ECIS Based Electric Fence Method for Measurement of Human Keratinocyte Migration on Different Substrates

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Instrumentation
2.3. ECIS Methods
2.3.1. Multiple Frequency Time-Series (MFT) Measurements
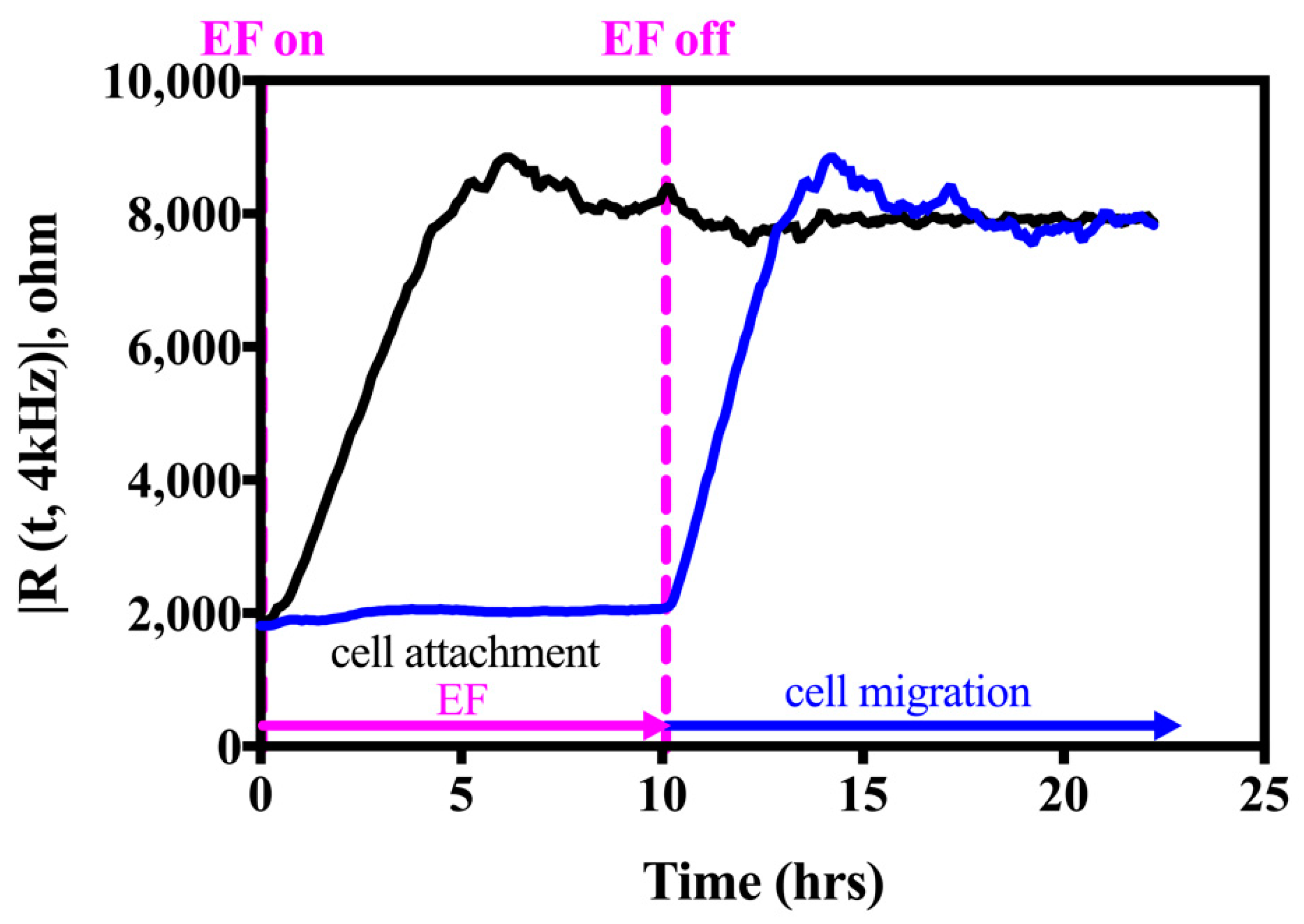
2.3.2. Assess Cell Migration Ability with an Electric Fence (EF) Assay
2.3.3. Frequency Scan Measurement
2.3.4. Micromotion Measurement
2.4. Zeta Potential Analysis
2.5. Analysis of Initial Adhesion
2.6. Statistical Analysis
3. Results
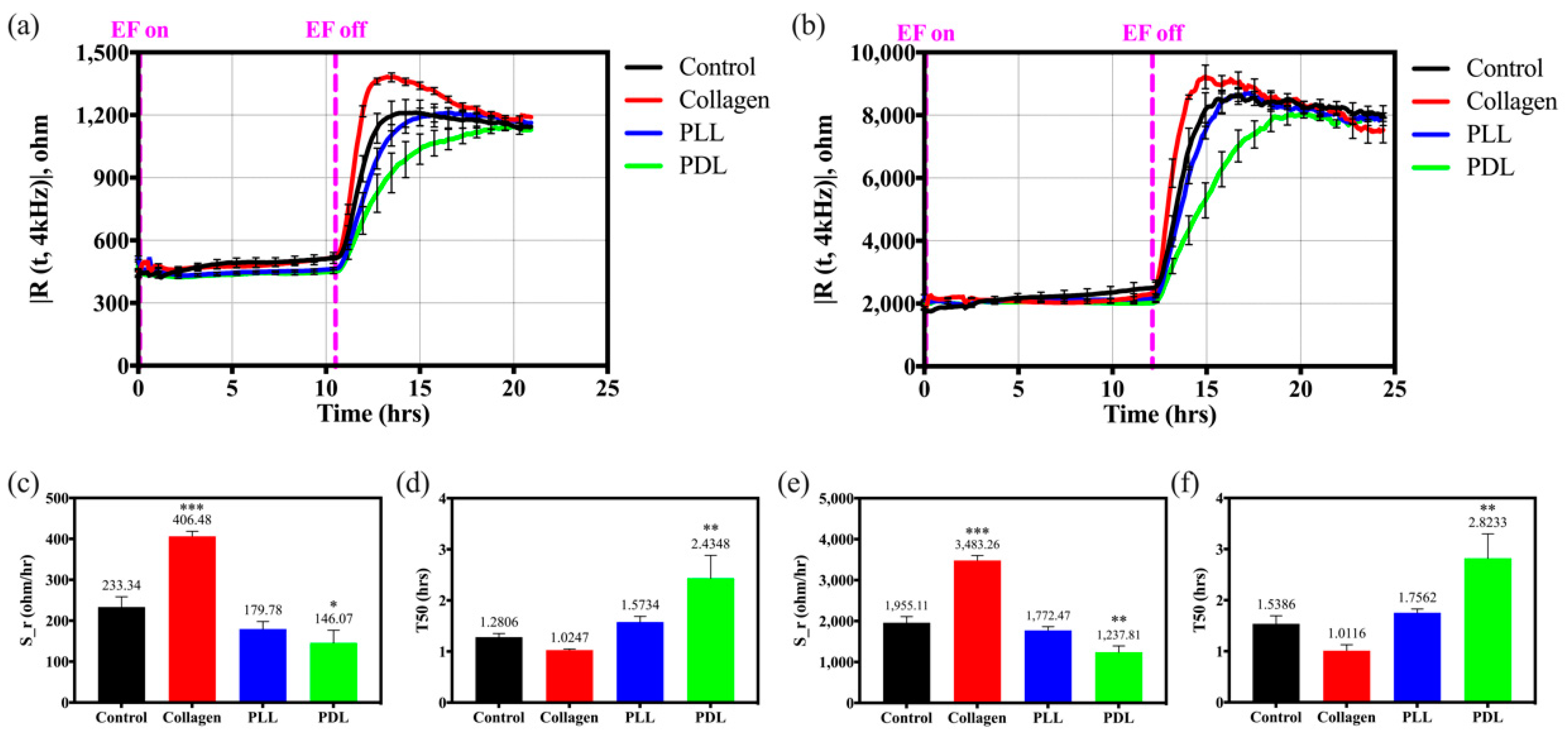
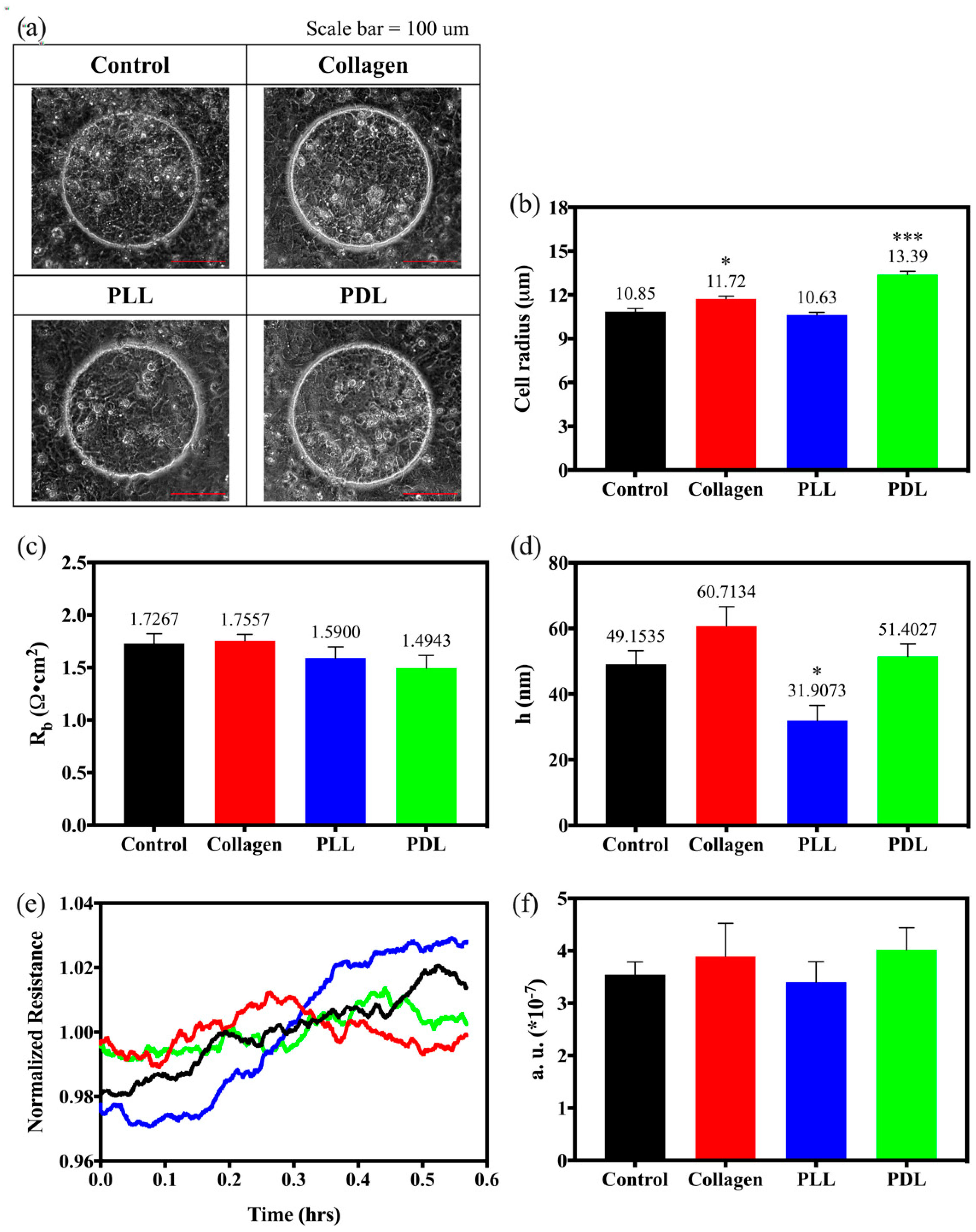
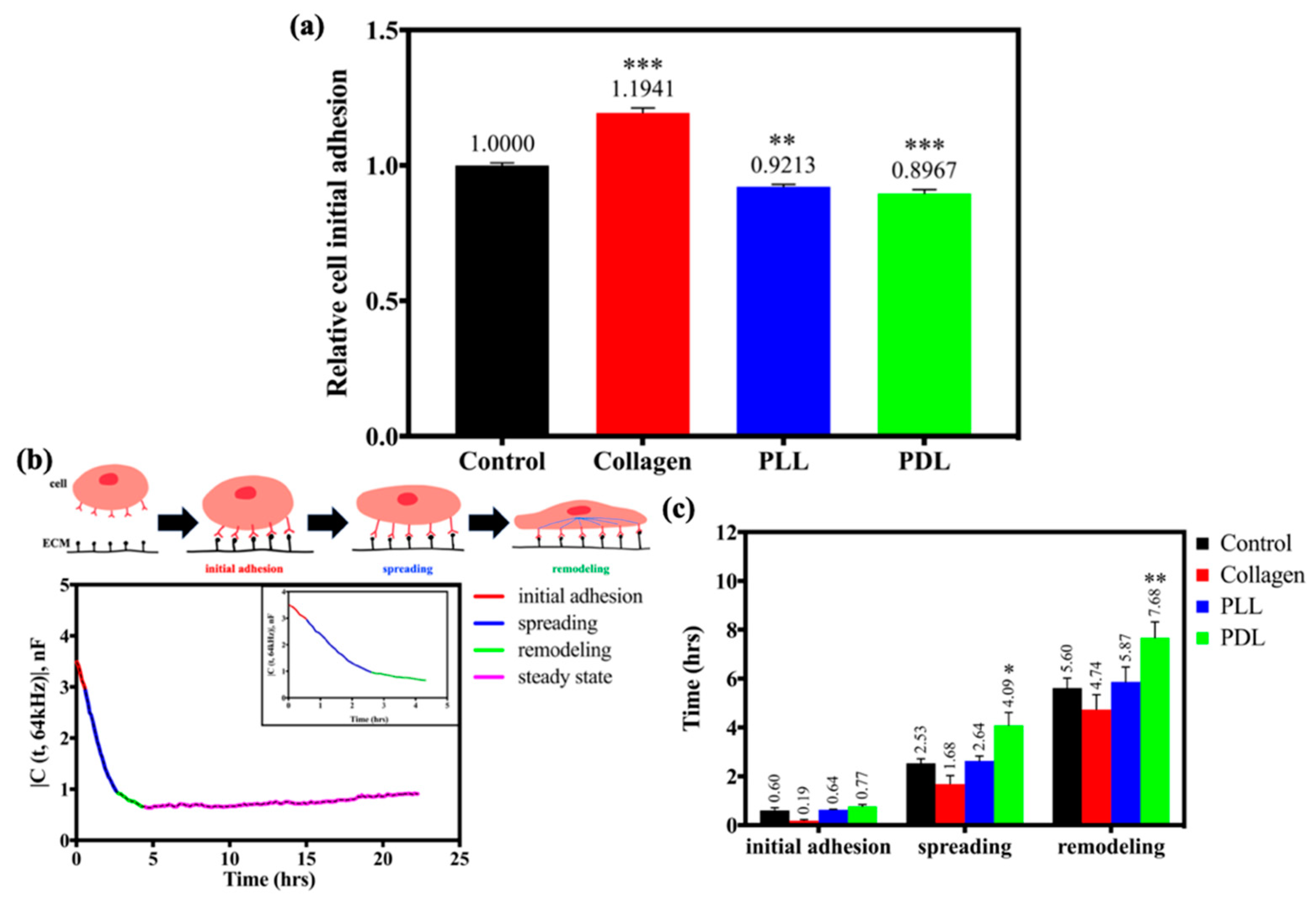
3.1. Effects of Different Substrates on HaCaT Cell Attachment and Spreading
3.2. Effects of Different Substrates on HaCaT Cell Migration
3.3. Effects of Morphological Parameters on the HaCaT Cultured on Different Substrates
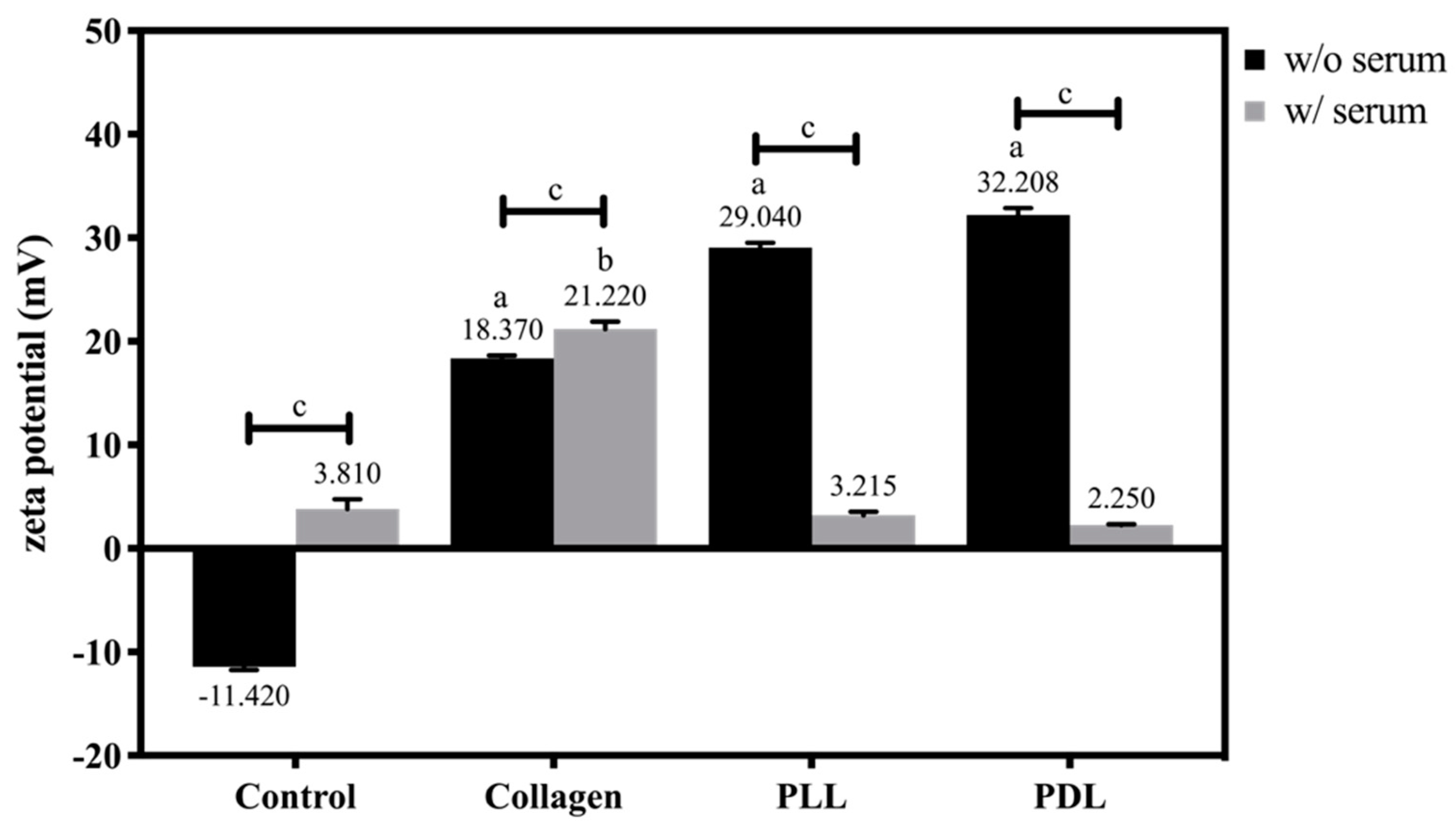
3.4. Effects of Electrostatic Cell-Substrate Interactions on the Initial Cell Adhesion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F. The activated keratinocyte: Up regulation of cell adhesion and migration during wound healing. J. Trauma Acute Care Surg. 1990, 30, 144–148. [Google Scholar] [CrossRef]
- Martin, P. Wound healing--aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Kirfel, G.; Herzog, V. Migration of epidermal keratinocytes: Mechanisms, regulation, and biological significance. Protoplasma 2004, 223, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Raja; Sivamani, K.; Garcia, M.S.; Isseroff, R.R. Wound re-epithelialization: Modulating keratinocyte migration in wound healing. Front. Biosci. 2007, 12, 2849–2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschlager, M.; Dolznig, H. In vitro cell migration and invasion assays. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef]
- Keese, C.R.; Wegener, J.; Walker, S.R.; Giaever, I. Electrical wound-healing assay for cells in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 1554–1559. [Google Scholar] [CrossRef] [Green Version]
- Stolwijk, J.A.; Matrougui, K.; Renken, C.W.; Trebak, M. Impedance analysis of GPCR-mediated changes in endothelial barrier function: Overview and fundamental considerations for stable and reproducible measurements. Pflügers Arch.-Eur. J. Physiol. 2015, 467, 2193–2218. [Google Scholar] [CrossRef] [Green Version]
- Wegener, J.; Keese, C.R.; Giaever, I. Electric cell–substrate impedance sensing (ECIS) as a noninvasive means to monitor the kinetics of cell spreading to artificial surfaces. Exp. Cell Res. 2000, 259, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.M.; Keese, C.R.; Giaever, I. Impedance analysis of MDCK cells measured by electric cell-substrate impedance sensing. Biophys. J. 1995, 69, 2800–2807. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Tung, T.H.; Huang, C.C.; Lin, S.Y.; Chao, S.C.; Chiu, S.P.; Lee, S.P.; Lo, C.M. Electrochemical Assessment of Anticancer Compounds on the Human Tongue Squamous Carcinoma Cells. Sensors 2020, 20, 2632. [Google Scholar] [CrossRef]
- Lo, C.M.; Lo, J.C.; Sato, P.Y.; Yeung, T.L.; Mok, S.C.; Yip, K.P. Monitoring of ovarian cancer cell invasion in real time with frequency-dependent impedance measurement. Am. J. Physiol. Cell Physiol. 2016, 311, C1040–C1047. [Google Scholar] [CrossRef]
- Bagnaninchi, P.O.; Drummond, N. Real-time label-free monitoring of adipose-derived stem cell differentiation with electric cell-substrate impedance sensing. Proc. Natl. Acad. Sci. USA 2011, 108, 6462–6467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, A.Y.; Kho, D.T.; Johnson, R.H.; Graham, E.S.; O’Carroll, S.J. In Viedance Sensing (ECIS)-Zθ Technology. Biosensors 2018, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Bosanquet, D.C.; Harding, K.G.; Jiang, W.G. ECIS, cellular adhesion and migration in keratinocytes. In Electric Cell-Substrate Impedance Sensing and Cancer Metastasis; Jiang, W.G., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 217–237. [Google Scholar]
- Renken, C.; Keese, C.R.; Giaever, I. Automated assays for quantifying cell migration. BioTechniques 2010, 49, 844. [Google Scholar] [CrossRef] [Green Version]
- Cavallini, F.; Tarantola, M. ECIS based wounding and reorganization of cardiomyocytes and fibroblasts in co-cultures. Prog. Biophys. Mol. Biol. 2019, 144, 116–127. [Google Scholar] [CrossRef]
- Evora, A.; de Freitas, V.; Mateus, N.; Fernandes, I. The effect of anthocyanins from red wine and blackberry on the integrity of a keratinocyte model using ECIS. Food Funct. 2017, 8, 3989–3998. [Google Scholar] [CrossRef]
- Giaever, I.; Keese, C.R. Micromotion of mammalian cells measured electrically. Proc. Natl. Acad. Sci. USA 1991, 88, 7896–7900. [Google Scholar] [CrossRef] [Green Version]
- Engin, A.B.; Nikitovic, D.; Neagu, M.; Henrich-Noack, P.; Docea, A.O.; Shtilman, M.I.; Golokhvast, K.; Tsatsakis, A.M. Mechanistic understanding of nanoparticles’ interactions with extracellular matrix: The cell and immune system. Part. Fibre Toxicol. 2017, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef]
- Birbrair, A.; Zhang, T.; Files, D.C.; Mannava, S.; Smith, T.; Wang, Z.-M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner. Stem Cell Res. Ther. 2014, 5, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaRocca, P.; Barker, S. Tissue culture surface treatments. Cell/Line 1996, 6, 1–6. [Google Scholar]
- Sitterley, G. Poly-l-lysine cell attachment protocol. BioFiles 2008, 3, 12. [Google Scholar]
- Lu, H.; Guo, L.; Kawazoe, N.; Tateishi, T.; Chen, G. Effects of poly (L-lysine), poly (acrylic acid) and poly (ethylene glycol) on the adhesion, proliferation and chondrogenic differentiation of human mesenchymal stem cells. J. Biomater. Sci. Polym. Ed. 2009, 20, 577–589. [Google Scholar] [CrossRef]
- Lauffenburger, D.A.; Horwitz, A.F. Cell migration: A physically integrated molecular process. Cell 1996, 84, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Palecek, S.P.; Cox, E.A.; Huttenlocher, A.; Lauffenburger, D.A.; Horwitz, A.F. Integrin adhesion in cell migration. In Advances in Molecular and Cell Biology; Bittar, E.E., Garrod, D.R., North, A.J., Chidgey, M.A.J., Eds.; JAI press: Stamford, CT, USA, 1999; Volume 28, pp. 367–388. [Google Scholar]
- O’toole, E. Extracellular matrix and keratinocyte migration. Clin. Exp. Dermatol. 2001, 26, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Toda, K.-L.; Grinnell, F. Activation of human keratinocyte migration on type I collagen and fibronectin. J. Cell Sci. 1990, 96, 197–205. [Google Scholar] [CrossRef]
- Sanganyado, E.; Lu, Z.; Fu, Q.; Schlenk, D.; Gan, J. Chiral pharmaceuticals: A review on their environmental occurrence and fate processes. Water Res. 2017, 124, 527–542. [Google Scholar] [CrossRef]
- Nelson, D.L.; Lehninger, A.L.; Cox, M.M. Lehninger Principles of Biochemistry; Macmillan: New York, NY, USA, 2008; p. 474. [Google Scholar]
- Zhao, B.-W.; Zhou, Z.; Shen, Y. Effects of chirality on gene delivery efficiency of polylysine. Chin. J. Polym. Sci. 2016, 34, 94–103. [Google Scholar] [CrossRef]
- Kim, Y.H.; Baek, N.S.; Han, Y.H.; Chung, M.-A.; Jung, S.-D. Enhancement of neuronal cell adhesion by covalent binding of poly-D-lysine. J. Neurosci. Methods 2011, 202, 38–44. [Google Scholar] [CrossRef]
- Qian, L.; Saltzman, W.M. Improving the expansion and neuronal differentiation of mesenchymal stem cells through culture surface modification. Biomaterials 2004, 25, 1331–1337. [Google Scholar] [CrossRef]
- Mazia, D.; Schatten, G.; Sale, W. Adhesion of cells to surfaces coated with polylysine. Applications to electron microscopy. J. Cell Biol. 1975, 66, 198–200. [Google Scholar] [CrossRef] [Green Version]
- Rigot, V.; Lehmann, M.; André, F.; Daemi, N.; Marvaldi, J.; Luis, J. Integrin ligation and PKC activation are required for migration of colon carcinoma cells. J. Cell Sci. 1998, 111, 3119–3127. [Google Scholar] [CrossRef]
- Novák, P.; Havlíček, V. Protein extraction and precipitation. In Proteomic Profiling and Analytical Chemistry; Silberring, J., Ciborowski, P., Eds.; Elsevier: Oxford, UK, 2016; pp. 51–62. [Google Scholar]
- Škvarla, J. A physico-chemical model of microbial adhesion. J. Chem. Soc. Faraday Trans. 1993, 89, 2913–2921. [Google Scholar] [CrossRef]
- Tung, T.-H.; Wang, S.-H.; Huang, C.-C.; Su, T.-Y.; Lo, C.-M. Use of Discrete Wavelet Transform to Assess Impedance Fluctuations Obtained from Cellular Micromotion. Sensors 2020, 20, 3250. [Google Scholar] [CrossRef]
- Opp, D.; Wafula, B.; Lim, J.; Huang, E.; Lo, J.-C.; Lo, C.-M. Use of electric cell–substrate impedance sensing to assess in vitro cytotoxicity. Biosens. Bioelectron. 2009, 24, 2625–2629. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, Y.-H.; Chiu, W.-C.; Fuh, S.-R.; Lai, Y.-T.; Tung, T.-H.; Huang, C.-C.; Lo, C.-M. ECIS Based Electric Fence Method for Measurement of Human Keratinocyte Migration on Different Substrates. Biosensors 2022, 12, 293. https://doi.org/10.3390/bios12050293
Hung Y-H, Chiu W-C, Fuh S-R, Lai Y-T, Tung T-H, Huang C-C, Lo C-M. ECIS Based Electric Fence Method for Measurement of Human Keratinocyte Migration on Different Substrates. Biosensors. 2022; 12(5):293. https://doi.org/10.3390/bios12050293
Chicago/Turabian StyleHung, Yu-Han, Wei-Chih Chiu, Shyh-Rong Fuh, Yi-Ting Lai, Tse-Hua Tung, Chun-Chung Huang, and Chun-Min Lo. 2022. "ECIS Based Electric Fence Method for Measurement of Human Keratinocyte Migration on Different Substrates" Biosensors 12, no. 5: 293. https://doi.org/10.3390/bios12050293
APA StyleHung, Y. -H., Chiu, W. -C., Fuh, S. -R., Lai, Y. -T., Tung, T. -H., Huang, C. -C., & Lo, C. -M. (2022). ECIS Based Electric Fence Method for Measurement of Human Keratinocyte Migration on Different Substrates. Biosensors, 12(5), 293. https://doi.org/10.3390/bios12050293