Profiles of Sterigmatocystin and Its Metabolites during Traditional Chinese Rice Wine Processing

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Pre-Treatment and Rice Wine Preparation
2.2. Instrument Conditions
2.2.1. LC–MS/MS Method for STG Analysis
2.2.2. LC–HRMS/MS Method for the Identification of STG Degradation Products and Non-Targeted Metabonomics
2.3. Statistical Analysis
3. Results and Discussion
3.1. Method Validation
3.2. The Fate of STG within the Chinese Rice Wine Process
3.2.1. Washing and Soaking
3.2.2. Influence of Steaming Time on STG Levels
3.2.3. Influence of Rice Leaven Levels on the Concentration of STG
3.2.4. Influence of Fermentation Time on STG Levels
3.3. Correlation of STG Levels between Soaked Rice and Rice Wine Final Product
3.4. Identification of Fermentation Degradation Products of STG
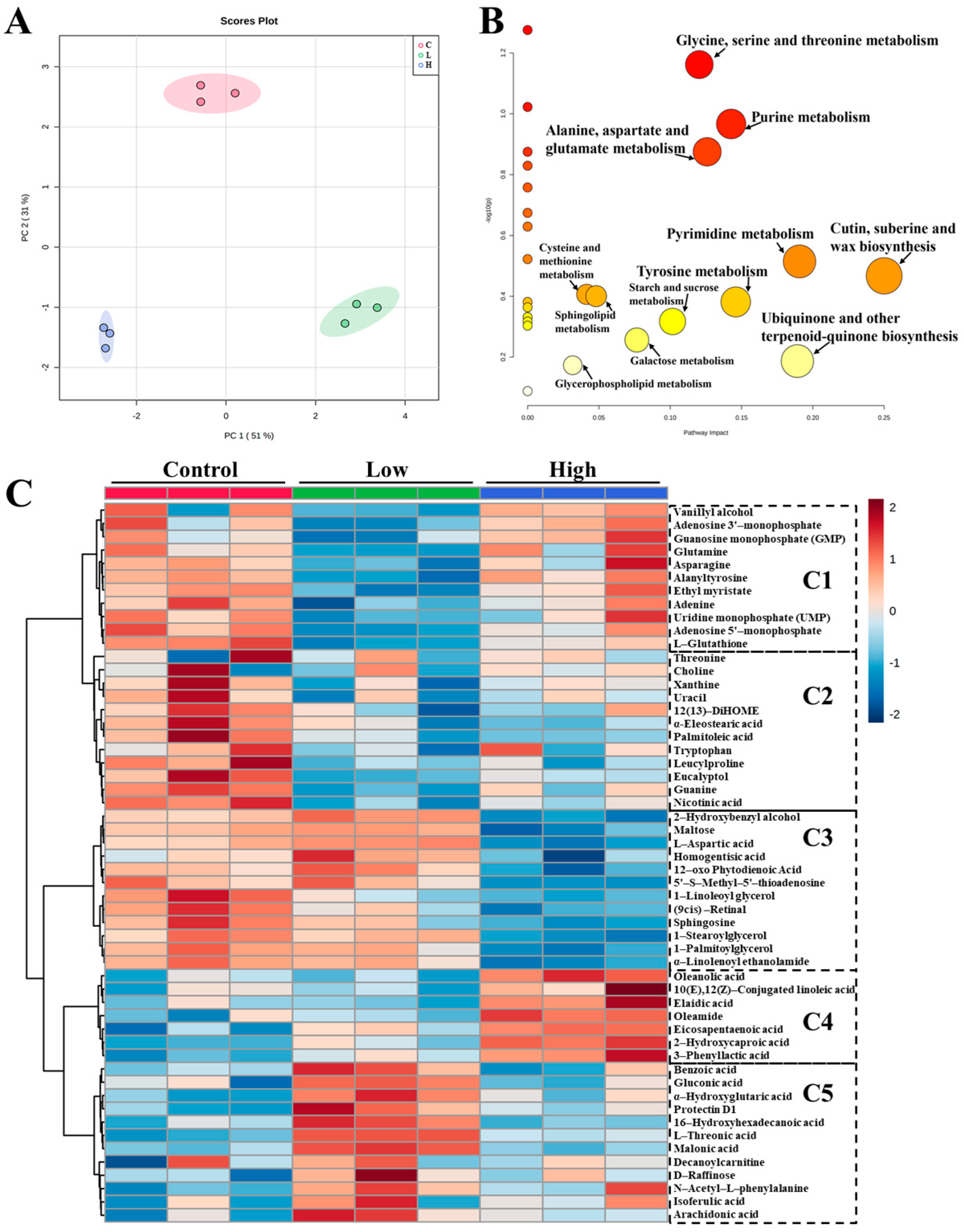
3.5. Effects of STG on the Metabolite Profiles of Rice Wine
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Gao, Q.; Ma, J.J.; Liu, Q.; Liao, M.; Xiao, J.J.; Jiang, M.H. Effect of application method and formulation on prothioconazole residue behavior and mycotoxin contamination in wheat. Sci. Total Environ. 2020, 729, 139019. [Google Scholar] [CrossRef] [PubMed]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Brauman, K.A.; Cunniffe, N.J.; Fedoroff, N.V.; Finegold, C.; Garrett, K.A.; Gilligan, C.A.; Jones, C.M.; et al. The persistent threat of emerging plant disease pandemics to global food security. Proc. Natl. Acad. Sci. USA 2021, 118, e2022239118. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Ryu, D. Worldwide occurrence of mycotoxins in cereals and cereal derived food products: Public health perspectives of their co-occurrence. J. Agric. Food Chem. 2017, 65, 7034–7051. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Song, G.; Lim, W. Effects of mycotoxin-contaminated feed on farm animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef] [PubMed]
- Jallow, A.; Xie, H.; Tang, X.; Qi, Z.; Li, P. Worldwide aflatoxin contamination of agricultural products and foods: From occurrence to control. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2332–2381. [Google Scholar] [CrossRef]
- Tian, Y.; Yu, D.Z.; Liu, N.; Tang, Y.; Wu, A. Confrontation assays and mycotoxin treatment reveal antagonistic activities of trichoderma and the fate of fusarium mycotoxins in microbial interaction. Environ. Pollut. 2020, 267, 115559. [Google Scholar] [CrossRef]
- Kong, D.Z.; Xie, Z.J.; Liu, L.Q.; Song, S.S.; Kuang, H.; Cui, G.; Xu, C.L. Development of indirect competitive ELISA and lateral-flow immunochromatographic assay strip for the detection of sterigmatocystin in cereal products. Food Agric. Immunol. 2017, 28, 260–273. [Google Scholar] [CrossRef] [Green Version]
- (EFSA) European Food Safety Authority. Scientific Opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2013, 11, 3254. [Google Scholar] [CrossRef]
- Mol, J.; Pietri, A.; Macdonald, S.J.; Anagnostopoulos, C.; Spanjer, M. Survey on sterigmatocystin in food. EFSA Support. Publ. 2015, 12, 774E. [Google Scholar]
- Noel, L.; Chafey, C.; Testu, C. Sterigmatocystin presence in typical Latvian grains. Food Chem. 2008, 109, 243–248. [Google Scholar]
- Zhao, Y.R.; Liu, X.X.; Zhao, J.; Wang, F.H. Research progress of sterigmatocystin pollution in food. Chin. J. Food Hyg. 2016, 28, 680–682. [Google Scholar]
- OECD/FAO. OECD-FAO Agricultural Outlook 2020–2029. 2020. Available online: https://www.oecd-ilibrary.org/agriculture-and-food/oecd-fao-agricultural-outlook-2020-2029_1112c23b-en (accessed on 16 July 2020).
- Jin, Z.; Cai, G.; Wu, C.; Hu, Z.; Lu, J. Profiling the key metabolites produced during the modern brewing process of Chinese rice wine. Food Res. Int. 2021, 139, 109955. [Google Scholar] [CrossRef] [PubMed]
- Dzuman, Z.; Stranska-Zachariasova, M.; Vaclavikova, M.; Tomaniova, M.; Veprikova, Z.; Slavikova, P.; Hajslova, J. Fate of free and conjugated mycotoxins within the production of distiller’s dried grains with solubles (DDGS). J. Agric. Food Chem. 2016, 64, 5085–5092. [Google Scholar] [CrossRef]
- Wu, L.; Wang, B. Transformation of deoxynivalenol and its acetylated derivatives in Chinese steamed bread making, as affected by pH, yeast, and steaming time. Food Chem. 2016, 202, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, E.; Sadeghi, E.; Bohlouli, S.; Karami, F. Fates of deoxynivalenol and deoxynivalenol-3-glucoside from wheat flour to iranian traditional breads. Food Control 2018, 91, 339–343. [Google Scholar] [CrossRef]
- Metwally, M.M.; El-Sayed, A.M.; Abd, A.M.E.; Mehriz, A.; Abu, S.Y.H. Sterigmatocystin: Incidence, fate and production by A. versicolor in Ras cheese. Mycotoxin Res. 1997, 13, 61–66. [Google Scholar] [CrossRef]
- Veršilovskis, A.; Bartkevičs, V. Stability of sterigmatocystin during the bread making process and its occurrence in bread from the Latvian market. Mycotoxin Res. 2012, 28, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Mengelers, M.; Yang, S.P.; Saeger, S.D.; Boevre, M.D. Mycotoxin biomarkers of exposure: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Huang, J.; Ma, L.; Wang, F. Development and validation of a simple and fast method for simultaneous determination of aflatoxin B1 and sterigmatocystin in grains. Food Chem. 2017, 221, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.-M.; Zhang, R.; Jin, N.; Quan, R.; Chen, D.-Y.; Francis, F.; Wang, F.Z.; Kong, Z.Q.; Fan, B. Effect of processing on herbicide residues and metabolite formation during traditional Chinese tofu production. LWT Food Sci. Technol. 2020, 131, 109707. [Google Scholar] [CrossRef]
- Xu, L.W.; Xu, X.X.; Guo, L.L.; Wang, Z.X.; Wu, X.L.; Kuang, H.; Xu, C.L. Potential Environmental Health Risk Analysis of Neonicotinoids and a Synergist. Environ. Sci. Technol. 2021, 55, 7541–7550. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.W.; Guo, L.L.; Wang, Z.X.; Xu, X.X.; Zhang, S.; Wu, X.L.; Kuang, H.; Xu, C.L. Profiling and identification of biocatalyzed transformation of sulfoxaflor in vivo. Angew. Chem. Int. Ed. 2020, 59, 16218–16224. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Fang, S.; Zhao, M.; Liu, W.; Jin, H. Effects of bisphenol a and bisphenol s exposure at low doses on the metabolome of adolescent male sprague-dawley rats. Chem. Res. Toxicol. 2021, 34, 1578–1587. [Google Scholar] [CrossRef]
- EC. Commission Regulation No. 657/2002 of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Union 2002, L221, 8–36. [Google Scholar]
- EC. Commission Regulation No. 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, L70, 12–34. [Google Scholar]
- Zhong, L.; Carere, J.; Mats, L.; Lu, Z.X.; Lu, F.X.; Zhou, T. Formation of glutathione patulin conjugates associated with yeast fermentation contributes to patulin reduction-sciencedirect. Food Control 2021, 123, 107334. [Google Scholar] [CrossRef]
- Sampaolesi, S.; Gamba, R.R.; Antoni, G.; Peláez, Á. Potentiality of yeasts obtained as beer fermentation residue to be used as probiotics. LWT Food Sci. Technol. 2019, 113, 108251. [Google Scholar] [CrossRef]
- Wall-Martínez, H.A.; Pascari, X.; Ramos, A.J.; Marín, S.; Sanchis, V. Fate of the mycotoxins in the wort and yeast during ale and lager fermentation and their evaluation under different technological parameters. LWT Food Sci. Technol. 2020, 132, 109877. [Google Scholar] [CrossRef]
- Huys, G.; Daniel, H.M.; Vuyst, L.D. Taxonomy and Biodiversity of Sourdough Yeasts and Lactic Acid Bacteria. 2013. Available online: https://link.springer.com/chapter/10.1007/978-1-4614-5425-0_5 (accessed on 1 January 2013).
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.G.; Bertin, G.; Jeminet, G.; Jouany, J.P. Adsorption of Zearalenone by beta-D-glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef]
- Kosztik, J.; Mörtl, M.; Székács, A.; Kukolya, J.; Bata-Vidács, I. Aflatoxin b1 and sterigmatocystin binding potential of lactobacilli. Toxins 2020, 12, 756. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowic, A. Binding of zearalenone, aflatoxin B1, and ochratoxin a by yeast-based products: A method for quantification of adsorption performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, R.J.; Kirksey, J.W.; Blankenship, B.R. Conversion of aflatoxin B1 to isomeric hydroxy compounds by Rhizopus subspecies. J. Agric. Food Chem. 1972, 20, 1100–1102. [Google Scholar] [CrossRef]
- Cole, R.J.; Kirksey, J.W. Aflatoxin G1 metabolism by Rhizopus species. J. Agric. Food Chem. 1971, 19, 222–223. [Google Scholar] [CrossRef] [PubMed]
- García-Béjar, B.; Owens, R.A.; Briones, A.; Arévalo-Villena, M. Proteomic profiling and glycomic analysis of the yeast cell wall in strains with Aflatoxin B1 elimination ability. Environ. Microbiol. 2021, 23, 5305–5319. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Gao, K.; Li, X.N.; Tang, Z.; Xiang, L.; Zhao, H.Z.; Fu, J.J.; Ling Wang, L.; Zhu, N.L.; Cai, Z.W.; et al. Mass Spectrometry-Based Metabolomics Reveals Occupational Exposure to Per- and Polyfluoroalkyl Substances Relates to Oxidative Stress, Fatty Acid beta-Oxidation Disorder, and Kidney Injury in a Manufactory in China. Environ. Sci. Technol. 2019, 53, 9800–9809. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Lee, S.; Singh, D.; Jang, E.S.; Shin, H.W.; Moon, B.S.; Lee, C.H. Time-resolved comparative metabolomes for koji fermentation with brown-, white-, and giant embryo-rice. Food Chem. 2017, 231, 258–266. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Matrix | Linear Range | Regression Equation | R2 | ME/% | LOD (μg/kg) | LOQ (μg/kg) |
|---|---|---|---|---|---|---|---|
| STG | Solvent | 5–200 | y = 190,442x + 268,234 | 0.9953 | |||
| Soaked rice | 5–200 | y = 176,167x + 236,281 | 0.9979 | −7.0 | 0.01 | 0.03 | |
| Steamed rice | 5–200 | y = 179,674x + 238,395 | 0.9969 | −6.0 | 0.01 | 0.03 | |
| Fermented rice | 5–200 | y = 208,146x + 392,325 | 0.9959 | +9.0 | 0.01 | 0.03 | |
| Fermented wine | 5–200 | y = 256,280x + 565,254 | 0.9910 | +35.0 | 0.07 | 0.25 | |
| Sample | 20 μg/kg | 100 μg/kg | 200 μg/kg | ||||
| Recoveries (%) | RSD (%) | Recoveries (%) | RSD (%) | Recoveries (%) | RSD (%) | ||
| Soaked rice | 102 | 2.3 | 107 | 2.3 | 119 | 2.4 | |
| Steamed rice | 77 | 6.2 | 85 | 8.7 | 112 | 3.5 | |
| Fermented rice | 118 | 2.1 | 118 | 7.6 | 116 | 6.9 | |
| Fermented wine | 73 | 3.6 | 105 | 5.0 | 119 | 4.3 | |
| Sample | Rice | Washed Rice | Soaked Rice | Steam Rice | Fermented Rice-1 g | Fermented Rice-3 g | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 15 min | 25 min | 35 min | 12 h | 36 h | 84 h | 12 h | 36 h | 84 h | ||||
| Level/(μg/kg) | 986.1 | 822.3 * | 750.8 * | 738.8 a | 733.2 a | 737.7 a | 594.8 *,a | 711.6 *,b | 913.1 *,c | 592.8 *,a | 712.3 *,b | 858.1 *,c |
| SD | 16.4 | 39.8 | 14.4 | 16.2 | 46.4 | 37.1 | 60.8 | 32.3 | 23.6 | 36.8 | 78.4 | 17.7 |
| Sample | Fermented rice-9g | Separated fermented wine | Separated fermented rice | Total rice wine | ||||||||
| 12 h | 36 h | 84 h | 1 g | 3 g | 9 g | 1 g | 3 g | 9 g | 1 g | 3 g | 9 g | |
| Level/(μg/kg) | 586.6 *,a | 736.7 *,b | 818.5 *,c | 169 a | 126.2 b | 112.2 c | 1214.1 a | 1164.9 a | 956.0 b | 925.4 | 850.6 | 613.4 * |
| SD | 74.9 | 19.4 | 58.3 | 2 | 5.5 | 3.9 | 43.1 | 108.4 | 39.0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Xu, L.; Xu, X.; Wu, X.; Kuang, H.; Xu, C. Profiles of Sterigmatocystin and Its Metabolites during Traditional Chinese Rice Wine Processing. Biosensors 2022, 12, 212. https://doi.org/10.3390/bios12040212
Zhang J, Xu L, Xu X, Wu X, Kuang H, Xu C. Profiles of Sterigmatocystin and Its Metabolites during Traditional Chinese Rice Wine Processing. Biosensors. 2022; 12(4):212. https://doi.org/10.3390/bios12040212
Chicago/Turabian StyleZhang, Jia, Liwei Xu, Xinxin Xu, Xiaoling Wu, Hua Kuang, and Chuanlai Xu. 2022. "Profiles of Sterigmatocystin and Its Metabolites during Traditional Chinese Rice Wine Processing" Biosensors 12, no. 4: 212. https://doi.org/10.3390/bios12040212
APA StyleZhang, J., Xu, L., Xu, X., Wu, X., Kuang, H., & Xu, C. (2022). Profiles of Sterigmatocystin and Its Metabolites during Traditional Chinese Rice Wine Processing. Biosensors, 12(4), 212. https://doi.org/10.3390/bios12040212