Genetic Features of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Poultry in Mayabeque Province, Cuba


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
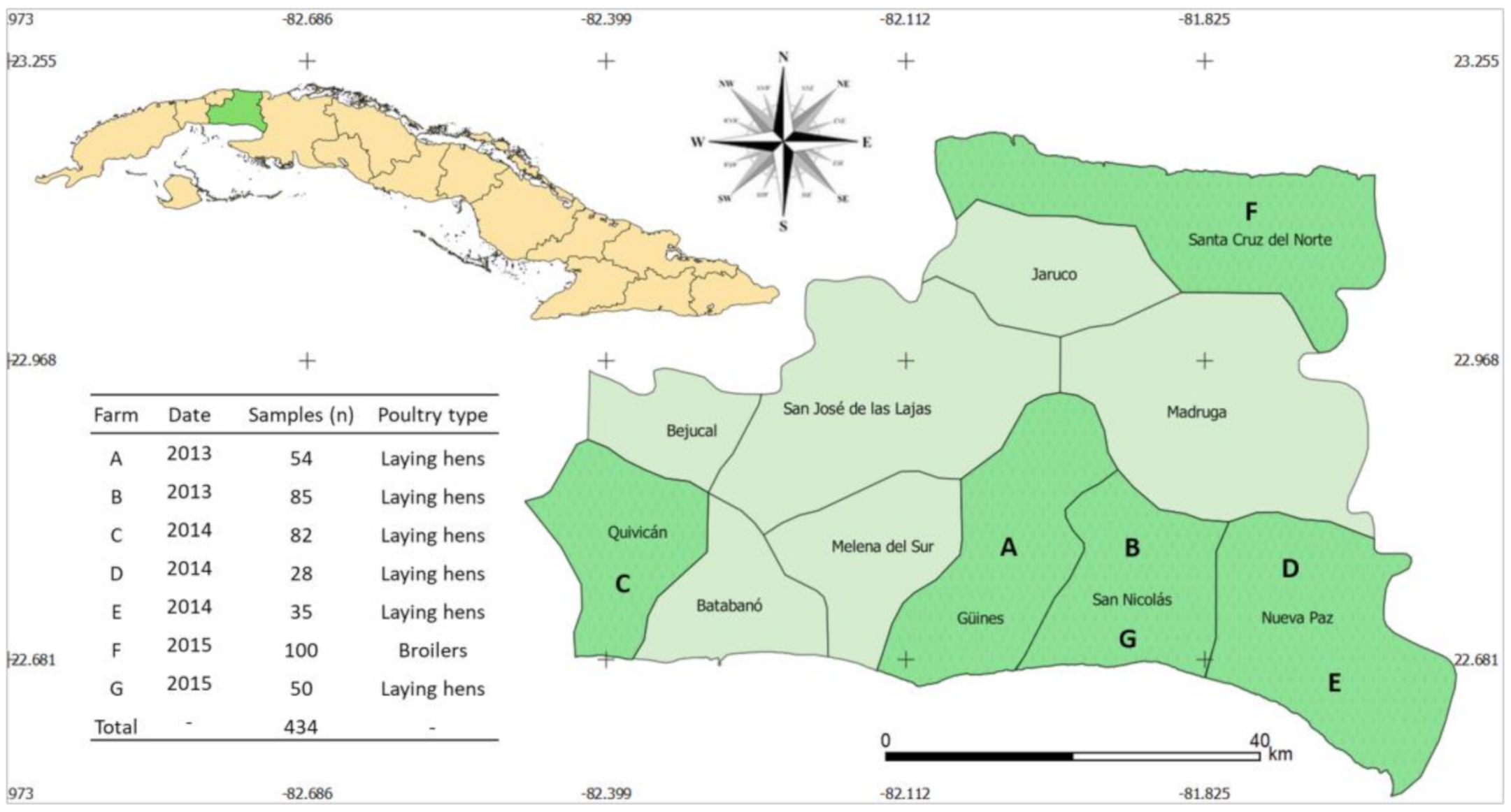
2.1. Distribution of 3GC-R-Ec
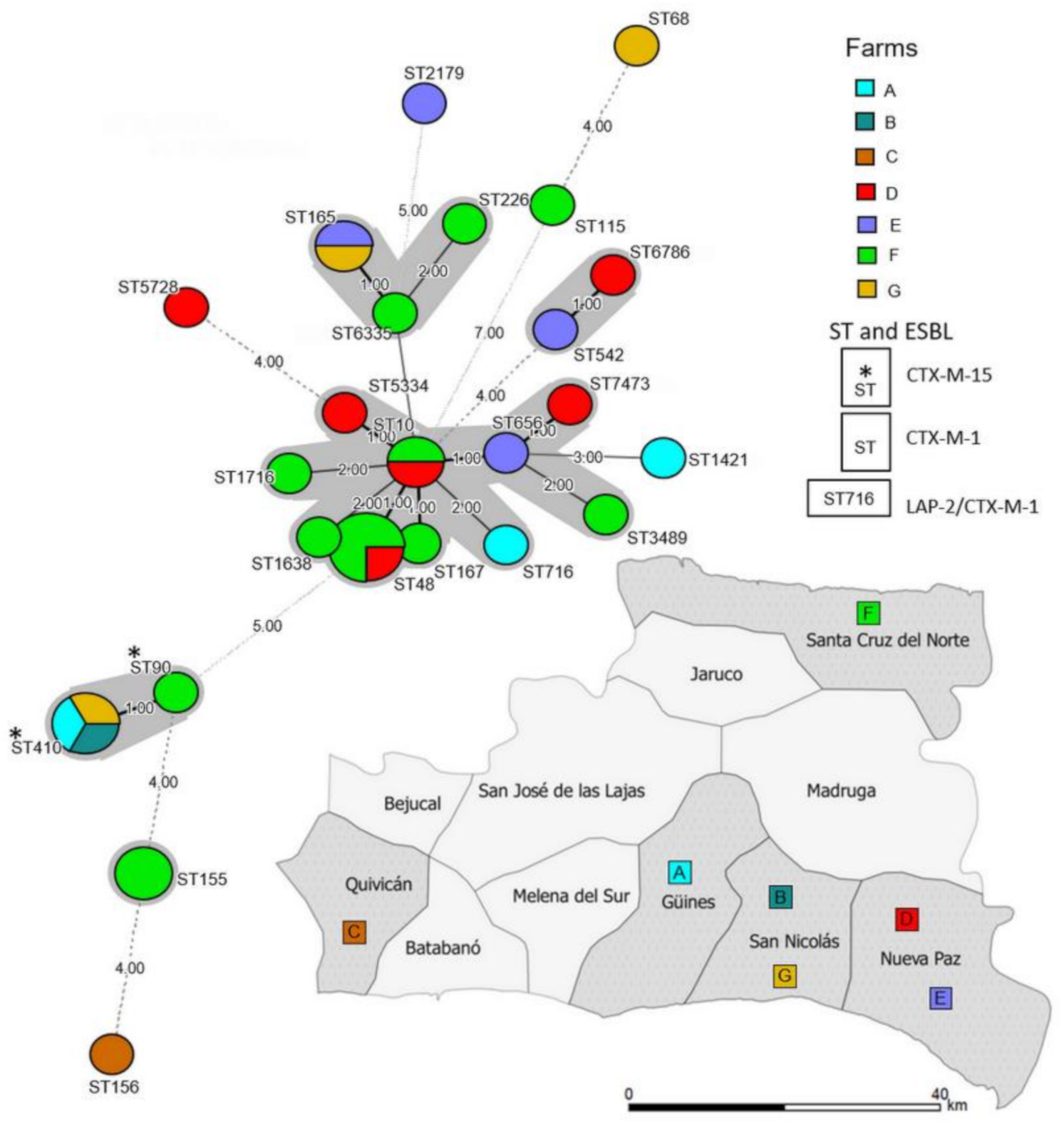
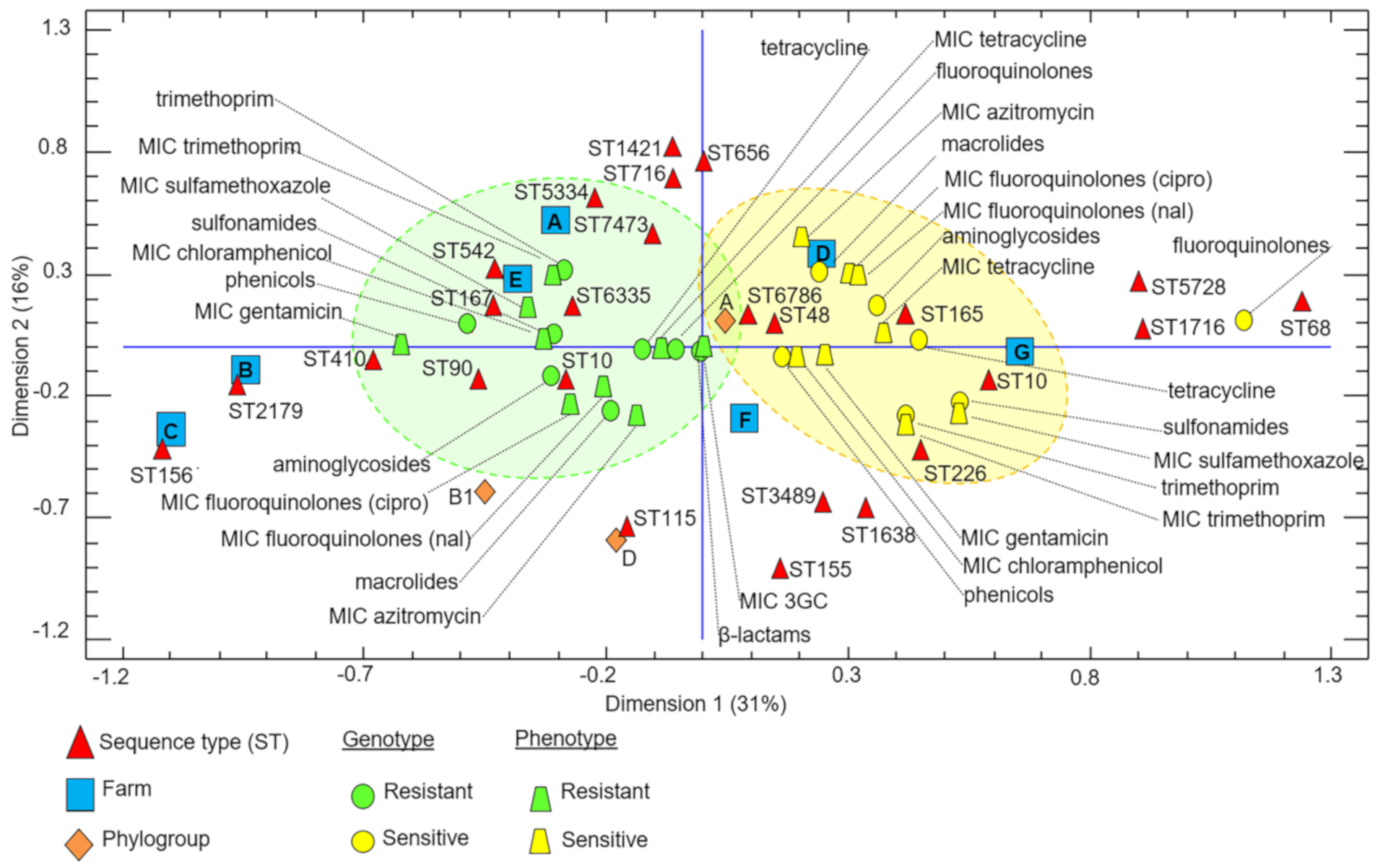
2.2. Genetic Diversity of the 3GC-R-Ec Isolates
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Isolation and Identification of 3GC-R-Ec
4.3. Rep-PCR-Based Genetic Relatedness and Selection of 3GCs-R-Ec Isolates
4.4. Antimicrobial Susceptibility Testing
4.5. Detection and Identification of Antibiotic-Resistance Genes
4.6. Multilocus Sequence Typing (MLST) and Determination of Phylogenetic Group
4.7. Pulsed-Field Gel Electrophoresis (PFGE)
4.8. PCR-Based Replicon Typing (PBRT) of Plasmids
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Ning, J.; Ahmed, S.; Huang, J.; Ullah, R.; An, B.; Hao, H.; Dai, M.; Huang, L.; Wang, X.; et al. Selection and dissemination of antimicrobial resistance in Agri-food production. Antimicrob. Resist. Infect. Control 2019, 8, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, R.H.; Bisgaard, M.; Lohren, U.; Robineau, B.; Christensen, H. Extended-spectrum beta-lactamase-producing Escherichia coli isolated from poultry: A review of current problems, illustrated with some laboratory findings. Avian Pathol. 2014, 43, 199–208. [Google Scholar] [CrossRef]
- Karaiskos, I.; Giamarellou, H. Carbapenem-sparing strategies for ESBL producers: When and how. Antibiotics 2020, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on the public health risks of bacterial strains producing extended-spectrum -lactamases and/or AmpC -lactamases in food and food-producing animals. Efsa J. 2011, 9, 2322. [Google Scholar] [CrossRef] [Green Version]
- Saliu, E.M.; Vahjen, W.; Zentek, J. Types and prevalence of extended-spectrum beta-lactamase producing Enterobacteriaceae in poultry. Anim. Health Res. Rev. 2017, 18, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Frömmel, U.; Lehmann, W.; Rödiger, S.; Böhm, A.; Nitschke, J.; Weinreich, J.; Gross, J.; Roggenbuck, D.; Zinke, O.; Ansorge, H.; et al. Adhesion of human and animal Escherichia coli strains in association with their virulence-associated genes and phylogenetic origins. Appl. Environ. Microbiol. 2013, 79, 5814–5829. [Google Scholar] [CrossRef] [Green Version]
- Ur Rahman, S.; Ali, T.; Ali, I. The growing genetic and functional diversity of extended spectrum beta-lactamases. Biomed. Res. Int. 2018, 2018, 9519718. [Google Scholar] [CrossRef]
- Giske, C.G.; Sundsfjord, A.S.; Kahlmeter, G.; Woodford, N.; Nordmann, P.; Paterson, D.L.; Canton, R.; Walsh, T.R. Redefining extended-spectrum β-lactamases: Balancing science and clinical need. J. Antimicrob. Chemother. 2009, 63, 1–4. [Google Scholar] [CrossRef]
- Zhao, W.-D.; Yan, P.; Guan, H.-N.; Zhang, Q.-Z. Characterization of CTX-M-type extended-spectrum beta-lactamase in clinical clones of Escherichia coli in Southwest China. J. Basic Microbiol. 2014, 54. [Google Scholar] [CrossRef]
- Blaak, H.; van Hoek, A.H.; Hamidjaja, R.A.; van der Plaats, R.Q.; Kerkhof-de Heer, L.; de Roda Husman, A.M.; Schets, F.M. Distribution, numbers, and diversity of ESBL-producing E. coli in the poultry farm environment. PLoS ONE 2015, 10, e0135402. [Google Scholar] [CrossRef]
- Kawamura, K.; Sugawara, T.; Matsuo, N.; Hayashi, K.; Norizuki, C.; Tamai, K.; Kondo, T.; Arakawa, Y. Spread of CTX-Type extended-spectrum β-lactamase-producing Escherichia coli isolates of epidemic clone B2-O25-ST131 among dogs and cats in Japan. Microb. Drug Resist. 2017, 23, 1059–1066. [Google Scholar] [CrossRef]
- Jacoby, G.A.; Strahilevitz, J.; Hooper, D.C. Plasmid-mediated quinolone resistance. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Chegene Lorestani, R.; Akya, A.; Elahi, A. The mutations of topoisomerase genes and their effect on resistance to fluoroquinolones in extended-spectrum β-lactamase-producing Escherichia coli. Jundishapur J. Nat. Pharm. Prod. 2018, 13, e57964. [Google Scholar] [CrossRef]
- Mora, A.; López, C.; Dabhi, G.; Blanco, M.; Blanco, J.E.; Alonso, M.P.; Herrera, A.; Mamani, R.; Bonacorsi, S.; Moulin-Schouleur, M.; et al. Extraintestinal pathogenic Escherichia coli O1:K1:H7/NM from human and avian origin: Detection of clonal groups B2 ST95 and D ST59 with different host distribution. BMC Microbiol. 2009, 9, 132. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Johnson, J.R. Reservoirs of extraintestinal pathogenic Escherichia coli. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Vogt, D.; Overesch, G.; Endimiani, A.; Collaud, A.; Thomann, A.; Perreten, V. Occurrence and genetic characteristics of third-generation cephalosporin-resistant Escherichia coli in Swiss retail meat. Microb. Drug Resist. 2014, 20, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Overdevest, I.; Willemsen, I.; Rijnsburger, M.; Eustace, A.; Xu, L.; Hawkey, P.; Heck, M.; Savelkoul, P.; Vandenbroucke-Grauls, C.; van der Zwaluw, K.; et al. Extended-spectrum beta-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands. Emerg. Infect. Dis. 2011, 17, 1216–1222. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D. Escherichia coli ST131: The quintessential example of an international multiresistant high-risk clone. Adv. Appl. Microbiol. 2015, 90, 109–154. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, S.N.; Carattoli, A.; Schwendener, S.; Collaud, A.; Endimiani, A.; Perreten, V. Plasmids carrying blaCMY-2/4 in Escherichia coli from poultry, poultry meat, and humans belong to a novel IncK subgroup designated IncK2. Front. Microbiol. 2017, 8, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousham, E.K.; Unicomb, L.; Islam, M.A. Human, animal and environmental contributors to antibiotic resistance in low-resource settings: Integrating behavioural, epidemiological and One Health approaches. Proc. Biol. Sci. 2018, 285, 20180332. [Google Scholar] [CrossRef] [PubMed]
- Schaufler, K.; Semmler, T.; Wieler, L.H.; Wohrmann, M.; Baddam, R.; Ahmed, N.; Muller, K.; Kola, A.; Fruth, A.; Ewers, C.; et al. Clonal spread and interspecies transmission of clinically relevant ESBL-producing Escherichia coli of ST410—Another successful pandemic clone? FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Guenther, S.; Ewers, C.; Wieler, L.H. Extended-spectrum Beta-lactamases producing E. coli in wildlife, yet another form of environmental pollution? Front. Microbiol. 2011, 2, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Schønning, K.; Wang, M.; Røder, B.L.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli sequence type 410 is causing new international high-risk clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Quiñones, D.; Aung, M.S.; Carmona, Y.; González, M.K.; Pereda, N.; Hidalgo, M.; Rivero, M.; Zayas, A.; Del Campo, R.; Urushibara, N.; et al. High prevalence of CTX-M type extended-spectrum beta-lactamase genes and detection of NDM-1 carbapenemase gene in extraintestinal pathogenic Escherichia coli in Cuba. Pathogens 2020, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Romeu, A.B.; Salazar, J.P.; Lugo, M.D.; Rojas, H.M.N.; Eslava, C.C.A. Antimicrobial susceptibility of Escherichia coli isolates from river water ecosystems. Rev. Cuba. Med Trop. 2012, 64, 132–141. [Google Scholar]
- Marrero-Moreno, C.M.; Mora-Llanes, M.; Hernández-Fillor, R.E.; Báez-Arias, M.; García-Morey, T.; Espinosa-Castaño, I. Identificación de enterobacterias productoras de betalactamasas de espectro extendido (BLEEs) en instalaciones porcinas de la provincia Matanzas. J. Rev. De Salud Anim. 2017, 39. Available online: http://scielo.sld.cu/scielo.php?script=sci_abstract&pid=S0253-570X2017000300006&lng=es&nrm=iso (accessed on 22 January 2021).
- González-Mesa, L.; González-Leyva, M.A.; Zayas-Tamayo, A.M.; Curbelo-Álvarez, M.; Garrido-Nicot, Y. Relación genética de aislados clínicos de Escherichia coli productores de Beta-Lactamasas de Espectro Extendido (BLEE) en un hospital de la Habana, Cuba. Rev. Cenic Cienc. Biol. 2017, 48, 107–111. [Google Scholar]
- Alonso, C.A.; Zarazaga, M.; Ben Sallem, R.; Jouini, A.; Ben Slama, K.; Torres, C. Antibiotic resistance in Escherichia coli in husbandry animals: The African perspective. Lett. Appl. Microbiol. 2017, 64, 318–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrijver, R.; Stijntjes, M.; Rodríguez-Baño, J.; Tacconelli, E.; Babu Rajendran, N.; Voss, A. Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 2018, 24, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.; Rokana, N.; Chandra, M.; Singh, B.P.; Gulhane, R.D.; Gill, J.P.S.; Ray, P.; Puniya, A.K.; Panwar, H. Antimicrobial resistance: Its surveillance, impact, and alternative management strategies in dairy animals. Front. Vet. Sci. 2018, 4, 237. [Google Scholar] [CrossRef]
- Escobar-Páramo, P.; Le Menac’h, A.; Le Gall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of forces shaping the commensal Escherichia coli genetic structure by comparing animal and human isolates. Environ. Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Adefioye, O.J.; Weinreich, J.; Rödiger, S.; Schierack, P.; Olowe, O.A. Phylogenetic characterization and multilocus sequence typing of extended-spectrum beta lactamase-producing Escherichia coli from food-producing animals, beef, and humans in Southwest Nigeria. Microb. Drug Resist. 2021, 27, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Stephan, R.; Nuesch-Inderbinen, M. Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Total Environ. 2016, 541, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Feng, Y.; Wu, W.; Xie, Y.; Wang, X.; Zhang, X.; Chen, X.; Zong, Z. First Report of OXA-181-producing Escherichia coli in China and characterization of the isolate using whole-genome sequencing. Antimicrob. Agents Chemother. 2015, 59, 5022–5025. [Google Scholar] [CrossRef] [Green Version]
- Piazza, A.; Comandatore, F.; Romeri, F.; Pagani, C.; Floriano, A.M.; Ridolfo, A.; Antona, C.; Brilli, M.; Mattioni Marchetti, V.; Bandi, C.; et al. First Report of an ST410 OXA-181 and CTX-M-15 coproducing Escherichia coli clone in Italy: A whole-genome sequence characterization. Microb. Drug Resist. 2018, 24, 1207–1209. [Google Scholar] [CrossRef]
- Blaak, H.; Hamidjaja, R.A.; van Hoek, A.H.; de Heer, L.; de Roda Husman, A.M.; Schets, F.M. Detection of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli on flies at poultry farms. Appl. Environ. Microbiol. 2014, 80, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahms, C.; Hubner, N.O.; Kossow, A.; Mellmann, A.; Dittmann, K.; Kramer, A. Occurrence of ESBL-producing Escherichia coli in livestock and farm workers in Mecklenburg-Western Pomerania, Germany. PLoS ONE 2015, 10, e0143326. [Google Scholar] [CrossRef] [PubMed]
- Endimiani, A.; Rossano, A.; Kunz, D.; Overesch, G.; Perreten, V. First countrywide survey of third-generation cephalosporin-resistant Escherichia coli from broilers, swine, and cattle in Switzerland. Diagn. Microbiol. Infect. Dis. 2012, 73, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.C.; Penha Filho, R.A.; Andrade, L.N.; Berchieri Junior, A.; Darini, A.L. Evaluation and characterization of plasmids carrying CTX-M genes in a non-clonal population of multidrug-resistant Enterobacteriaceae isolated from poultry in Brazil. Diagn. Microbiol. Infect. Dis. 2016, 85, 444–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinueza-Burgos, C.; Ortega-Paredes, D.; Narvaez, C.; De Zutter, L.; Zurita, J. Characterization of cefotaxime resistant Escherichia coli isolated from broiler farms in Ecuador. PLoS ONE 2019, 14, e0207567. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef] [PubMed]
- Dhanji, H.; Murphy, N.M.; Doumith, M.; Durmus, S.; Lee, S.S.; Hope, R.; Woodford, N.; Livermore, D.M. Cephalosporin resistance mechanisms in Escherichia coli isolated from raw chicken imported into the UK. J. Antimicrob. Chemother. 2010, 65, 2534–2537. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, L.R.; Donado-Godoy, P.; Leon, M.; Clavijo, V.; Arevalo, A.; Bernal, J.F.; Timmerman, A.J.; Mevius, D.J.; Wagenaar, J.A.; Hordijk, J. High heterogeneity of Escherichia coli sequence types harbouring ESBL/AmpC genes on IncI1 plasmids in the Colombian poultry chain. PLoS ONE 2017, 12, e0170777. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Pitout, J.D.D. Extended-spectrum β-lactamase-producing Enterobacteriaceae: Update on molecular epidemiology and treatment options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef]
- Lazarus, B.; Paterson, D.L.; Mollinger, J.L.; Rogers, B.A. Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin. Infect. Dis. 2015, 60, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Dame-Korevaar, A.; Fischer, E.A.J.; van der Goot, J.; Stegeman, A.; Mevius, D. Transmission routes of ESBL/pAmpC producing bacteria in the broiler production pyramid, a literature review. Prev. Vet. Med. 2019, 162, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and characterization of ESBL-producing Escherichia coli from humans and poultry in Ghana. Front. Microbiol. 2018, 9, 3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, D.C.; Lima, W.G.; de Paiva, M.C. Plasmid-mediated quinolone resistance (PMQR) among Enterobacteriales in Latin America: A systematic review. Mol. Biol. Rep. 2020, 47, 1471–1483. [Google Scholar] [CrossRef]
- Strahilevitz, J.; Jacoby, G.A.; Hooper, D.C.; Robicsek, A. Plasmid-mediated quinolone resistance: A multifaceted threat. Clin. Microbiol. Rev. 2009, 22, 664–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto de Medicina Veterinaria. Listado Oficial de Productos Veterinarios Registrados; Grupo De Registro De Productos De Uso Veterinario: La Habana, Cuba, 2018; pp. 1–138. [Google Scholar]
- Cantón, R.; Coque, T.M. The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Baquero, F.; Canton, R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Eurosurveillance 2008, 13, 19044. [Google Scholar] [CrossRef]
- Börjesson, S.; Bengtsson, B.; Jernberg, C.; Englund, S. Spread of extended-spectrum beta-lactamase producing Escherichia coli isolates in Swedish broilers mediated by an incl plasmid carrying blaCTX-M-1. Acta Vet. Scand. 2013, 55, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, V.L.; Scandorieiro, S.; Vespero, E.C.; Oba, A.; de Brito, B.G.; de Brito, K.C.; Nakazato, G.; Kobayashi, R.K. Comparison of antibiotic resistance and virulence factors among Escherichia coli isolated from conventional and free-range poultry. Biomed. Res. Int. 2015, 2015, 618752. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Review on Antimicrobial Resistance: London, UK, 2016; pp. 1–84. [Google Scholar]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J.; WHO Guideline Development Group. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrob. Resist. Infect. Control. 2018, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- World Organisation for Animal Health (OIE). Harmonisation of national antimicrobial resistance surveillance and monitoring programmes. In Terrestrial Animal Health Code, 19th ed.; OIE: Paris, France, 2010; Chapter 6.7; Available online: https://www.oie.int/en/international-standard-setting/terrestrial-code/access-online/?htmfile=chapitre_1.6.7.htm (accessed on 22 January 2021).
- Vila, J.; Marcos, M.A.; Jimenez de Anta, M.T. A comparative study of different PCR-based DNA fingerprinting techniques for typing of the Acinetobacter calcoaceticus-A. baumannii complex. J. Med. Microbiol. 1996, 44, 482–489. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 8.1; 2018. Available online: http://www.eucast.org (accessed on 22 January 2021).
- CLSI. CLSI Supplement M100, 27th ed.; Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Dahmen, S.; Poirel, L.; Mansour, W.; Bouallegue, O.; Nordmann, P. Prevalence of plasmid-mediated quinolone resistance determinants in Enterobacteriaceae from Tunisia. Clin. Microbiol. Infect. 2010, 16, 1019–1023. [Google Scholar] [CrossRef] [Green Version]
- Cano, M.E.; Rodríguez-Martínez, J.M.; Agüero, J.; Pascual, A.; Calvo, J.; García-Lobo, J.M.; Velasco, C.; Francia, M.V.; Martínez-Martínez, L. Detection of plasmid-mediated quinolone resistance genes in clinical isolates of Enterobacter spp. in Spain. J. Clin. Microbiol. 2009, 47, 2033–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; Fan, Y.; Agama Study Group; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baez, M.; Espinosa, I.; Collaud, A.; Miranda, I.; Montano, D.d.l.N.; Feria, A.L.; Hernández-Fillor, R.E.; Obregón, D.; Alfonso, P.; Perreten, V. Genetic Features of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Poultry in Mayabeque Province, Cuba. Antibiotics 2021, 10, 107. https://doi.org/10.3390/antibiotics10020107
Baez M, Espinosa I, Collaud A, Miranda I, Montano DdlN, Feria AL, Hernández-Fillor RE, Obregón D, Alfonso P, Perreten V. Genetic Features of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Poultry in Mayabeque Province, Cuba. Antibiotics. 2021; 10(2):107. https://doi.org/10.3390/antibiotics10020107
Chicago/Turabian StyleBaez, Michel, Ivette Espinosa, Alexandra Collaud, Iliana Miranda, Damarys de las Nieves Montano, Angel L. Feria, Rosa Elena Hernández-Fillor, Dasiel Obregón, Pastor Alfonso, and Vincent Perreten. 2021. "Genetic Features of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Poultry in Mayabeque Province, Cuba" Antibiotics 10, no. 2: 107. https://doi.org/10.3390/antibiotics10020107
APA StyleBaez, M., Espinosa, I., Collaud, A., Miranda, I., Montano, D. d. l. N., Feria, A. L., Hernández-Fillor, R. E., Obregón, D., Alfonso, P., & Perreten, V. (2021). Genetic Features of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Poultry in Mayabeque Province, Cuba. Antibiotics, 10(2), 107. https://doi.org/10.3390/antibiotics10020107