
Anti-Candidal Activity of the Parasitic Plant Orobanche crenata Forssk

 ,
, 
 ,
,  ,
,  , , , , , ,
, , , , , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Antifungal Activity
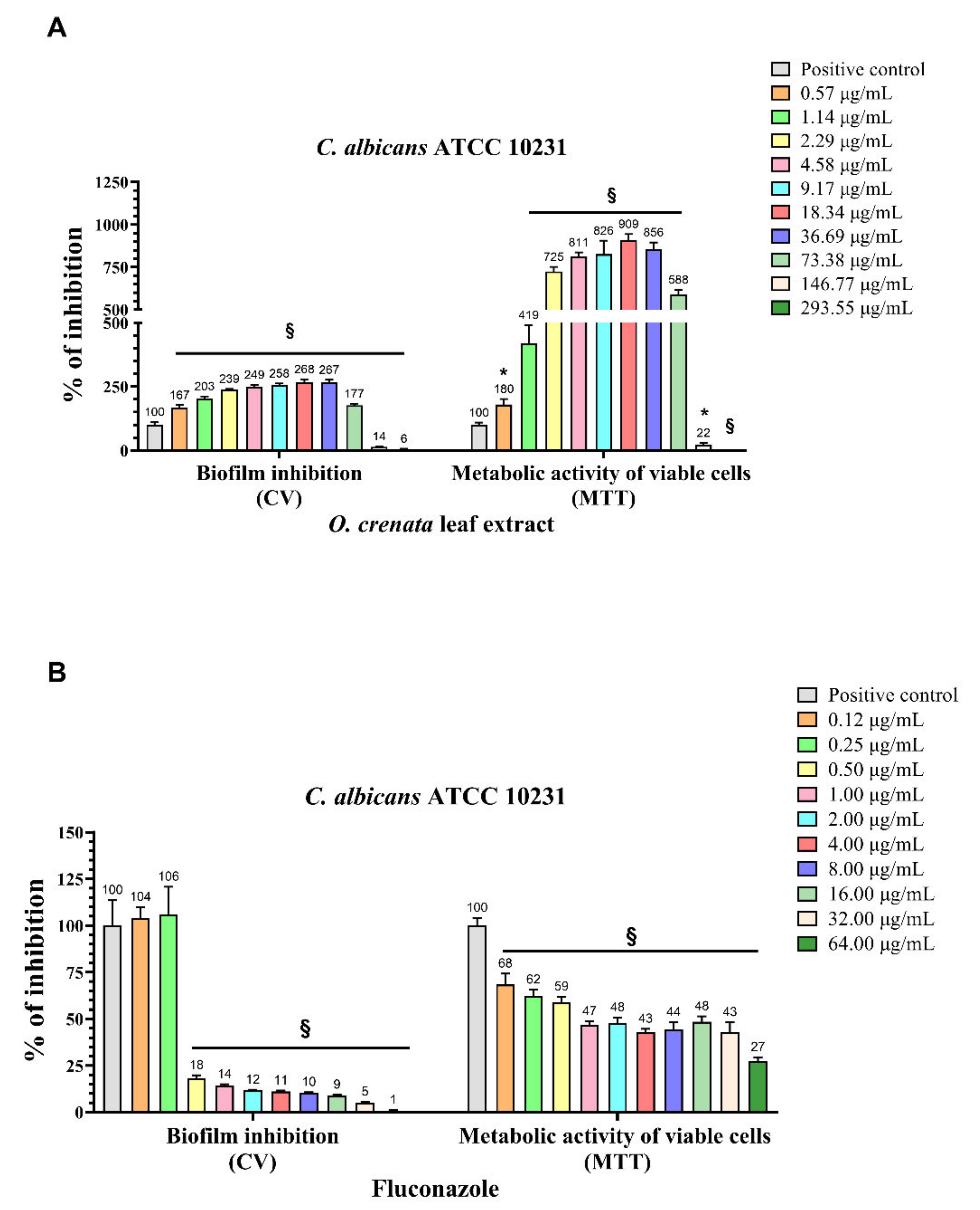
2.2. Biofilm Inhibition and 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Reduction Assays
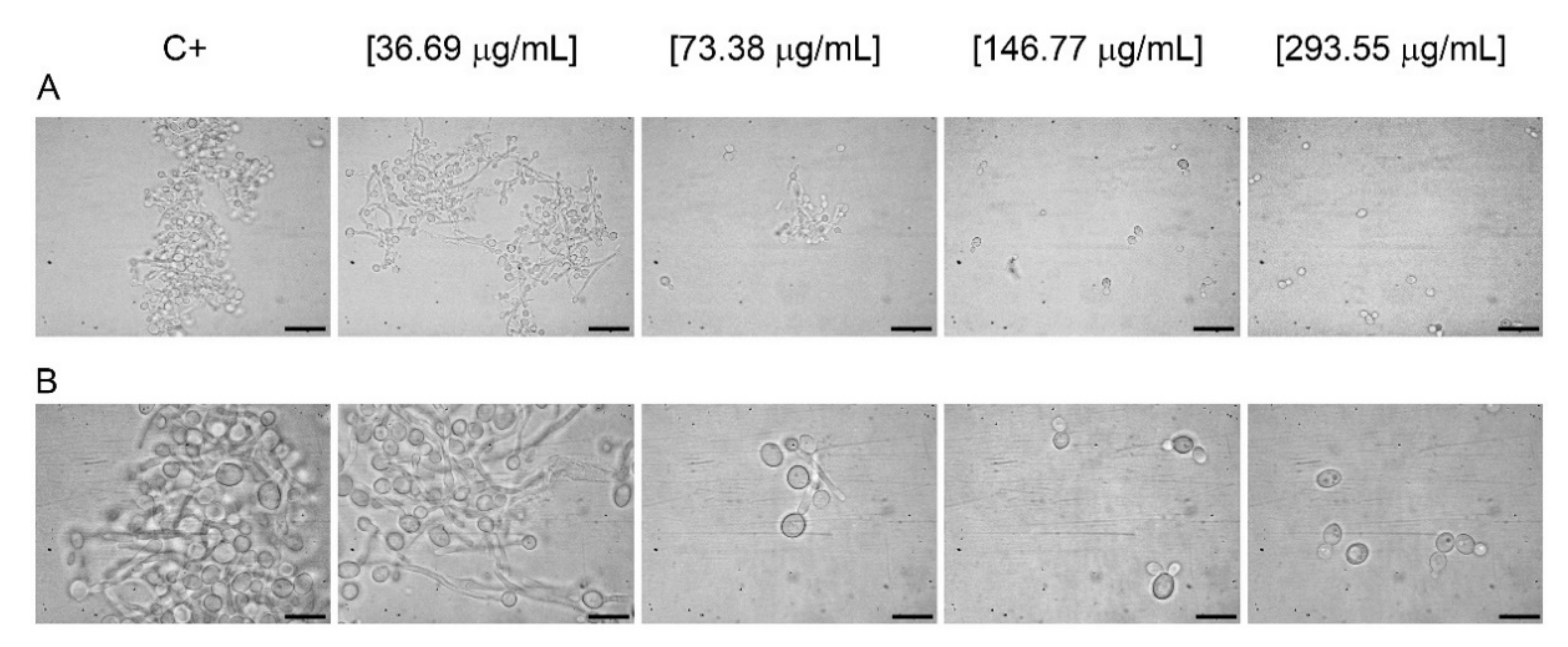
2.3. Germ Tube Assay
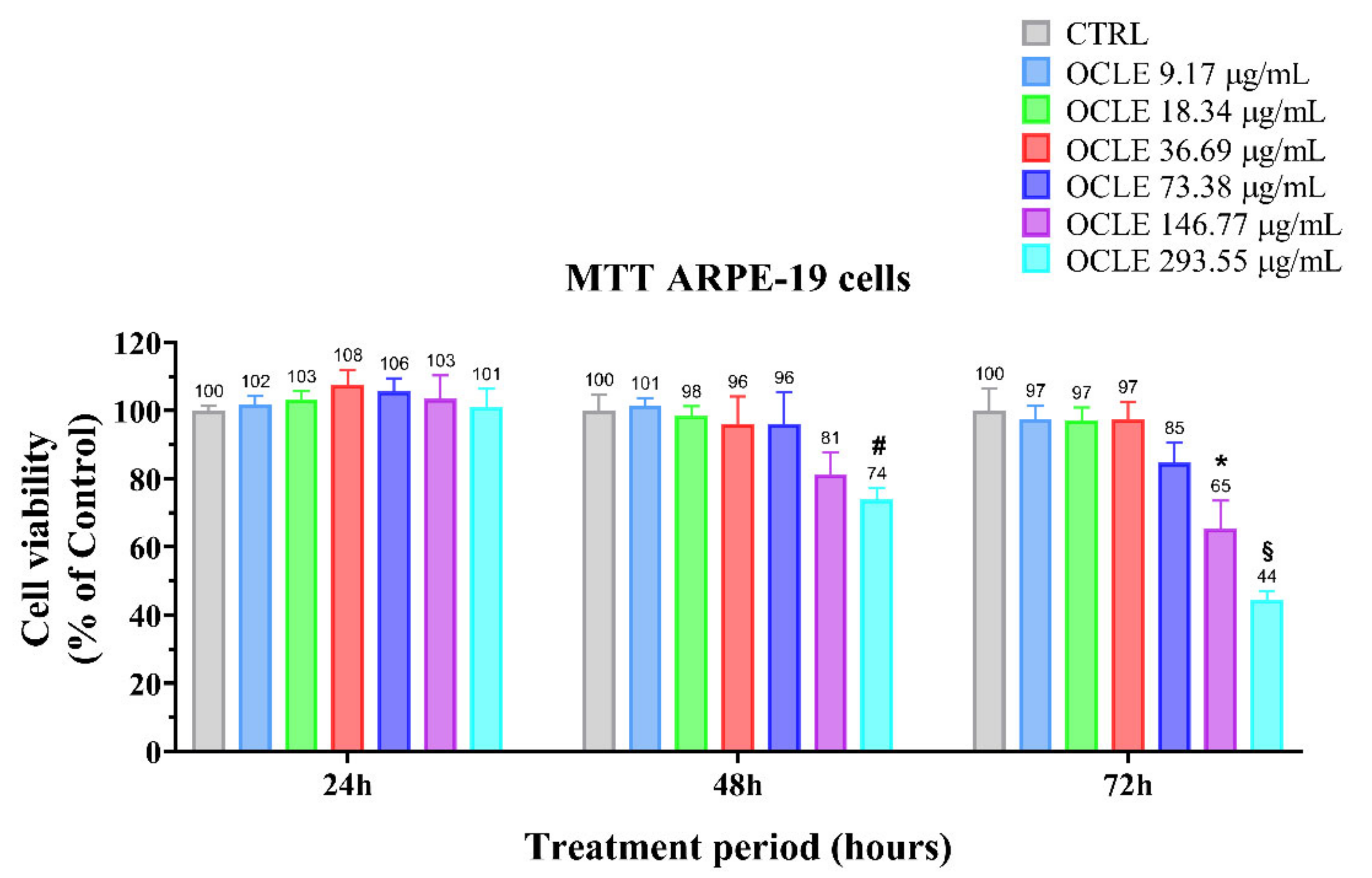
2.4. Cytotoxicity Assay
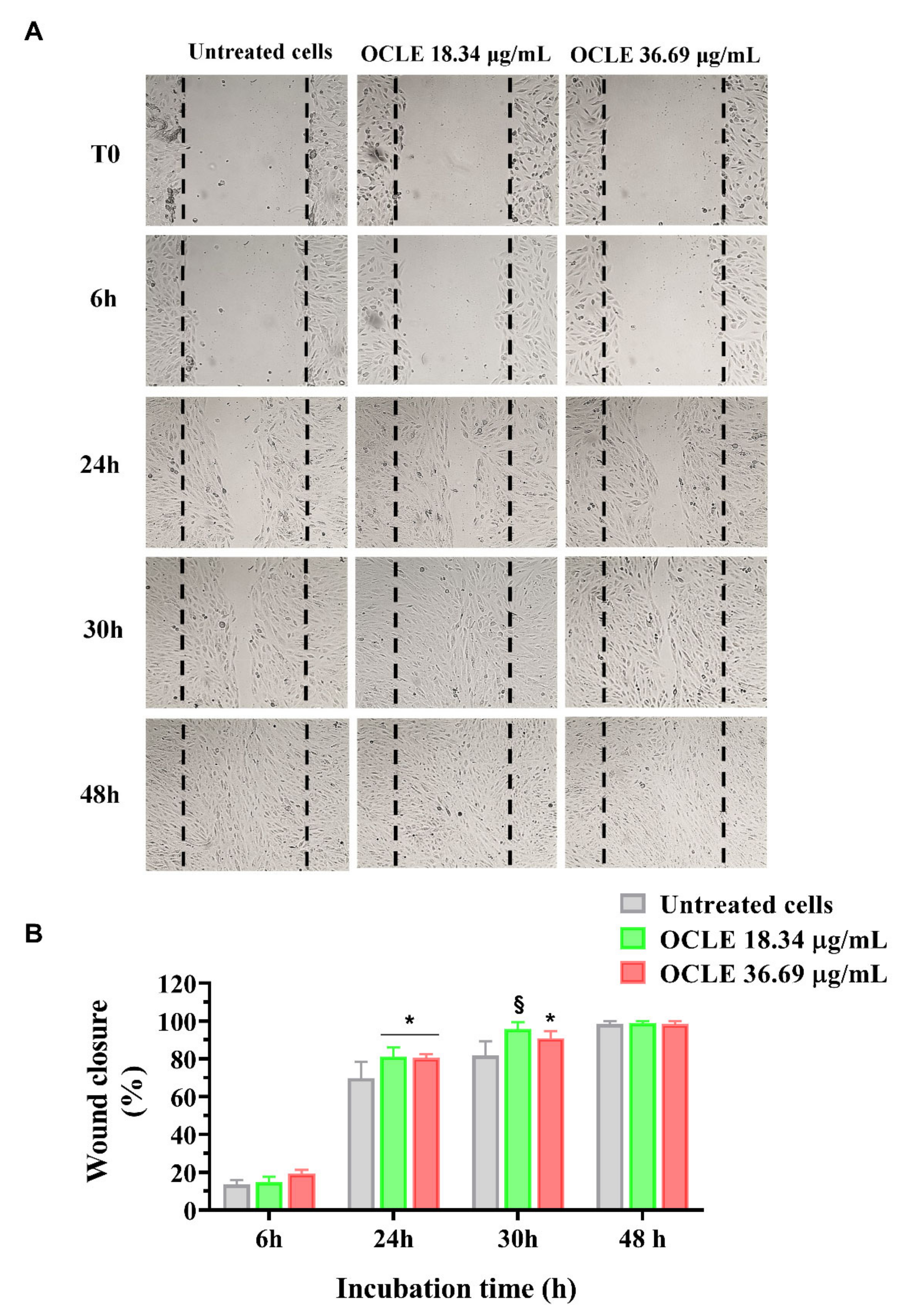
2.5. Wound Healing Assay
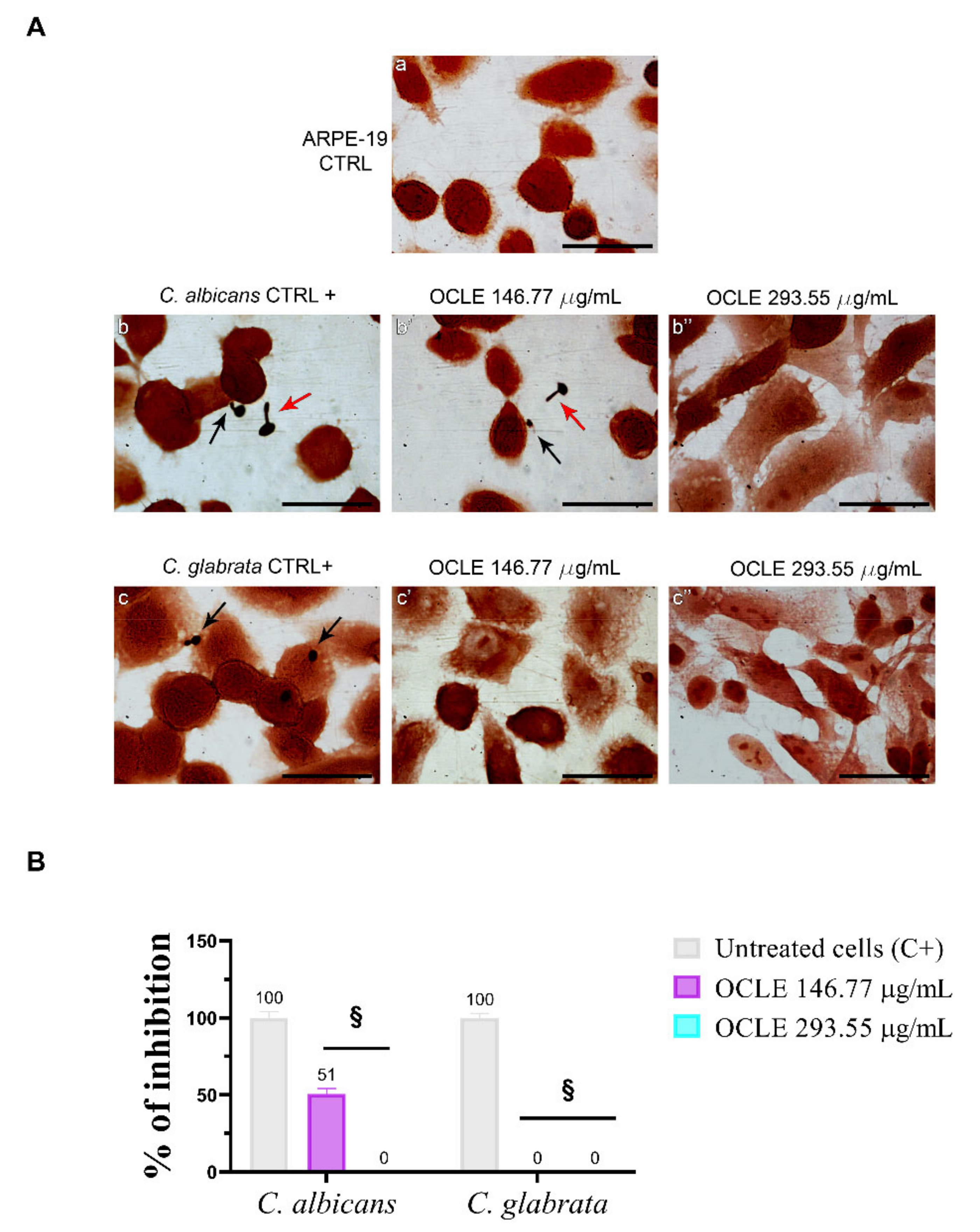
2.6. Effect on the Adhesion of Candida on the Human Retinal Pigment Epithelial Cell Line (ARPE-19)
2.7. Determination of Total Phenols Content of O. crenata Leaf Extract
2.8. Chemical Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Preparation of the Extract
4.3. Microorganisms
4.4. Antifungal Susceptibility Test
4.5. In Vitro Biofilm Formation and Inhibition Assay
4.6. Determination of Fungal Viability
4.7. Germ Tube Assay
4.8. Cell Culture
4.9. Cytotoxicity Assay
4.10. Wound Healing Assay
4.11. Adhesion Assay on Human Retinal Pigment Epithelial Cell Line (ARPE-19)
4.12. Total Phenolic Content
4.13. Chemical Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Integrated Taxonomic Information System. Candida Berkhout. Taxonomic Serial No.: 194591. NODC Taxonomic Code, Database (Version 8.0). Available online: https://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=194591#null (accessed on 20 October 2021).
- Singh, D.K.; Toth, R.; Gacser, A. Mechanisms of Pathogenic Candida Species to Evade the Host Complement Attack. Front. Cell. Infect. Microbiol. 2020, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottier, F.; Hall, R.A. Face/Off: The Interchangeable Side of Candida albicans. Front. Cell. Infect. Microbiol. 2019, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- Poulain, D. Candida albicans, plasticity and pathogenesis. Crit. Rev. Microbiol 2015, 41, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; Kavanagh, K.; Velasco-Torrijos, T. Targeting adhesion in fungal pathogen Candida albicans. Future Med. Chem. 2021, 13, 313–334. [Google Scholar] [CrossRef]
- Jacobsen, I.D.; Hube, B. Candida albicans morphology: Still in focus. Expert. Rev. Anti-Infect. Ther. 2017, 15, 327–330. [Google Scholar] [CrossRef] [Green Version]
- Erum, R.; Samad, F.; Khan, A.; Kazmi, S.U. A comparative study on production of extracellular hydrolytic enzymes of Candida species isolated from patients with surgical site infection and from healthy individuals and their co-relation with antifungal drug resistance. BMC Microbiol. 2020, 20, 368. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, S.; Li, H.; Shen, L.; Dong, C.; Sun, Y.; Chen, H.; Xu, B.; Zhuang, W.; Deighton, M.; et al. Biofilm Formation of Candida albicans Facilitates Fungal Infiltration and Persister Cell Formation in Vaginal Candidiasis. Front. Microbiol. 2020, 11, 1117. [Google Scholar] [CrossRef]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Jenks, J.D.; Cornely, O.A.; Chen, S.C.; Thompson, G.R., 3rd; Hoenigl, M. Breakthrough invasive fungal infections: Who is at risk? Mycoses 2020, 63, 1021–1032. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. Methods Mol. Biol. 2017, 1508, 17–65. [Google Scholar] [CrossRef]
- Sallam, A.; Lynn, W.; McCluskey, P.; Lightman, S. Endogenous Candida endophthalmitis. Expert. Rev. Anti-Infect. Ther. 2006, 4, 675–685. [Google Scholar] [CrossRef]
- Bae, J.H.; Lee, S.C. Intravitreal liposomal amphotericin B for treatment of endogenous Candida endophthalmitis. Jpn. J. Ophthalmol. 2015, 59, 346–352. [Google Scholar] [CrossRef]
- Shirvani, M.; Hosseini, S.; Maalhagh, M.; Mohaghegh, S. Endogenous Candida Endophthalmitis as a Rare Complication of Trans-Urethral Lithotripsy in a Healthy Woman: A Case Report. Turk. Oftalmol. Derg. 2019, 49, 294–296. [Google Scholar] [CrossRef]
- Kostick, D.A.; Foster, R.E.; Lowder, C.Y.; Meyers, S.M.; Mchenry, M.C. Endogenous Endophthalmitis Caused by Candida-Albicans in a Healthy Woman. Am. J. Ophthalmol. 1992, 113, 593–595. [Google Scholar] [CrossRef]
- Chen, S.J.; Chung, Y.M.; Liu, J.H. Endogenous Candida endophthalmitis after induced abortion. Am. J. Ophthalmol. 1998, 125, 873–875. [Google Scholar] [CrossRef]
- Sadeghi, G.; Ebrahimi-Rad, M.; Mousavi, S.F.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Emergence of non-Candida albicans species: Epidemiology, phylogeny and fluconazole susceptibility profile. J. Mycol. Med. 2018, 28, 51–58. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Silva, S.; Henriques, M. Candida glabrata: A review of its features and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Steinbach, W.J.; Dvorak, C.C. 293—Antifungal Agents. In Principles and Practice of Pediatric Infectious Diseases, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1484–1492.e5. [Google Scholar] [CrossRef]
- Galocha, M.; Pais, P.; Cavalheiro, M.; Pereira, D.; Viana, R.; Teixeira, M.C. Divergent Approaches to Virulence in C. albicans and C. glabrata: Two Sides of the Same Coin. Int. J. Mol. Sci. 2019, 20, 2345. [Google Scholar] [CrossRef] [Green Version]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Bilska-Stoklosa, J.; Hampelska, K.; Osmola, K.; Czajka, J.; Bogdanowicz-Gapinska, D.; Tomczak, H. Candida glabrata as an aetiological factor of the fulminant course of panophthalmitis. Ann. Agric. Environ. Med. 2020, 27, 540–543. [Google Scholar] [CrossRef]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Lee, C.H.; Chen, Y.C.; Chen, I.L.; Chen, F.J.; Chien, C.C. Impact of biofilm production by Candida species and antifungal therapy on mortality of patients with candidemia. Mycoses 2020, 63, 1382–1391. [Google Scholar] [CrossRef]
- Butler, M.S. The role of natural product chemistry in drug discovery. J. Nat. Prod. 2004, 67, 2141–2153. [Google Scholar] [CrossRef]
- Berlin Grace, V.M.; Viswanathan, S.; David Wilson, D.; Jagadish Kumar, S.; Sahana, K.; Maria Arbin, E.F.; Narayanan, J. Significant action of Tridax procumbens L. leaf extract on reducing the TNF-alpha and COX-2 gene expressions in induced inflammation site in Swiss albino mice. Inflammopharmacology 2020, 28, 929–938. [Google Scholar] [CrossRef]
- Moore, J.; Yousef, M.; Tsiani, E. Anticancer Effects of Rosemary (Rosmarinus officinalis L.) Extract and Rosemary Extract Polyphenols. Nutrients 2016, 8, 731. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula etnensis Raf. (Betulaceae) Extract Induced HO-1 Expression and Ferroptosis Cell Death in Human Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 2723. [Google Scholar] [CrossRef] [Green Version]
- Attaguile, G.; Russo, A.; Campisi, A.; Savoca, F.; Acquaviva, R.; Ragusa, N.; Vanella, A. Antioxidant activity and protective effect on DNA cleavage of extracts from Cistus incanus L. and Cistus monspeliensis L. Cell. Biol. Toxicol. 2000, 16, 83–90. [Google Scholar] [CrossRef]
- Acquaviva, R.; Genovese, C.; Amodeo, A.; Tomasello, B.; Malfa, G.A.; Sorrenti, V.; Tempera, G.; Addamo, A.P.; Ragusa, S.; Rosa, T.; et al. Biological activities of Teucrium flavum L., Teucrium fruticans L., and Teucrium siculum rafin crude extracts. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2018, 152, 720–727. [Google Scholar] [CrossRef]
- Genovese, C.; D’Angeli, F.; Bellia, F.; Distefano, A.; Spampinato, M.; Attanasio, F.; Nicolosi, D.; Di Salvatore, V.; Tempera, G.; Lo Furno, D.; et al. In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics 2021, 10, 428. [Google Scholar] [CrossRef]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Nat. Prod. Res. 2021, 35, 2076–2081. [Google Scholar] [CrossRef] [PubMed]
- D’Angeli, F.; Malfa, G.A.; Garozzo, A.; Li Volti, G.; Genovese, C.; Stivala, A.; Nicolosi, D.; Attanasio, F.; Bellia, F.; Ronsisvalle, S.; et al. Antimicrobial, Antioxidant, and Cytotoxic Activities of Juglans regia L. Pellicle Extract. Antibiotics 2021, 10, 159. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; Acquaviva, R.; Ronsisvalle, S.; Tempera, G.; Antonio Malfa, G.; D’Angeli, F.; Ragusa, S.; Nicolosi, D. In vitro evaluation of biological activities of Orobanche crenata Forssk. leaves extract. Nat. Prod. Res. 2020, 34, 3234–3238. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; D’Angeli, F.; Attanasio, F.; Caserta, G.; Scarpaci, K.S.; Nicolosi, D. Phytochemical composition and biological activities of Orobanche crenata Forssk.: A review. Nat. Prod. Res. 2020, 1–17. [Google Scholar] [CrossRef]
- Pereira, A.M.; Hernandes, C.; Pereira, S.I.; Bertoni, B.W.; Franca, S.C.; Pereira, P.S.; Taleb-Contini, S.H. Evaluation of anticandidal and antioxidant activities of phenolic compounds from Pyrostegia venusta (Ker Gawl.) Miers. Chem. Biol. Interact. 2014, 224, 136–141. [Google Scholar] [CrossRef]
- Kahraman, C.; Tatli, I.I.; Kart, D.; Ekizoglu, M.; Akdemir, Z.S. Structure Elucidation and Antimicrobial Activities of Secondary Metabolites from the Flowery Parts of Verbascum mucronatum Lam. Turk. J. Pharm. Sci. 2018, 15, 231–237. [Google Scholar] [CrossRef]
- Ali, I.; Sharma, P.; Suri, K.A.; Satti, N.K.; Dutt, P.; Afrin, F.; Khan, I.A. In vitro antifungal activities of amphotericin B in combination with acteoside, a phenylethanoid glycoside from Colebrookea oppositifolia. J. Med. Microbiol. 2011, 60, 1326–1336. [Google Scholar] [CrossRef]
- Chen, Q.; Xi, X.; Zeng, Y.; He, Z.; Zhao, J.; Li, Y. Acteoside inhibits autophagic apoptosis of retinal ganglion cells to rescue glaucoma-induced optic atrophy. J. Cell. Biochem. 2019, 120, 13133–13140. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.; Chen, Q.; Ma, J.; Wang, X.; Xia, Y.; Wen, X.; Cai, B.; Yan, L. Acteoside Protects Retinal Ganglion Cells from Experimental Glaucoma by Activating PI3K/AKT Signaling Pathway via Upregulating Caveolin 1. 2021. Available online: https://ssrn.com/abstract=3925505 (accessed on 20 October 2021).
- Wu, M.R.; Lin, C.H.; Ho, J.D.; Hsiao, G.; Cheng, Y.W. Novel Protective Effects of Cistanche Tubulosa Extract Against Low-Luminance Blue Light-Induced Degenerative Retinopathy. Cell. Physiol. Biochem. 2018, 51, 63–79. [Google Scholar] [CrossRef]
- Lee, H.; Woo, E.R.; Lee, D.G. Apigenin induces cell shrinkage in Candida albicans by membrane perturbation. FEMS Yeast Res. 2018, 18, foy003. [Google Scholar] [CrossRef]
- Lee, W.; Woo, E.R.; Lee, D.G. Effect of apigenin isolated from Aster yomena against Candida albicans: Apigenin-triggered apoptotic pathway regulated by mitochondrial calcium signaling. J. Ethnopharmacol. 2019, 231, 19–28. [Google Scholar] [CrossRef]
- Yordanov, M.; Dimitrova, P.; Patkar, S.; Saso, L.; Ivanovska, N. Inhibition of Candida albicans extracellular enzyme activity by selected natural substances and their application in Candida infection. Can. J. Microbiol. 2008, 54, 435–440. [Google Scholar] [CrossRef]
- Ivanov, M.; Kannan, A.; Stojkovic, D.S.; Glamoclija, J.; Calhelha, R.C.; Ferreira, I.; Sanglard, D.; Sokovic, M. Flavones, Flavonols, and Glycosylated Derivatives-Impact on Candida albicans Growth and Virulence, Expression of CDR1 and ERG11, Cytotoxicity. Pharmaceuticals 2020, 14, 27. [Google Scholar] [CrossRef]
- Jiang, W.; Chen, H.; Tai, Z.; Li, T.; Luo, L.; Tong, Z.; Zhu, W. Apigenin and Ethaverine Hydrochloride Enhance Retinal Vascular Barrier In Vitro and In Vivo. Transl. Vis. Sci. Technol. 2020, 9, 8. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Yu, H.; Li, M.; Hang, L.; Xu, X. Apigenin Protects Mouse Retina against Oxidative Damage by Regulating the Nrf2 Pathway and Autophagy. Oxid. Med. Cell. Longev. 2020, 2020, 9420704. [Google Scholar] [CrossRef]
- Chumsakul, O.; Wakayama, K.; Tsuhako, A.; Baba, Y.; Takai, Y.; Kurose, T.; Honma, Y.; Watanabe, S. Apigenin Regulates Activation of Microglia and Counteracts Retinal Degeneration. J. Ocul. Pharmacol. Ther. 2020, 36, 311–319. [Google Scholar] [CrossRef]
- Hytti, M.; Piippo, N.; Korhonen, E.; Honkakoski, P.; Kaarniranta, K.; Kauppinen, A. Fisetin and luteolin protect human retinal pigment epithelial cells from oxidative stress-induced cell death and regulate inflammation. Sci. Rep. 2015, 5, 17645. [Google Scholar] [CrossRef]
- Park, S.W.; Cho, C.S.; Jun, H.O.; Ryu, N.H.; Kim, J.H.; Yu, Y.S.; Kim, J.S.; Kim, J.H. Anti-angiogenic effect of luteolin on retinal neovascularization via blockade of reactive oxygen species production. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7718–7726. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.E.; Chen, Y.C.; Sun, X.B.; Tong, B.; Fan, X.H. Effects of luteolin on retinal oxidative stress and inflammation in diabetes. RSC Adv. 2015, 5, 4898–4904. [Google Scholar] [CrossRef]
- Liu, X.B.; Liu, F.; Liang, Y.Y.; Yin, G.; Zhang, H.J.; Mi, X.S.; Zhang, Z.J.; So, K.F.; Li, A.; Xu, Y. Luteolin delays photoreceptor degeneration in a mouse model of retinitis pigmentosa. Neural Regen. Res. 2021, 16, 2109–2120. [Google Scholar] [CrossRef]
- Huang, W.C.; Liou, C.J.; Shen, S.C.; Hu, S.; Hsiao, C.Y.; Wu, S.J. Luteolin Attenuates IL-1beta-Induced THP-1 Adhesion to ARPE-19 Cells via Suppression of NF-kappaB and MAPK Pathways. Mediat. Inflamm. 2020, 2020, 9421340. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, D.; Tian, D. Salidroside prevents hydroperoxide-induced oxidative stress and apoptosis in retinal pigment epithelium cells. Exp. Ther. Med. 2018, 16, 2363–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, K.; Wang, X.; Zhu, J.; Cao, G.; Zhang, K.; Su, Z. Salidroside protects retinal endothelial cells against hydrogen peroxide-induced injury via modulating oxidative status and apoptosis. Biosci. Biotechnol. Biochem. 2015, 79, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Dong, N.; Qiu, Q.; Li, S.; Zhang, J. Salidroside Prevents Hypoxia-Induced Human Retinal Microvascular Endothelial Cell Damage Via miR-138/ROBO4 Axis. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef]
- Ning, Z.; Hongdan, Y.; Zhen, F.; Jiayuan, D.; Xuezheng, L. Salidroside inhibits apoptosis of retinal Müller cells induced by high glucose in rats. Chin. J. Tissue Eng. Res. 2021, 25, 1664–1669. [Google Scholar] [CrossRef]
- Kumamoto, C.A.; Gresnigt, M.S.; Hube, B. The gut, the bad and the harmless: Candida albicans as a commensal and opportunistic pathogen in the intestine. Curr. Opin. Microbiol. 2020, 56, 7–15. [Google Scholar] [CrossRef]
- Gabaldon, T.; Gomez-Molero, E.; Bader, O. Molecular Typing of Candida glabrata. Mycopathologia 2020, 185, 755–764. [Google Scholar] [CrossRef]
- Song, G.; Liang, G.; Liu, W. Fungal Co-infections Associated with Global COVID-19 Pandemic: A Clinical and Diagnostic Perspective from China. Mycopathologia 2020, 185, 599–606. [Google Scholar] [CrossRef]
- Peman, J.; Ruiz-Gaitan, A.; Garcia-Vidal, C.; Salavert, M.; Ramirez, P.; Puchades, F.; Garcia-Hita, M.; Alastruey-Izquierdo, A.; Quindos, G. Fungal co-infection in COVID-19 patients: Should we be concerned? Rev. Iberoam. Micol. 2020, 37, 41–46. [Google Scholar] [CrossRef]
- Peng, J.; Wang, Q.; Mei, H.; Zheng, H.; Liang, G.; She, X.; Liu, W. Fungal co-infection in COVID-19 patients: Evidence from a systematic review and meta-analysis. Aging 2021, 13, 7745–7757. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Nair, R.; Banerjee, A. Emerging Mechanisms of Drug Resistance in Candida albicans. Prog. Mol. Subcell. Biol. 2019, 58, 135–153. [Google Scholar] [CrossRef]
- Revankar, S.G.; Kirkpatrick, W.R.; McAtee, R.K.; Dib, O.P.; Fothergill, A.W.; Redding, S.W.; Rinaldi, M.G.; Patterson, T.F. Detection and significance of fluconazole resistance in oropharyngeal candidiasis in human immunodeficiency virus-infected patients. J. Infect. Dis. 1996, 174, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Law, D.; Moore, C.B.; Wardle, H.M.; Ganguli, L.A.; Keaney, M.G.; Denning, D.W. High prevalence of antifungal resistance in Candida spp. From patients with AIDS. J. Antimicrob. Chemother. 1994, 34, 659–668. [Google Scholar] [CrossRef]
- Marr, K.A.; Seidel, K.; White, T.C.; Bowden, R.A. Candidemia in allogeneic blood and marrow transplant recipients: Evolution of risk factors after the adoption of prophylactic fluconazole. J. Infect. Dis. 2000, 181, 309–316. [Google Scholar] [CrossRef]
- Lewis, J.S., 2nd; Wiederhold, N.P.; Wickes, B.L.; Patterson, T.F.; Jorgensen, J.H. Rapid emergence of echinocandin resistance in Candida glabrata resulting in clinical and microbiologic failure. Antimicrob. Agents Chemother. 2013, 57, 4559–4561. [Google Scholar] [CrossRef] [Green Version]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Chandra, G.; Mukherjee, D.; Ray, A.S.; Chatterjee, S.; Bhattacharjee, I. Phytoextracts as Antibacterials: A Review. Curr. Drug Discov. Technol. 2020, 17, 523–533. [Google Scholar] [CrossRef]
- Kokoska, L.; Kloucek, P.; Leuner, O.; Novy, P. Plant-Derived Products as Antibacterial and Antifungal Agents in Human Health Care. Curr. Med. Chem. 2019, 26, 5501–5541. [Google Scholar] [CrossRef]
- Teodoro, G.R.; Ellepola, K.; Seneviratne, C.J.; Koga-Ito, C.Y. Potential Use of Phenolic Acids as Anti-Candida Agents: A Review. Front. Microbiol. 2015, 6, 1420. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V.; Efferth, T. Cameroonian medicinal plants: Pharmacology and derived natural products. Front. Pharmacol. 2010, 1, 123. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.R.; Kuete, V.; Jager, A.K.; Meyer, J.J.; Lall, N. Antimicrobial activity of selected South African medicinal plants. BMC Complement. Altern. Med. 2012, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- D’Enfert, C.; Janbon, G. Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches? FEMS Yeast Res. 2016, 16, fov111. [Google Scholar] [CrossRef] [Green Version]
- Douglas, L.J. Medical importance of biofilms in Candida infections. Rev. Iberoam. Micol. 2002, 19, 139–143. [Google Scholar]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Alnuaimi, A.D.; O’Brien-Simpson, N.M.; Reynolds, E.C.; McCullough, M.J. Clinical isolates and laboratory reference Candida species and strains have varying abilities to form biofilms. FEMS Yeast Res. 2013, 13, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J. Biphasic dose responses in biology, toxicology and medicine: Accounting for their generalizability and quantitative features. Environ. Pollut. 2013, 182, 452–460. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Dhawan, G.; Kapoor, R.; Iavicoli, I.; Calabrese, V. HORMESIS: A Fundamental Concept with Widespread Biological and Biomedical Applications. Gerontology 2016, 62, 530–535. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Iavicoli, I.; Calabrese, V. Hormesis: Its impact on medicine and health. Hum. Exp. Toxicol. 2013, 32, 120–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Calabrese, E.J.; Lian, B.; Lin, Z.; Calabrese, V. Hormesis as a mechanistic approach to understanding herbal treatments in traditional Chinese medicine. Pharmacol. Ther. 2018, 184, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Southam, C.M.; Erlich, J. Effects of extract of western red-cedar heartwood on certain wood-decaying fungi in culture. Phytopathology 1943, 33, 517–524. [Google Scholar]
- Liu, Y.; Wu, Z.; Feng, S.; Yang, X.; Huang, D. Hormesis of glyceollin I, an induced phytoalexin from soybean, on budding yeast chronological lifespan extension. Molecules 2014, 19, 568–580. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kwon, D.Y.; Cho, H.J.; Lee, M.S. Protective effects of Korean herbal remedy against oxidative stress in cardiomyocytes. Phytother. Res. 2006, 20, 235–236. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Baliga, M.S. Evaluation of the radioprotective action of geriforte in mice exposed to different doses of gamma-radiation. Am. J. Chin. Med. 2004, 32, 551–567. [Google Scholar] [CrossRef]
- Fu, Y.T.; Chen, K.Y.; Chen, Y.S.; Yao, C.H. Earthworm (Pheretima aspergillum) extract stimulates osteoblast activity and inhibits osteoclast differentiation. BMC Complement. Altern. Med. 2014, 14, 440. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, K.E.; Carter, D.A. Cellular plasticity of pathogenic fungi during infection. PLoS Pathog. 2020, 16, e1008571. [Google Scholar] [CrossRef]
- Raut, J.S.; Rajput, S.B.; Shinde, R.B.; Surwase, B.S.; Karuppayil, S.M.J.J.o.B.A.P.f.N. Vanillin inhibits growth, morphogenesis and biofilm formation by Candida albicans. J. Biol. Act. Prod. Nat. 2013, 3, 130–138. [Google Scholar]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Phenylpropanoids of Plant Origin as Inhibitors of Biofilm Formation by Candida albicans. J. Microbiol. Biotechnol. 2014, 24, 1216–1225. [Google Scholar] [CrossRef] [Green Version]
- Oliva, M.; Bar, E.; Ovadia, R.; Perl, A.; Galili, G.; Lewinsohn, E.; Oren-Shamir, M. Phenylpyruvate Contributes to the Synthesis of Fragrant Benzenoid-Phenylpropanoids in Petunia × hybrida Flowers. Front. Plant Sci. 2017, 8, 769. [Google Scholar] [CrossRef] [Green Version]
- Dini, I.; Iodice, C.; Ramundo, E. Phenolic metabolites from Orobanche speciosa. Planta Med. 1995, 61, 389–390. [Google Scholar] [CrossRef]
- Goyet, V.; Wada, S.; Cui, S.; Wakatake, T.; Shirasu, K.; Montiel, G.; Simier, P.; Yoshida, S. Haustorium Inducing Factors for Parasitic Orobanchaceae. Front. Plant Sci. 2019, 10, 1056. [Google Scholar] [CrossRef] [Green Version]
- Shah, C.P.; McKey, J.; Spirn, M.J.; Maguire, J. Ocular candidiasis: A review. Br. J. Ophthalmol. 2008, 92, 466–468. [Google Scholar] [CrossRef]
- Cascorbi, I.; Bittrich, H.; Ricklinkat, J.; Voss, W.; Seyfarth, A.; Foret, M. Effects of a heterogeneous set of xenobiotics on growth and plasma membranes of mammalian and fungal cell cultures. Ecotoxicol. Environ. Saf. 1993, 26, 113–126. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: A fundamental concept in biology. Microb. Cell. 2014, 1, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Demirovic, D.; Rattan, S.I. Curcumin induces stress response and hormetically modulates wound healing ability of human skin fibroblasts undergoing ageing in vitro. Biogerontology 2011, 12, 437–444. [Google Scholar] [CrossRef]
- Souza, L.; Silva-Rocha, W.P.; Ferreira, M.R.A.; Soares, L.A.L.; Svidzinski, T.I.E.; Milan, E.P.; Pires, R.H.; Fusco Almeida, A.M.; Mendes-Giannini, M.J.S.; Maranhao Chaves, G. Influence of Eugenia uniflora Extract on Adhesion to Human Buccal Epithelial Cells, Biofilm Formation, and Cell Surface Hydrophobicity of Candida spp. from the Oral Cavity of Kidney Transplant Recipients. Molecules 2018, 23, 2418. [Google Scholar] [CrossRef] [Green Version]
- Chrzanowski, L.; Bielicka-Daszkiewicz, K.; Owsianiak, M.; Aurich, A.; Kaczorek, E.; Olszanowski, A. Phenol and n-alkanes (C-12 and C-16) utilization: Influence on yeast cell surface hydrophobicity. World J. Microb. Biotechnol. 2008, 24, 1943–1949. [Google Scholar] [CrossRef]
- Kimani, B.G.; Kerekes, E.B.; Szebenyi, C.; Krisch, J.; Vagvolgyi, C.; Papp, T.; Tako, M. In Vitro Activity of Selected Phenolic Compounds against Planktonic and Biofilm Cells of Food-Contaminating Yeasts. Foods 2021, 10, 1652. [Google Scholar] [CrossRef]
- de Paula, S.B.; Bartelli, T.F.; Di Raimo, V.; Santos, J.P.; Morey, A.T.; Bosini, M.A.; Nakamura, C.V.; Yamauchi, L.M.; Yamada-Ogatta, S.F. Effect of Eugenol on Cell Surface Hydrophobicity, Adhesion, and Biofilm of Candida tropicalis and Candida dubliniensis Isolated from Oral Cavity of HIV-Infected Patients. Evid. Based Complement. Altern. Med. 2014, 2014, 505204. [Google Scholar] [CrossRef] [Green Version]
- Do, Q.D.; Angkawijaya, A.E.; Phuong, L.T.N.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Ben Attia, I.; Zucca, P.; Marincola, F.C.; Nieddu, M.; Piras, A.; Rosa, A.; Rescigno, A.; Chaieb, M. Evaluation of the Antioxidant and Cytotoxic Activities on Cancer Cell Line of Extracts of Parasitic Plants Harvested in Tunisia. Pol. J. Food Nutr. Sci. 2020, 70, 253–263. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). M27-A3: Reference Method for Broth Dilution Antifungal Susceptibiliy Testing of Yeasts, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Melo, A.S.; Bizerra, F.C.; Freymuller, E.; Arthington-Skaggs, B.A.; Colombo, A.L. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex. Med. Mycol. 2011, 49, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.A.; Fatima, Z.; Hameed, S. Anticandidal Effect and Mechanisms of Monoterpenoid, Perillyl Alcohol against Candida albicans. PLoS ONE 2016, 11, e0162465. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, I.; Felipe Rodrigues, M.P.; Bacelli, G.K.; Munin, E.; Alves, L.P.; Costa, M.S. Aloe vera extract reduces both growth and germ tube formation by Candida albicans. Mycoses 2012, 55, 257–261. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Timm, M.; Saaby, L.; Moesby, L.; Hansen, E.W. Considerations regarding use of solvents in in vitro cell based assays. Cytotechnology 2013, 65, 887–894. [Google Scholar] [CrossRef] [Green Version]
- Genovese, C.; Cambria, M.T.; D’Angeli, F.; Addamo, A.P.; Malfa, G.A.; Siracusa, L.; Pulvirenti, L.; Anfuso, C.D.; Lupo, G.; Salmeri, M. The double effect of walnut septum extract (Juglans regia L.) counteracts A172 glioblastoma cell survival and bacterial growth. Int. J. Oncol. 2020, 57, 1129–1144. [Google Scholar] [CrossRef]
- Motta, C.; D’Angeli, F.; Scalia, M.; Satriano, C.; Barbagallo, D.; Naletova, I.; Anfuso, C.D.; Lupo, G.; Spina-Purrello, V. PJ-34 inhibits PARP-1 expression and ERK phosphorylation in glioma-conditioned brain microvascular endothelial cells. Eur. J. Pharmacol. 2015, 761, 55–64. [Google Scholar] [CrossRef]
- Giurdanella, G.; Motta, C.; Muriana, S.; Arena, V.; Anfuso, C.D.; Lupo, G.; Alberghina, M. Cytosolic and calcium-independent phospholipase A(2) mediate glioma-enhanced proangiogenic activity of brain endothelial cells. Microvasc. Res. 2011, 81, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ammann, K.R.; DeCook, K.J.; Li, M.; Slepian, M.J. Migration versus proliferation as contributor to in vitro wound healing of vascular endothelial and smooth muscle cells. Exp. Cell. Res. 2019, 376, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Dalle, F.; Jouault, T.; Trinel, P.A.; Esnault, J.; Mallet, J.M.; d’Athis, P.; Poulain, D.; Bonnin, A. Beta-1,2- and alpha-1,2-linked oligomannosides mediate adherence of Candida albicans blastospores to human enterocytes in vitro. Infect. Immun. 2003, 71, 7061–7068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, T.; Liu, X.; Mao, P.; Wang, X.; Rui, C.; Yan, L.; Wang, Y.; Fan, C.; Li, P.; Zeng, X. The Role of 17beta-Estrogen in Candida albicans Adhesion on Human Vaginal Epithelial Cells via FAK Phosphorylation. Mycopathologia 2020, 185, 425–438. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Strains | OCLE 1 | Fluconazole | ||
|---|---|---|---|---|
| MFC50 2 | MIC50 3 | MIC50 | I.C. 4 | |
| Candida albicans ATCC 10231 | >293.55 | 146.77 | 16.00 | S-DD |
| Candida glabrata ATCC 2001 | >293.55 | 73.38 | 16.00 | S-DD |
| Fungal Strain | OCLE 1 (µg/mL) | ||||
|---|---|---|---|---|---|
| C+ 2 | 36.69 | 73.38 | 146.77 | 293.55 | |
| Candida albicans ATCC 10231 | - | - | + | ++ | ++ |
| Fungal Strains | Treatment | Number of Adhered Yeasts/100 Cells (% Adherence) | p-Value |
|---|---|---|---|
| Candida albicans ATCC 10231 | CTRL+ 1 | 31/100 (100%) | - |
| OCLE 2 146.77 µg/mL | 16/100 (51%) | p < 0.0001 | |
| OCLE 293.55 µg/mL | 0/100 (0%) | p < 0.0001 | |
| Candida glabrata ATCC 2001 | CTRL+ | 50/100 (100%) | - |
| OCLE 146.77 µg/mL | 0/100 (0%) | p < 0.0001 | |
| OCLE 293.55 µg/mL | 0/100 (0%) | p < 0.0001 |
| OCLE 1 (µg/mL) | ||||||
|---|---|---|---|---|---|---|
| 9.17 | 18.34 | 36.69 | 73.38 | 146.77 | 293.55 | |
| Total phenols content 2 | 186 ± 1.23 | 188 ± 1.89 | 194 ± 1.72 | 199 ± 0.44 | 203 ± 1.59 | 209 ± 3.79 |
| Chemical Name | Chemical Structure | m/z (g/mol) | Peak (Polarity) | RT * (min) | Biological Activities |
|---|---|---|---|---|---|
| 3-O-acetylepisamarcandin |  | 442.236 | 443.24 (+) 441.22 (−) | 38.62 | n.a. |
| Acteoside |  | 624.205 | 625.21 (+) 623.19 (−) | 23.23 | Anti-candidal [37,38,39] Anti-biofilm [39] Cytoprotective (retinal cells) [40,41,42] |
| Apigenin |  | 270.053 | 271.06 (+) 269.04 (−) | 35.97 | Anti-candidal [43,44,45,46] Anti-hyphal forming activity [46] Anti-biofilm [46] Cytoprotective (retinal cells) [47,48,49] |
| Asacoumarin A |  | 398.209 | 399.20 (+) 397.20 (−) | 37.98 | n.a. |
| Campneoside |  | 654.216 | n.r. | 43.27 | n.a. |
| Acutissimin A |  | 1206.822 | 1207.14 (+) 1205.10 (−) | n.r. | n.a. |
| Castacrenin F |  | 1218.145 | 1219.07 (+) 1217.10 (−) | n.r. | n.a. |
| Castacrenin C |  | 614.054 | 615.06 (+) 613.05 (−) | 36.96 | n.a. |
| Castacrenin E |  | 1056.072 | 1057.07 (+) 1055.06 (−) | 32.20 | n.a. |
| Chesnatin |  | 638.112 | 639.11 (+) 637.11 (−) | 28.46 | n.a. |
| Chestanin |  | 938.196 | 939.20 (+) 937.18 (−) | 43.62 | n.a. |
| Crenatoside |  | 622.190 | n.r. | 54.02 | n.a. |
| Dibutyl disulfide |  | 178.085 | 179.09 (+) 177.07 (−) | 39.53 | n.a. |
| Kurigalin |  | 636.096 | 637.10 (+) 635.10 (−) | 44.66 | n.a. |
| Leucosceptoside A |  | 638.221 | n.r. | 38.38 | n.a. |
| Luteolin |  | 286.048 | 287.05 (+) 285.03 (−) | 21.85 | Anti-candidal [46] Anti-hyphal forming activity [46] Anti-biofilm [46] Cytoprotective (retinal cells) [50,51,52,53,54] |
| Nevskin |  | 400.225 | 401.23 (+) 399.20 (−) | 14.06 | n.a. |
| Salidroside |  | 300.121 | 301.12 (+) 299.10 (−) | 35.10 | Cytoprotective (retinal cells) [55,56,57,58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Angeli, F.; Guadagni, F.; Genovese, C.; Nicolosi, D.; Trovato Salinaro, A.; Spampinato, M.; Mannino, G.; Lo Furno, D.; Petronio Petronio, G.; Ronsisvalle, S.; et al. Anti-Candidal Activity of the Parasitic Plant Orobanche crenata Forssk. Antibiotics 2021, 10, 1373. https://doi.org/10.3390/antibiotics10111373
D’Angeli F, Guadagni F, Genovese C, Nicolosi D, Trovato Salinaro A, Spampinato M, Mannino G, Lo Furno D, Petronio Petronio G, Ronsisvalle S, et al. Anti-Candidal Activity of the Parasitic Plant Orobanche crenata Forssk. Antibiotics. 2021; 10(11):1373. https://doi.org/10.3390/antibiotics10111373
Chicago/Turabian StyleD’Angeli, Floriana, Fiorella Guadagni, Carlo Genovese, Daria Nicolosi, Angela Trovato Salinaro, Mariarita Spampinato, Giuliana Mannino, Debora Lo Furno, Giulio Petronio Petronio, Simone Ronsisvalle, and et al. 2021. "Anti-Candidal Activity of the Parasitic Plant Orobanche crenata Forssk" Antibiotics 10, no. 11: 1373. https://doi.org/10.3390/antibiotics10111373
APA StyleD’Angeli, F., Guadagni, F., Genovese, C., Nicolosi, D., Trovato Salinaro, A., Spampinato, M., Mannino, G., Lo Furno, D., Petronio Petronio, G., Ronsisvalle, S., Sipala, F., Falzone, L., & Calabrese, V. (2021). Anti-Candidal Activity of the Parasitic Plant Orobanche crenata Forssk. Antibiotics, 10(11), 1373. https://doi.org/10.3390/antibiotics10111373