Rapid Detection of Pathogenic Bacteria by the Naked Eye




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Materials and Reagents
2.2. Instrumentation
2.3. Preparation of Bacterial Samples
2.4. Endogenous Peroxidase and Catalase Reactions of Bacteria
2.5. A Two-Step Method for Distinguishing Gram-Positive from Gram-Negative Bacteria
2.6. Using Cherry Tomatoes as the Simulated Real Sample
2.7. Detection of Bacteria from Door Handles
3. Results and Discussion
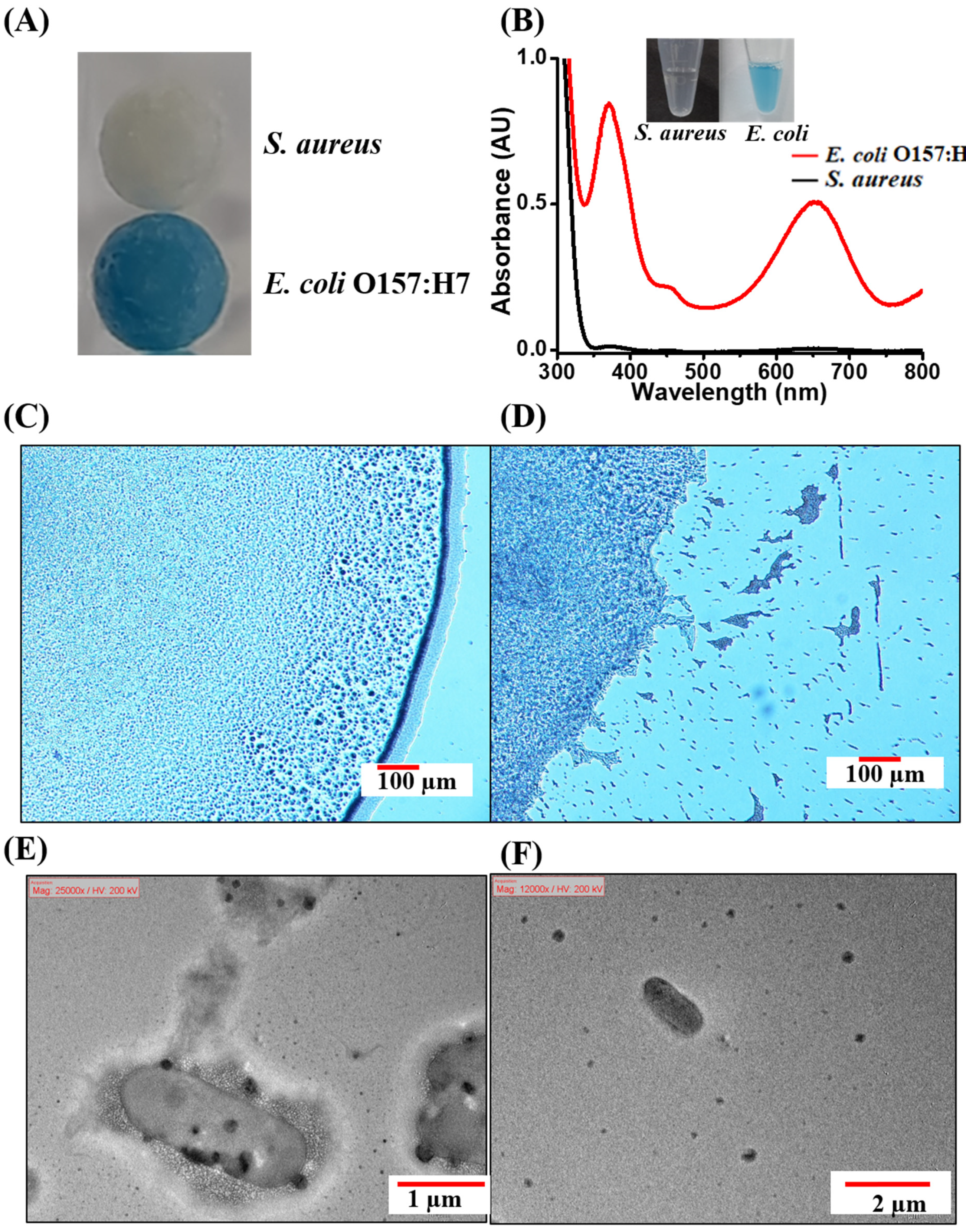
3.1. Endogenous Peroxidase Reactions Derived from Bacterial Samples
3.2. Optimization of Experimental Parameters
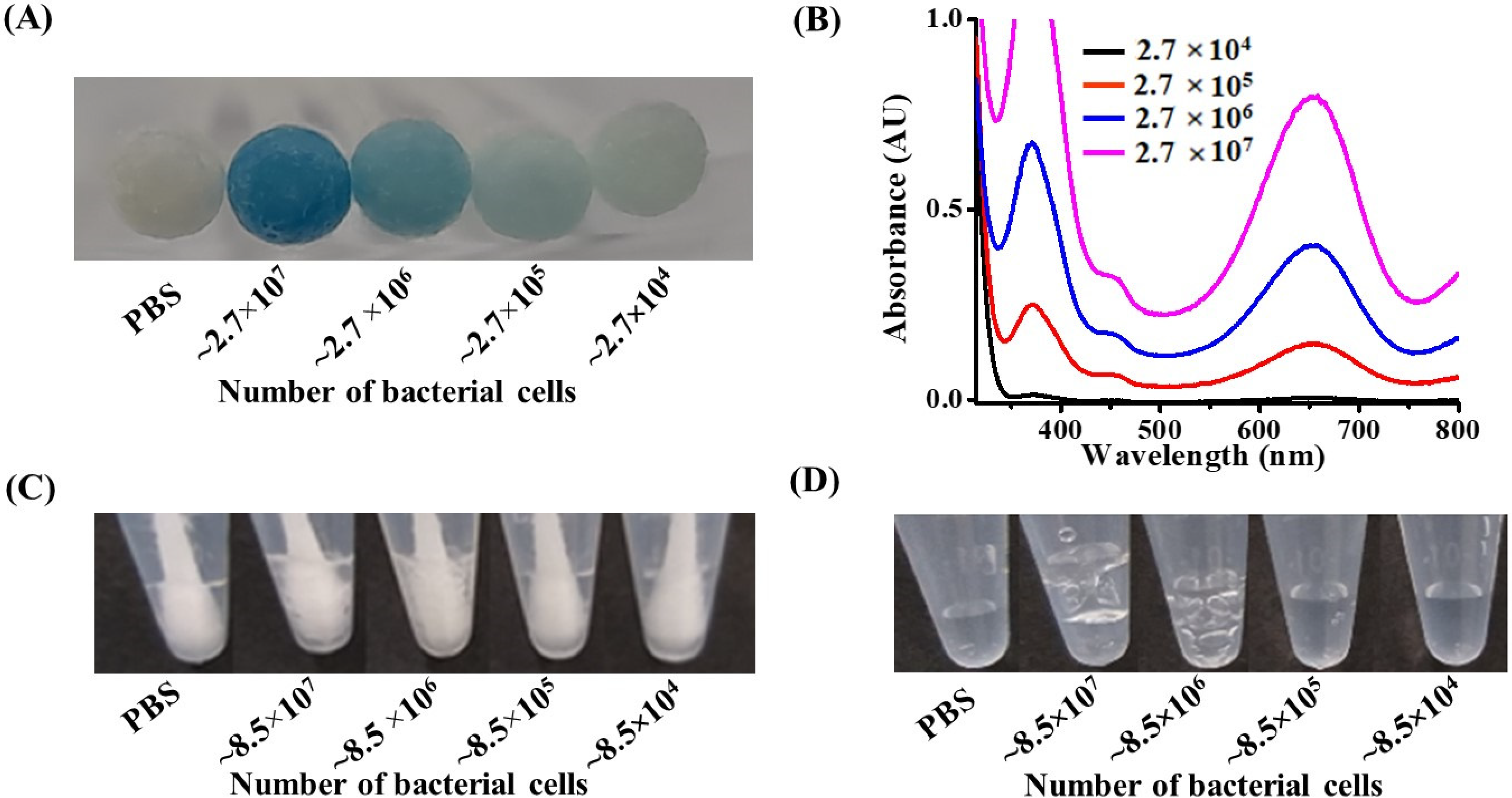
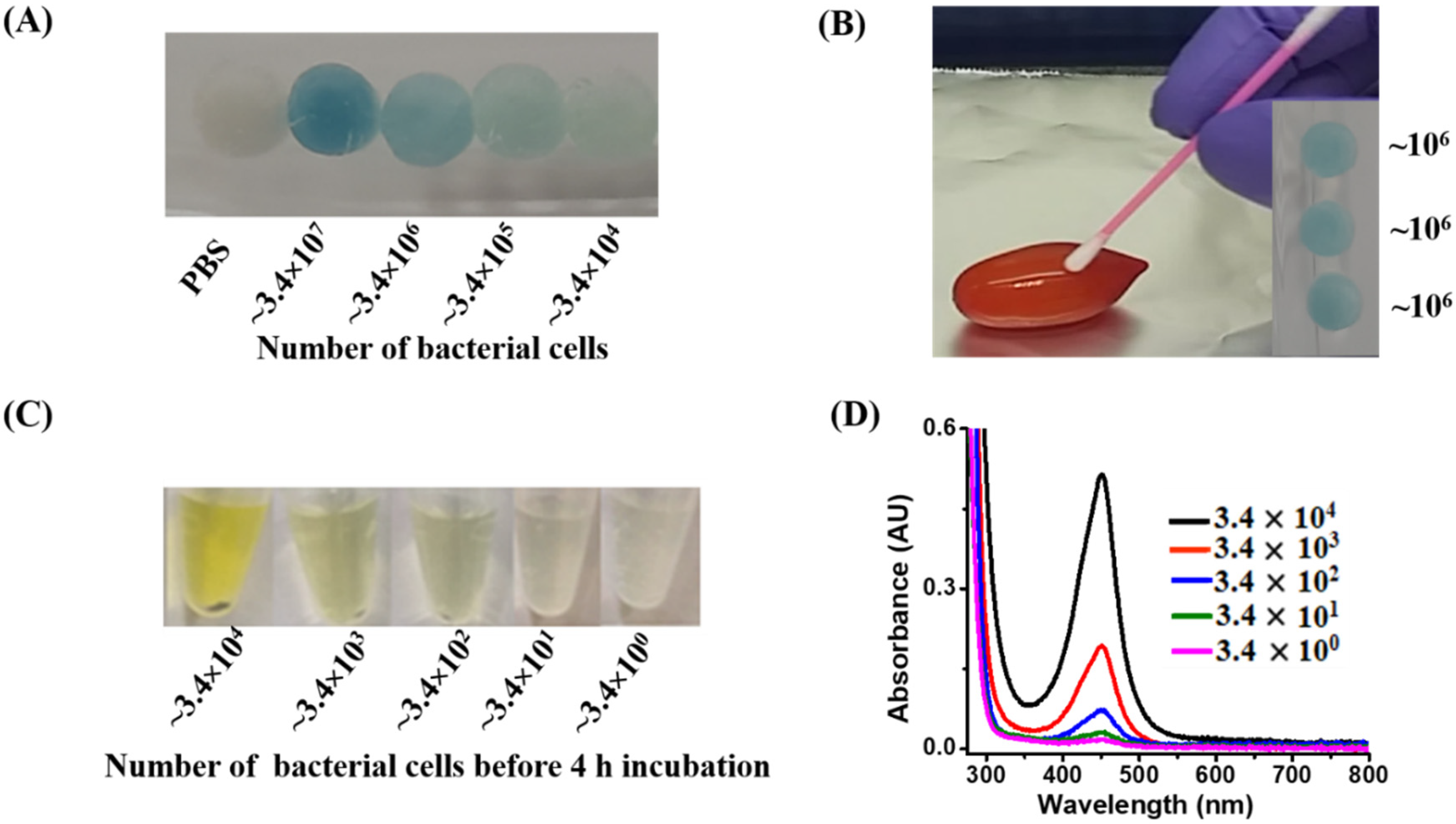
3.3. Examination of the Lowest Detectable Concentration by the Naked Eye
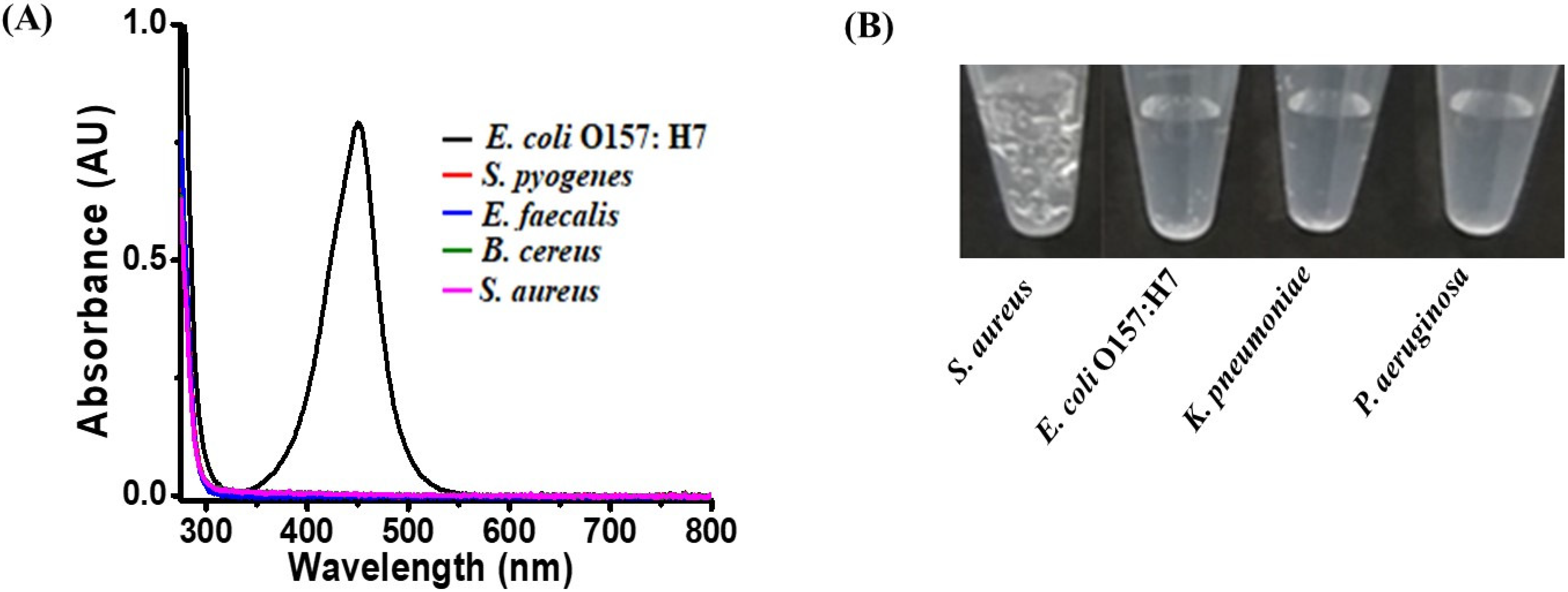
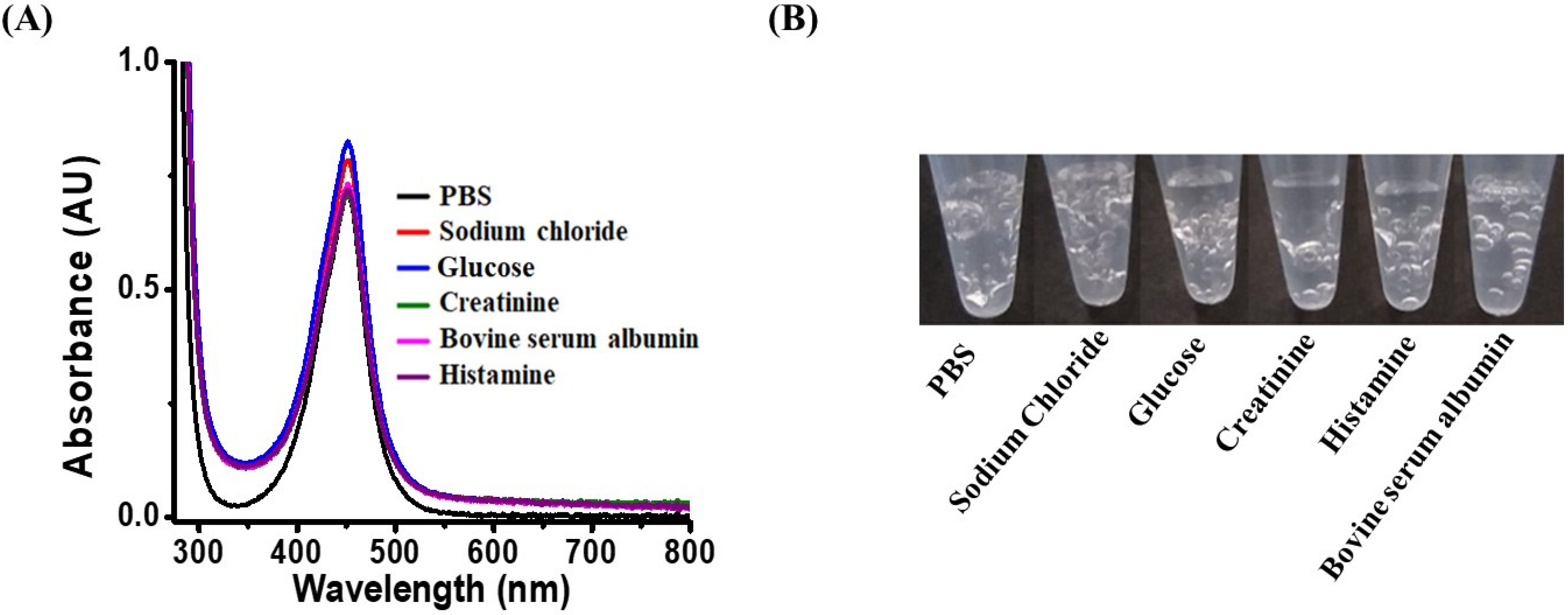
3.4. Examination of Selectivity
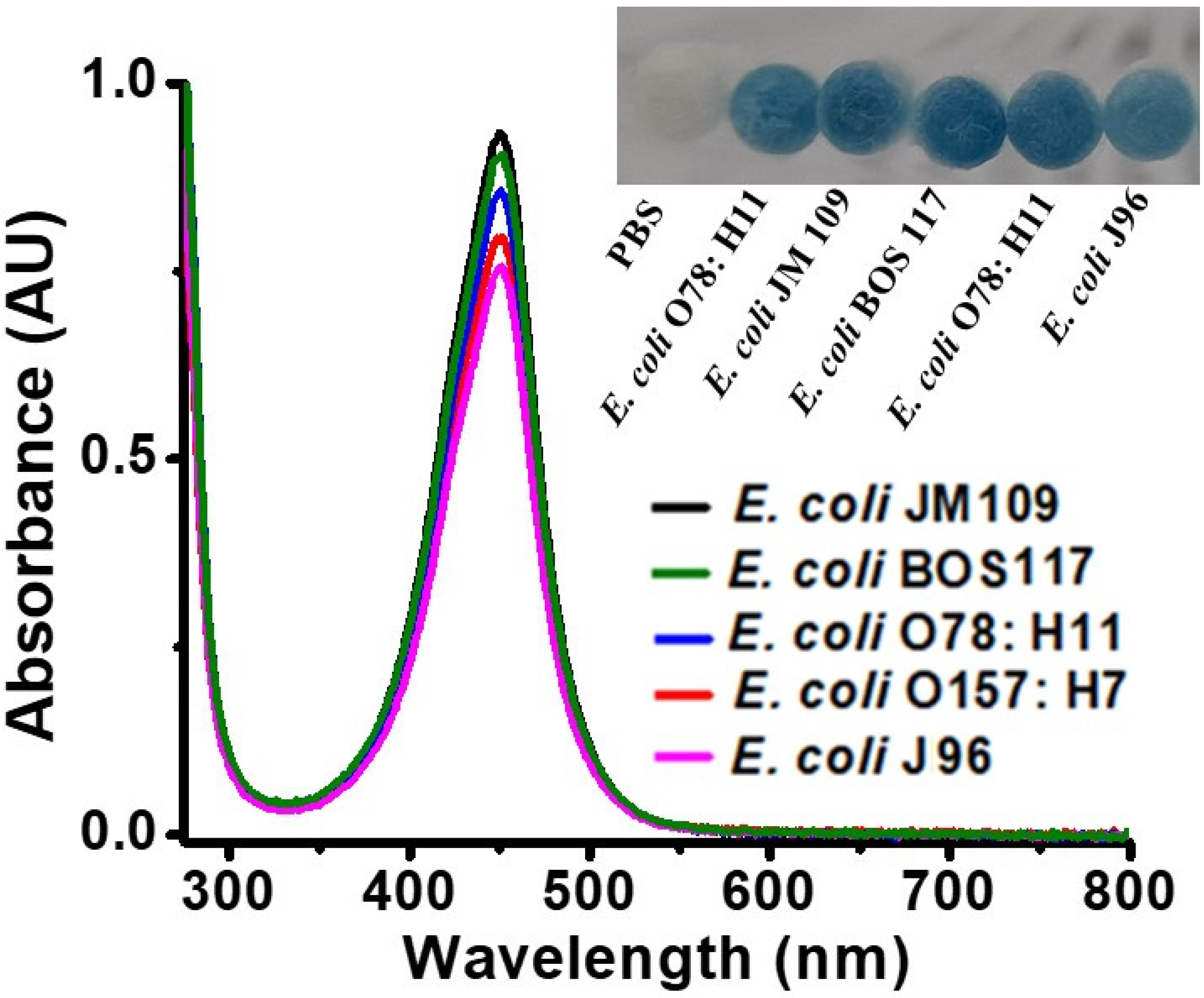
3.5. Analysis of Different Strains of E. coli
3.6. Examination of Interference Effects
3.7. Analysis of Real Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607. [Google Scholar] [CrossRef]
- FDA. Bad Bug Book, Foodborne Pathogenic Microorganisms and Natural Toxins, 2nd ed.; FDA: Silver Spring, MD, USA, 2012.
- Boyer, O.; Niaudet, P. Hemolytic uremic syndrome: New developments in pathogenesis and treatment. Int. J. Nephrol. 2011, 2011, 908407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Guevara, J.J.; Arenas-Hernandez, M.M.P.; Martínez de la Peña, C.; Silva, J.L.; Luna-Guevara, M.L. The role of pathogenic E. coli in fresh vegetables: Behavior, contamination factors, and preventive measures. Int. J. Microbiol. 2019, 2019, 2894328. [Google Scholar] [CrossRef] [Green Version]
- Heiman, K.E.; Mody, R.K.; Johnson, S.D.; Griffin, P.M.; Gould, L.H. Escherichia coli O157 outbreaks in the United States, 2003–2012. Emerg. Infect. Dis. 2015, 21, 1293–1301. [Google Scholar] [CrossRef]
- Nygren, B.L.; Schilling, K.A.; Blanton, E.M.; Silk, B.J.; Cole, D.J.; Mintz, E.D. Foodborne outbreaks of shigellosis in the USA, 1998–2008. Epidemiol. Infect. 2013, 141, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.L.; Gorris, L.G.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Camps, N.; Dominguez, A.; Perez, M.; Pardos, J.; Llobet, T.; Usera, M.A.; Salleras, L. A foodborne outbreak of Salmonella infection due to overproduction of egg-containing foods for a festival. Epidemiol. Infect. 2005, 133, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. Bio. Med. Res. Int. 2014, 2014, 827965. [Google Scholar]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Denis, N.; Zhang, H.; Leroux, A.; Trudel, R.; Bietlot, H. Prevalence and trends of bacterial contamination in fresh fruits and vegetables sold at retail in Canada. Food Control 2016, 67, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Matthysse, A.G. Adherence of bacteria to plant surfaces measured in the laboratory. J. Vis. Exp. 2018, 136, e56599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention National Health Interview Durvey [Data Set]. Available online: http://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 3 May 2021).
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Bloom, D.E.; Cadarette, D. Infectious disease threats in the twenty-first century: Strengthening the global response. Front. Immunol. 2019, 10, 549. [Google Scholar] [CrossRef] [Green Version]
- Käferstein, F.K.; Motarjemi, Y.; Bettcher, D.W. Foodborne disease control: A transnational challenge. Emerg. Infect. Dis. 1997, 3, 503–510. [Google Scholar] [CrossRef]
- Maugeri, G.; Lychko, I.; Sobral, R.; Roque, A.C. Identification and antibiotic-susceptibility profiling of infectious bacterial agents: A review of current and future trends. Biotechnol. J. 2019, 14, 1700750. [Google Scholar] [CrossRef]
- Tong, C.; Li, L.; Xiao, F.; Fan, J.; Zhong, X.; Liu, X.; Liu, B.; Wu, Z.; Zhou, J. Daptomycin and AgNP co-loaded rGO nanocomposites for specific treatment of Gram-positive bacterial infection in vitro and in vivo. Biomater. Sci. 2019, 7, 5097–5111. [Google Scholar] [CrossRef]
- Claeys, K.C.; Heil, E.L.; Pogue, J.M.; Lephart, P.R.; Johnson, J.K. The Verigene dilemma: Gram-negative polymicrobial bloodstream infections and clinical decision making. Diagn. Microbiol. Infect. Dis. 2018, 91, 144–146. [Google Scholar] [CrossRef]
- Bu, S.J.; Wang, K.Y.; Bai, H.S.; Leng, Y.; Ju, C.J.; Wang, C.Y.; Liu, W.S.; Wan, J.Y. Immunoassay for pathogenic bacteria using platinum nanoparticles and a hand-held hydrogen detector as transducer. Application to the detection of Escherichia coli O157: H7. Microchim. Acta 2019, 186, 296. [Google Scholar] [CrossRef]
- Ouyang, Z.; Zhai, Z.; Qin, A.N.; Li, H.; Liu, X.; Qu, X.; Dai, K. Limitations of Gram staining for the diagnosis of infections following total hip or knee arthroplasty. Exp. Ther. Med. 2015, 9, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Froböse, N.J.; Bjedov, S.; Schuler, F.; Kahl, B.C.; Kampmeier, S.; Schaumburg, F. Gram staining: A comparison of two automated systems and manual staining. J. Clin. Microbiol. 2020, 58, e01914-20. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, A.M.; Al-Tawfiq, J.A.; Rabaan, A.A. Misinterpretation of gram stain from the stationary growth phase of positive blood cultures for Brucella and Acinetobacter species. Open Microbiol. J. 2017, 11, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Nóbrega, C.S.; Pauleta, S.R. Reduction of 0.2 in gram-negative bacteria–bacterial peroxidases. Adv. Microb. Physiol. 2019, 74, 415–464. [Google Scholar]
- Ramasamy, M.; Yi, D.K.; An, S.S.A. Enhanced detection sensitivity of Escherichia coli O157: H7 using surface-modified gold nanorods. Int. J. Nanomed. 2015, 10, 179–190. [Google Scholar]
- Wang, X.; Yang, Y.; Li, L.; Sun, M.; Yin, H.; Qin, W. A polymeric liquid membrane electrode responsive to 3, 3′, 5, 5′-tetramethylbenzidine oxidation for sensitive peroxidase/peroxidase mimetic-based potentiometric biosensing. Anal. Chem. 2014, 86, 4416–4422. [Google Scholar] [CrossRef]
- Frey, A.; Meckelein, B.; Externest, D.; Schmidt, M.A. A stable and highly sensitive 3, 3′, 5, 5′-tetramethylbenzidine-based substrate reagent for enzyme-linked immunosorbent assays. J. Immunol. Methods 2009, 233, 47–56. [Google Scholar] [CrossRef]
- John Goka, A.K.; Farthing, M.J. The use of 3, 3′, 5, 5′-tetramethylbenzidine as a peroxidase substrate in microplate enzyme-linked immunosorbent assay. J. Immunoass. 1987, 8, 29–41. [Google Scholar] [CrossRef]
- Chen, Y.; Zhong, Q.; Wang, Y.; Yuan, C.; Qin, X.; Xu, Y. Colorimetric detection of H2O2 and glucose by exploiting the peroxidase-like activity of papain. RSC Adv. 2019, 9, 16566–16570. [Google Scholar] [CrossRef] [Green Version]
- Shim, K.H.; Kang, M.; Kim, M.G.; Chung, B.H.; An, S.S.A. Detection of E. coli O157: H7 using its endogenous active membrane peroxidase. J. Toxicol. Environ. 2011, 3, 80–85. [Google Scholar] [CrossRef]
- Shenge, K.C.; Whong, C.M.; Yakubu, L.L.; Omolehin, R.A.; Erbaugh, J.; Miller, S.A.; LeJeune, J.T. Contamination of tomatoes with coliforms and Escherichia coli on farms and in markets of northwest Nigeria. J. Food Prot. 2015, 78, 57–64. [Google Scholar] [CrossRef]
- Obeng, F.A.; Gyasi, P.B.; Olu-Taiwo, M.; Ayeh-Kumi, F.P. Microbial assessment of tomatoes (Lycopersicon esculentum) sold at some central markets in ghana. Bio. Med. Res. Int. 2018, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeddi, M.Z.; Yunesian, M.; Gorji, M.E.H.; Noori, N.; Pourmand, M.R.; Khaniki, G.R.J. Microbial evaluation of fresh, minimally-processed vegetables and bagged sprouts from chain supermarkets. J. Health Popul. Nutr. 2014, 32, 391–399. [Google Scholar]
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal response to oxidative stress. Front. Cell. Infect. Microbiol. 2012, 2, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukhari, Z.; Weihe, J.; LeChevallier, M. Development of procedures for rapid detection of E. coli O157: H7 from source and finished water samples. Water Sci. Technol. 2004, 50, 33–237. [Google Scholar] [CrossRef]
- Bonham, P.A. Swab cultures for diagnosing wound infections: A literature review and clinical guideline. J. Wound Ostomy Cont. Nurs. 2009, 36, 389–395. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. H2O2 fluxes and compartmentalization inside growing Escherichia coli. J. Bacteriol. 2001, 183, 7182–7189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffellini, S.; Guerrero, S.; Alzamora, S.M. Effect of H2O2 concentration and pH on inactivation kinetics of Escherichia coli. J. Food Saf. 2008, 28, 514–533. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Alkyl hydroperoxide reductase is the primary scavenger of endogenous H2O2 in Escherichia coli. J. Bacteriol. 2001, 183, 7173–7181. [Google Scholar] [CrossRef] [Green Version]
- Frankenberg, L.; Brugna, M.; Hederstedt, L. Enterococcus faecalis heme-dependent Catalase. J. Bacteriol. 2002, 184, 6351–6356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi-Torii, K.; Hayashi, S.; Nakamoto, H.; Nakamura, S. Properties of Aspergillus niger catalase. J. Biochem. 1982, 92, 1449–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, B.; Wilson, D.J.; Feil, E.; Eyre-Walker, A. The distribution of bacterial doubling times in the wild. Proc. Biol. Sci. 2018, 285, 20180789. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandasamy, K.; Jannatin, M.; Chen, Y.-C. Rapid Detection of Pathogenic Bacteria by the Naked Eye. Biosensors 2021, 11, 317. https://doi.org/10.3390/bios11090317
Kandasamy K, Jannatin M, Chen Y-C. Rapid Detection of Pathogenic Bacteria by the Naked Eye. Biosensors. 2021; 11(9):317. https://doi.org/10.3390/bios11090317
Chicago/Turabian StyleKandasamy, Karthikeyan, Miftakhul Jannatin, and Yu-Chie Chen. 2021. "Rapid Detection of Pathogenic Bacteria by the Naked Eye" Biosensors 11, no. 9: 317. https://doi.org/10.3390/bios11090317
APA StyleKandasamy, K., Jannatin, M., & Chen, Y. -C. (2021). Rapid Detection of Pathogenic Bacteria by the Naked Eye. Biosensors, 11(9), 317. https://doi.org/10.3390/bios11090317