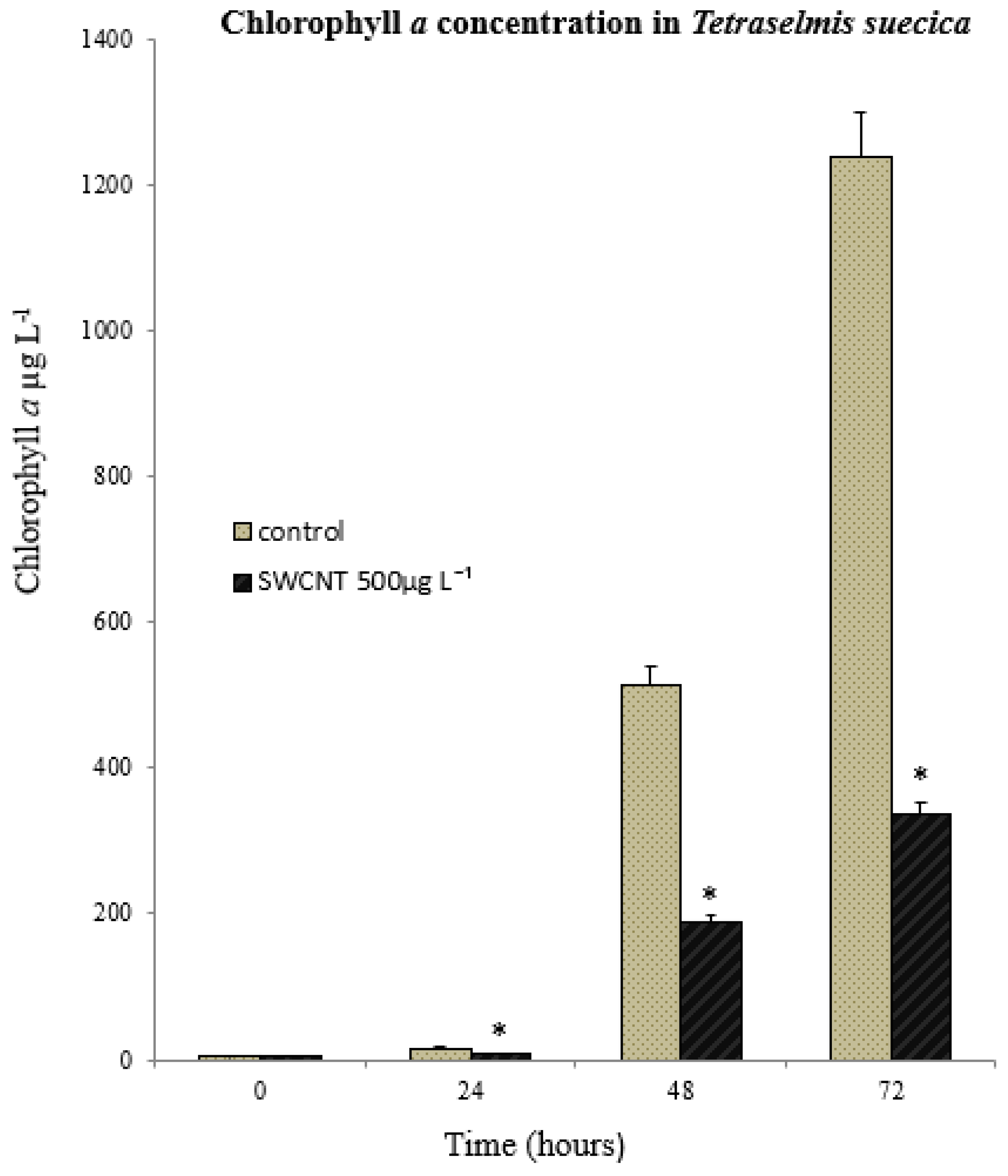
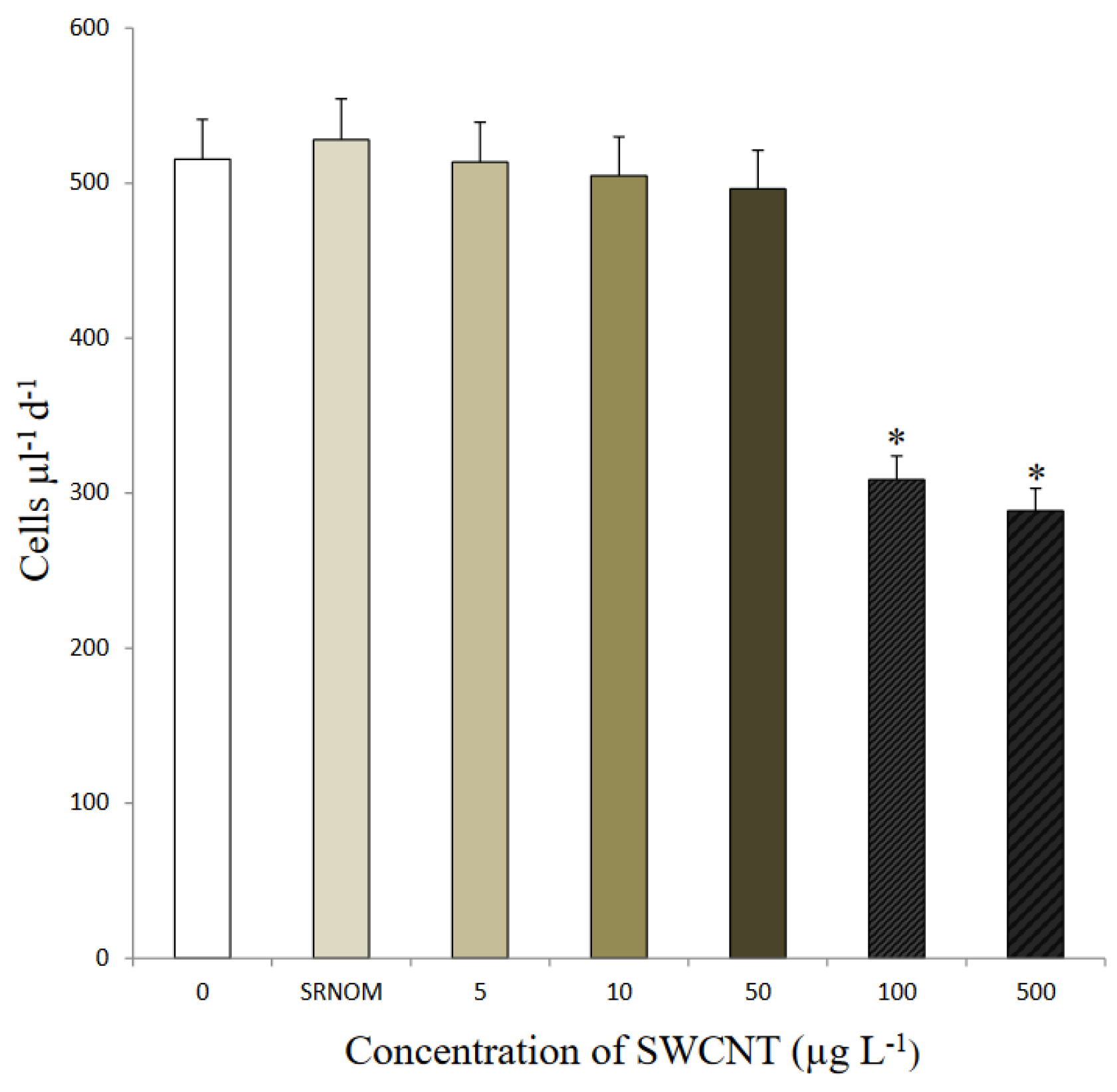
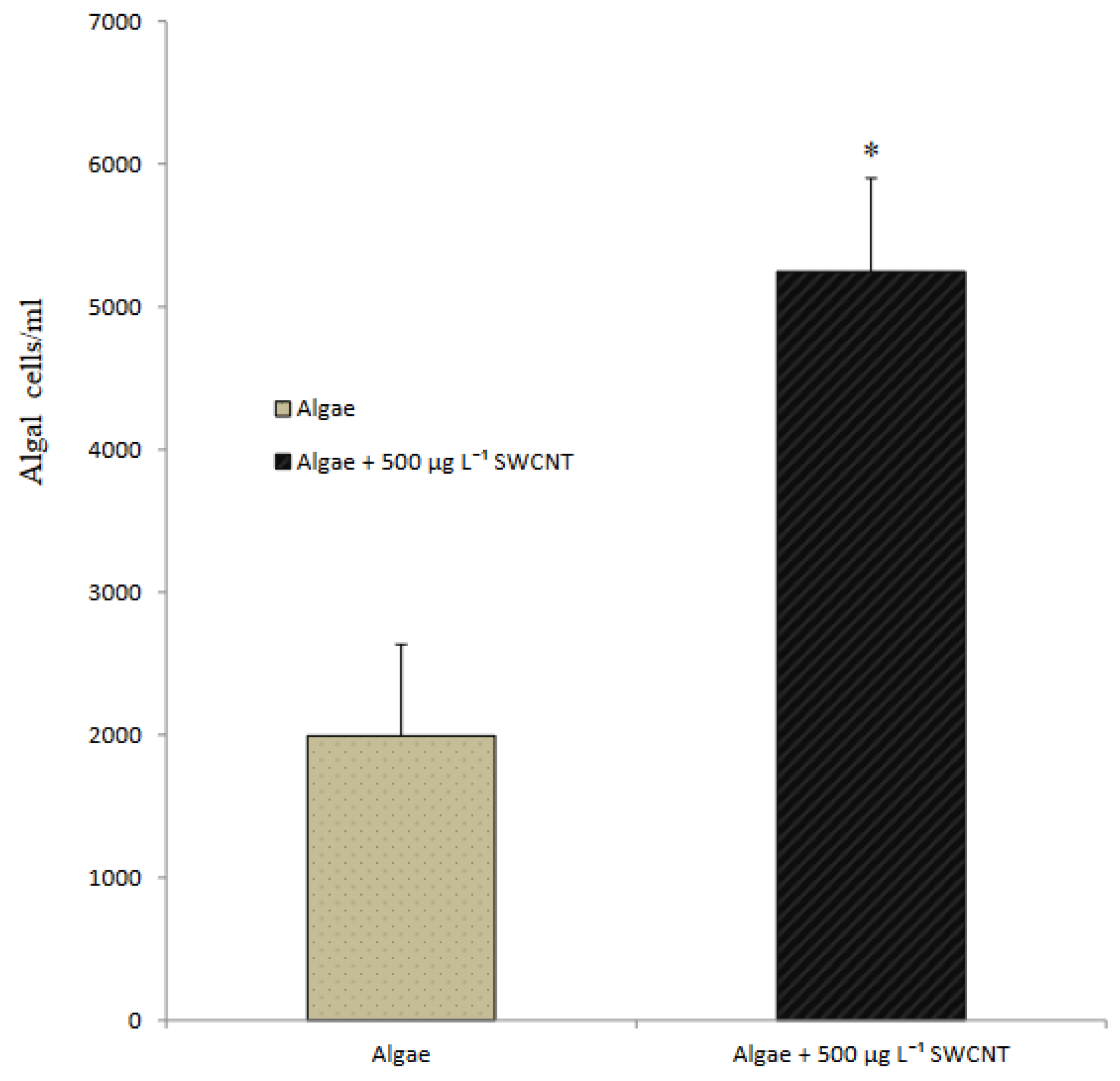
3.2. Toxicological Response of Algae to SWCNT Exposure
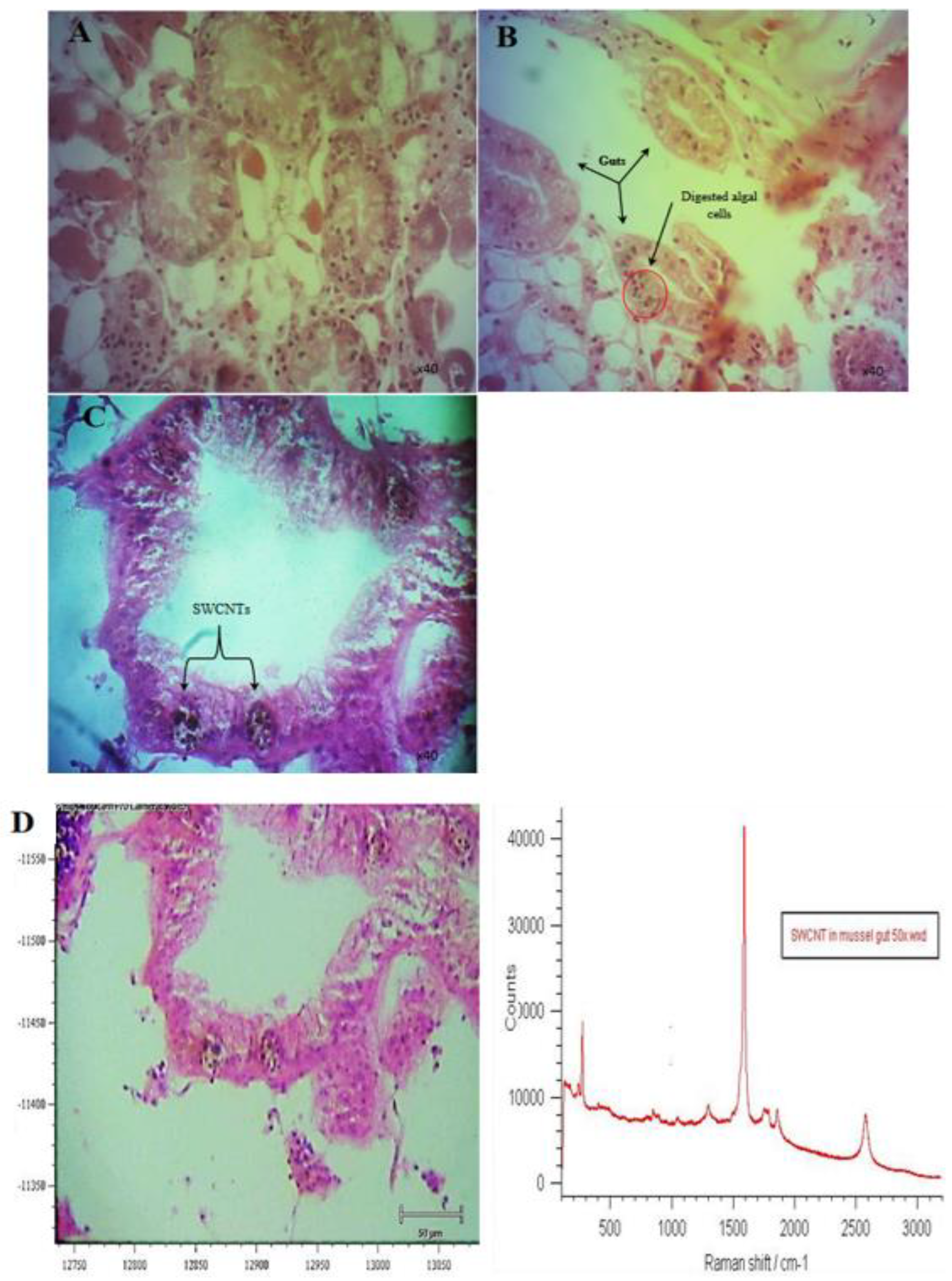
3.3. Response of Mussels to SWCNT Contaminated Algae
3.4. Trophic Transfer of SWCNTs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petosa, A.R.; Jaisi, D.P.; Quevedo, I.R.; Elimelech, M.; Tufenkji, N. Aggregation and Deposition of Engineered Nanomaterials in Aquatic Environments: Role of Physicochemical Interactions. Environ. Sci. Technol. 2010, 44, 6532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquivel, E.; Murr, L.A. TEM analysis of nanoparticulates in a polar ice core. Mater. Charact. 2004, 52, 15–25. [Google Scholar] [CrossRef]
- Navarro, E.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.; Quigg, A.; Santschi, P.H.; Sigg, L. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luther, W.G., III; Rickard, T.D. Metal sulfide cluster complexes and their biogeochemical importance in the environment. J. Nanoparticle Res. 2005, 7, 389–407. [Google Scholar] [CrossRef]
- Aitken, J. XV.—On the formation of small clear spaces in dusty air. Earth Environ. Sci. Trans. R. Soc. Edinb. 1884, 32, 239–272. [Google Scholar] [CrossRef] [Green Version]
- Biswas, P.; Wu, Y.C. Nanoparticles and the environment. J. Air Waste Manag. Assoc. 2005, 55, 708–746. [Google Scholar] [CrossRef] [PubMed]
- Deudero, S.; Cabanellas, M.; Blanco, A.; Tejada, S. Stable isotope fractionation in the digestive gland, muscle and gills tissues of the marine mussel Mytilus galloprovincialis. J. Exp. Mar. Biol. Ecol. 2009, 368, 181–188. [Google Scholar] [CrossRef]
- Petersen, E.J.; Flores-Cervantes, D.X.; Bucheli, T.D.; Elliott, L.C.C.; Fagan, J.A.; Gogos, A.; Hanna, S.; Kägi, R.; Mansfield, E.; Bustos, A.R.M.; et al. Quantification of Carbon Nanotubes in Environmental Matrices: Current Capabilities, Case Studies, and Future Prospects. Environ. Sci. Technol. 2016, 50, 4587–4605. [Google Scholar] [CrossRef]
- Ren, X.; Chen, C.; Nagatsu, M.; Wang, X. Carbon nanotubes as adsorbents in environmental pollution management: A review. Chem. Eng. J. 2011, 170, 395–410. [Google Scholar] [CrossRef]
- Al-Shaeri, M.; Ahmed, D.; McCluskey, F.; Turner, G.; Paterson, L.; Dyrynda, E.A.; Hartl, M.G. Potentiating toxicological interaction of single-walled carbon nanotubes with dissolved metals. Environ. Toxicol. Chem. 2013, 32, 2701–2710. [Google Scholar] [CrossRef]
- Choi, S.-J.; Kwon, T.; Im, H.; Moon, D.-I.; Baek, D.J.; Seol, M.-L.; Duarte, J.P.; Choi, Y.-K. A Polydimethylsiloxane (PDMS) Sponge for the Selective Absorption of Oil from Water. ACS Appl. Mater. Interfaces 2011, 3, 4552–4556. [Google Scholar] [CrossRef] [PubMed]
- Mahdavian, L. Investigation of Polychlorinated Biphenyls (Pcbs) Removal by Signal Walled Carbon Nanotubes: Studies. J. Theor. Comput. Chem. 2011, 10, 651–657. [Google Scholar] [CrossRef]
- Gottschalk, F.; Nowack, B. The release of engineered nanomaterials to the environment. J. Environ. Monit. 2011, 13, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.S.-J. Contamination and Release of Nanomaterials Associated with the Use of Personal Protective Clothing. Ann. Occup. Hyg. 2015, 59, 491–503. [Google Scholar] [PubMed]
- Pereira, F.F.; Ferreira, M.D.; Jonsson, C.M.; de Jesus, K.R.; de Castro, V.L.S.; Correa, D.S. Toxicity of Engineered Nanostructures in Aquatic Environments. In Nanotoxicology and Nanoecotoxicology; Springer: Cham, Switzerland, 2021; Volume 1, pp. 171–202. [Google Scholar]
- Saxena, P.; Sangela, V.; Ranjan, S.; Dutta, V.; Dasgupta, N.; Phulwaria, M.; Rathore, D.S. Aquatic nanotoxicology: Impact of carbon nanomaterials on algal flora. Energy Ecol. Environ. 2020, 5, 240–252. [Google Scholar] [CrossRef]
- Wigger, H.; Hackmann, S.; Zimmermann, T.; Köser, J.; Thöming, J.; von Gleich, A. Influences of use activities and waste management on environmental releases of engineered nanomaterials. Sci. Total Environ. 2015, 535, 160–171. [Google Scholar] [CrossRef]
- Buzea, C.; Pacheco, I.I.; Robbie, K. Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases 2007, 2, MR17–MR71. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Q.; Pan, C.; Chu, W.; Vipin, A.K.; Sun, L. Environmental Remediation Applications of Carbon Nanotubes and Graphene Oxide: Adsorption and Catalysis. Nanomaterials 2019, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Kroll, A.; Kühnel, D.; Schirmer, K. Testing Nanomaterial Toxicity in Unicellular Eukaryotic Algae and Fish Cell Lines. In Oxidative Stress and Nanotechnology, Methods in Molecular Biology; Armstrong, D., Bharali, D., Eds.; Humana Press: Totowa, NJ, USA, 2013. [Google Scholar]
- Borowitzka, M.A. Microalgae for aquaculture: Opportunities and constraints. J. Appl. Phycol. 1997, 9, 393–401. [Google Scholar] [CrossRef]
- Hund-Rinke, K.; Simon, M. Ecotoxic effect of photocatalytic active nanoparticles (TiO2) on algae and daphnids. Environ. Sci. Pollut. Res. 2006, 13, 225–232. [Google Scholar] [CrossRef]
- Kwok, K.W.; Leung, K.M.; Flahaut, E.; Cheng, J.; Cheng, S.H. Chronic toxicity of double-walled carbon nanotubes to three marine organisms: Influence of different dispersion methods. Nanomedicine 2010, 6, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.; Joo, S.H.; Su, C.M.; Toborek, M. Toxicity of ZnO/TiO2-conjugated carbon-based nanohybrids on the coastal marine alga Thalassiosira pseudonana. Environ. Toxicol. 2020, 35, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pikula, K.; Chaika, V.; Zakharenko, A.; Markina, Z.; Vedyagin, A.; Kuznetsov, V.; Gusev, A.; Park, S.; Golokhvast, K. Comparison of the Level and Mechanisms of Toxicity of Carbon Nanotubes, Carbon Nanofibers, and Silicon Nanotubes in Bioassay with Four Marine Microalgae. Nanomaterials 2020, 10, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakkar, M. Comparative toxicity effects of carboxylated carbon nanotubes to fresh water and marine algae. In Abstracts of Papers of the American Chemical Society; American Chemical Society: Washington, DC, USA, 2016; Volume 252. [Google Scholar]
- Zhang, C.; Chen, X.H.; Tan, L.J.; Wang, J.T. Combined toxicities of copper nanoparticles with carbon nanotubes on marine microalgae Skeletonema costatum. Environ. Sci. Pollut. Res. 2018, 25, 13127–13133. [Google Scholar] [CrossRef] [PubMed]
- Fabrega, J.; Fawcett, S.R.; Renshaw, J.C.; Lead, J.R. Silver nanoparticle impact on bacterial growth: Effect of pH, concentration, and organic matter. Environ. Sci. Toxicol. 2009, 43, 7285–7290. [Google Scholar] [CrossRef]
- Schwab, F.; Bucheli, T.D.; Lukhele, L.P.; Magrez, A.; Nowack, B.; Sigg, L.; Knauer, K. Are carbon nanotube effects on green algae caused by shading and agglomeration? Environ. Sci. Technol. 2011, 45, 6136–6144. [Google Scholar] [CrossRef]
- Youn, S.; Wang, R.; Gao, J.; Hovespyan, A.; Ziegler, K.J.; Bonzongo, J.-C.J.; Bitton, G. Mitigation of the impact of single-walled carbon nanotubes on a freshwater green algae: Pseudokirchneriella subcapitata. Nanotoxicology 2012, 6, 161–172. [Google Scholar] [CrossRef]
- Politowski, I.; Regnery, P.; Hennig, M.P.; Siebers, N.; Ottermanns, R.; Schäffer, A. Fate of weathered multi-walled carbon nanotubes in an aquatic sediment system. Chemosphere 2021, 277, 130319. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Egede, P.P.; Saavedra, I.B. Feeding Behaviour of the Mussel, Mytilus edulis: New Observations, with a Minireview of Current Knowledge. J. Mar. Biol. 2011, 2011, 312459. [Google Scholar] [CrossRef] [Green Version]
- SEPA. State of Scotland’s Environment; Scottish Environment Protection Agency: Stirling, UK, 2006. [Google Scholar]
- Guiselin, N.; Courcot, L.; Artigas, L.F.; Le Jéloux, A.; Brylinski, J.M. An optimised protocol to prepare Phaeocystis globosa morphotypes for scanning electron microscopy observation. J. Microbiol. Methods 2009, 77, 119–123. [Google Scholar] [CrossRef]
- Azma, M.; Mohamad, R.; Rahim, R.A.; Ariff, A.B. Improved protocol for the preparation of Tetraselmis suecica axenic culture and adaptation to heterotrophic cultivation. Open Biotechnol. J. 2010, 4, 36–46. [Google Scholar] [CrossRef]
- Lobban, C.; Chapman, D.; Kremer, B. Experimental Phycology: A Laboratory Manual; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Wei, L.; Thakkar, M.; Chen, Y.; Ntim, S.A.; Mitra, S.; Zhang, X. Cytotoxicity effects of water dispersible oxidized multiwalled carbon nanotubes on marine alga, Dunaliella tertiolecta. Aquat. Toxicol. 2010, 100, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Wikfors, G.H.; Patterson, G.W.; Ghosh, P.; Lewin, R.A.; Smith, B.C.; Alix, J.H. Growth of post-set oysters, Crassostrea virginica, on high-lipid strains of algal flagellates Tetraselmis spp. Aquaculture 1996, 143, 411–419. [Google Scholar] [CrossRef]
- Petrescu, C.M.; Turcus, V.; Bratosin, D. Toxicity evaluation by flow cytometric analysis of nanoparticles using the unicellular alga Chlorella. Stud. Univ. Vasile Goldis Arad. Ser. Stiint. Vietii (Life Sci. Ser.) 2013, 23, 381. [Google Scholar]
- Kumar, G.; Degheidy, H.; Casey, B.J.; Goering, P.L. Flow cytometry evaluation of in vitro cellular necrosis and apoptosis induced by silver nanoparticles. Food Chem. Toxicol. 2015, 85, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Muller-Feuga, A.; Moal, J.; Kaas, R. Live Feeds in Marine Aquaculture; Støttrup, J.G., McEvoy, L.A., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2007; p. 253. [Google Scholar]
- Cucci, T.; Shumway, S.; Newell, R.; Selvin, R.; Guillard, R.; Yentsch, C. Flow cytometry: A new method for characterization of differential ingestion, digestion and egestion by suspension feeders. Mar. Ecol. Prog. Ser. 1985, 24, 201–204. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Hyun, C.-K. Comparative evaluation of the alkaline comet assay with the micronucleus test for genotoxicity monitoring using aquatic organisms. Ecotoxicol. Environ. Saf. 2006, 64, 288–297. [Google Scholar] [CrossRef]
- Coughlan, B.M.; Hartl, M.G.J.; O’Reilly, S.J.; Sheehan, D.; Morthersill, C.; van Pelt, F.N.A.M.; O’Halloran, J.; O’Brien, N.M. Detecting genotoxicity using the Comet assay following chronic exposure of Manila clam Tapes semidecussatus to polluted estuarine sediments. Mar. Pollut. Bull. 2002, 44, 1359–1365. [Google Scholar] [CrossRef]
- Singh, R.; Hartl, M.G.J. Fluctuating estuarine conditions are not confounding factors for the Comet assay assessment of DNA damage in the mussel Mytilus edulis. Ecotoxicology 2012, 21, 1998–2003. [Google Scholar] [CrossRef]
- Hartl, M.G.J.; Grigson, S.; Sinet, E. Maintenance of bivalve haemocytes for the purpose of delayed DNA strand break assessment using the Comet assay. Environ. Mol. Mutagen. 2010, 51, 64–68. [Google Scholar]
- Smith, C.J.; Shaw, B.J.; Handy, R.D. Toxicity of single walled carbon nanotubes to rainbow trout (Oncorhynchus mykiss): Respiratory toxicity, organ pathologies, and other physiological effects. Aquat. Toxicol. 2007, 82, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Lovell, D.P. The Comet Assay in Toxicology; Dhawan, A., Anderson, D., Eds.; Royal Society of Chemistry: Cambridge, UK, 2009; pp. 424–450. [Google Scholar]
- Sparks, T. (Ed.) Statistics in Ecotoxicology; John Wiley & Sons: Chichester, UK, 2000. [Google Scholar]
- Chinnapongse, S.L.; MacCuspie, R.I.; Hackley, V.A. Persistence of singly dispersed silver nanoparticles in natural freshwaters, synthetic seawater, and simulated estuarine waters. Sci. Total Environ. 2011, 409, 2443–2450. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, J.S.; Hyeok An, K.; Lee, K.; Kim, D.Y.; Bae, D.J.; Lee, Y.H. Electrophoretic and Dynamic Light Scattering in Evaluating Dispersion and Size Distribution of Single-Walled Carbon Nanotubes. J. Nanosci. Nanotechnol. 2005, 5, 1045–1049. [Google Scholar] [CrossRef] [Green Version]
- Dillon, A.C.; Gennett, T.; Jones, K.M.; Alleman, J.L.; Parilla, P.A.; Heben, M.J. A Simple and Complete Purification of Single-Walled Carbon Nanotube Materials. Adv. Mater. 1999, 11, 1354–1358. [Google Scholar] [CrossRef]
- Long, Z.; Ji, J.; Yang, K.; Lin, D.; Wu, F. Systematic and Quantitative Investigation of the Mechanism of Carbon Nanotubes’ Toxicity toward Algae. Environ. Sci. Technol. 2012, 46, 8458–8466. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tabakman, S.; Welsher, K.; Dai, H. Carbon nanotubes in biology and medicine: In vitro and in vivo detection, imaging and drug delivery. Nano Res. 2009, 2, 85–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Gladue, R.; Hansen, J.; Wojnar, C.; Chalmers, J.J. Growth inhibition of dinoflagellate algae in shake flasks: Not due to shear this time! Biotechnol. Prog. 2010, 26, 79–87. [Google Scholar] [CrossRef]
- Baun, A.; Hartmann, N.B.; Grieger, K.; Kusk, K.O. Ecotoxicity of engineered nanoparticles to aquatic invertebrates: A brief review and recommendations for future toxicity testing. Ecotoxicology 2008, 17, 387–395. [Google Scholar] [CrossRef]
- Fabrega, J.; Luoma, S.N.; Tyler, C.R.; Galloway, T.S.; Lead, J.R. Silver nanoparticles: Behaviour and effects in the aquatic environment. Environ. Int. 2011, 37, 517–531. [Google Scholar] [CrossRef]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar] [PubMed]
- Ghafari, P.; St-Denis, C.H.; Power, M.E.; Jin, X.; Tsou, V.; Mandal, H.S.; Bols, N.C.; Tang, X. Impact of carbon nanotubes on the ingestion and digestion of bacteria by ciliated protozoa. Nat. Nanotechnol. 2008, 3, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Vanderploeg, H.A.; Liebig, J.R.; Carmichael, W.W.; Agy, M.A.; Johengen, T.H.; Fahnenstiel, G.L.; Nalepa, T.F. Zebra mussel (Dreissena polymorpha) selective filtration promoted toxic Microcystis blooms in Saginaw Bay (Lake Huron) and Lake Erie. Can. J. Fish. Aquat. Sci. 2001, 58, 1208–1221. [Google Scholar] [CrossRef]
- Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Fernandes, T.F.; Handy, R.D.; Lyon, D.Y.; Mahendra, S.; McLaughlin, M.J.; Lead, J.R. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ. Toxicol. Chem. 2008, 27, 1825–1851. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, J.; Zhang, X.; Chang, Y.; Chen, Y. Trophic transfer of TiO nanoparticles from daphnia to zebrafish in a simplified freshwater food chain. Chemosphere 2010, 79, 928–933. [Google Scholar] [CrossRef]
- Werlin, R.; Priester, J.H.; Mielke, R.E.; Krämer, S.; Jackson, S.; Stoimenov, P.K.; Stucky, G.D.; Cherr, G.N.; Orias, E.; Holden, P.A. Biomagnification of cadmium selenide quantum dots in a simple experimental microbial food chain. Nat. Nanotechnol. 2011, 6, 65–71. [Google Scholar] [CrossRef]
- Lowry, G.V.; Hotze, E.M.; Bernhardt, E.S.; Dionysiou, D.D.; Pedersen, J.A.; Wiesner, M.R.; Xing, B. Environmental occurrences, behaviour, fate, and ecological effects of nanomaterials: An introduction to the special series. J. Environ. Qual. 2010, 39, 1867–1874. [Google Scholar] [CrossRef] [Green Version]
- Farré, M.; Gajda-Schrantz, K.; Kantiani, L.; Barceló, D. Ecotoxicity and analysis of nanomaterials in the aquatic environment. Anal. Bioanal. Chem. 2009, 393, 81–95. [Google Scholar] [CrossRef]
- Holbrook, R.D.; Murphy, K.E.; Morrow, J.B.; Cole, K.D. Trophic transfer of nanoparticles in a simplified invertebrate food web. Nat. Nanotechnol. 2008, 3, 352–355. [Google Scholar] [CrossRef]
- Xiaojun, Z.; Yanghui, X.; Qiang, H.; Zhengsong, W.; Ruixing, H.; Jiahui, S.; Xiaoliu, H. Effects of Nanoparticles on Growth of Freshwater Algae: Toxicity Mechanism and Combined Toxicity. Asian J. Ecotoxicol. 2021, 3, 128–143. [Google Scholar]
- Shi, Q.; Wang, C.L.; Zhang, H.; Chen, C.; Zhang, X.; Chang, X.-L. Trophic transfer and biomagnification of fullerenol nanoparticles in an aquatic food chain. Environ. Sci. Nano 2020, 7, 1240–1251. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SWCNT (µg L−1) | Zeta Potential | DLS (nm) |
|---|---|---|
| 5 | −8.84 | 475 |
| 10 | −10.83 | 1384 |
| 50 | −10.13 | 1740 |
| 100 | −15.93 | 4982 |
| 500 | −13.73 | 6206 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shaeri, M.; Paterson, L.; Stobie, M.; Cyphus, P.; Hartl, M.G.J. Trophic Transfer of Single-Walled Carbon Nanotubes at the Base of the Food Chain and Toxicological Response. Nanomaterials 2022, 12, 4363. https://doi.org/10.3390/nano12244363
Al-Shaeri M, Paterson L, Stobie M, Cyphus P, Hartl MGJ. Trophic Transfer of Single-Walled Carbon Nanotubes at the Base of the Food Chain and Toxicological Response. Nanomaterials. 2022; 12(24):4363. https://doi.org/10.3390/nano12244363
Chicago/Turabian StyleAl-Shaeri, Majed, Lynn Paterson, Margret Stobie, Paul Cyphus, and Mark G. J. Hartl. 2022. "Trophic Transfer of Single-Walled Carbon Nanotubes at the Base of the Food Chain and Toxicological Response" Nanomaterials 12, no. 24: 4363. https://doi.org/10.3390/nano12244363
APA StyleAl-Shaeri, M., Paterson, L., Stobie, M., Cyphus, P., & Hartl, M. G. J. (2022). Trophic Transfer of Single-Walled Carbon Nanotubes at the Base of the Food Chain and Toxicological Response. Nanomaterials, 12(24), 4363. https://doi.org/10.3390/nano12244363