
Existence of Multiple ESBL Genes among Phenotypically Confirmed ESBL Producing Klebsiella pneumoniae and Escherichia coli Concurrently Isolated from Clinical, Colonization and Contamination Samples from Neonatal Units at Bugando Medical Center, Mwanza, Tanzania

 , ,
, ,
Abstract
:1. Introduction
2. Results
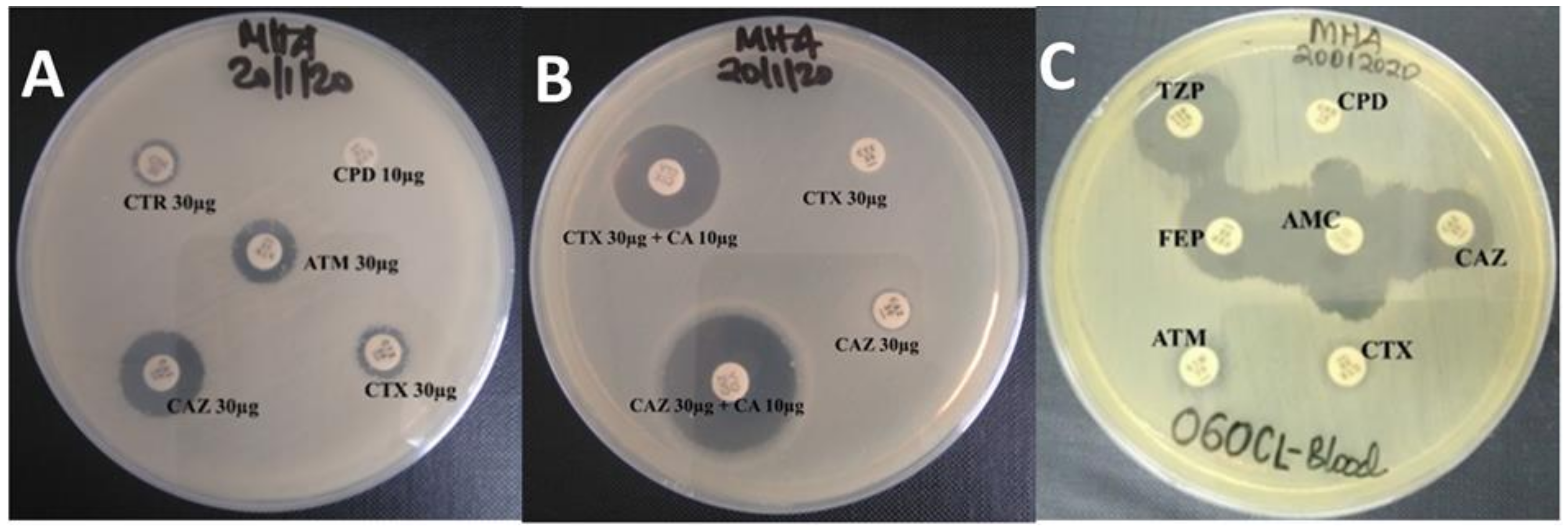
2.1. Phenotypic Screening and Confirmation of ESBL Production
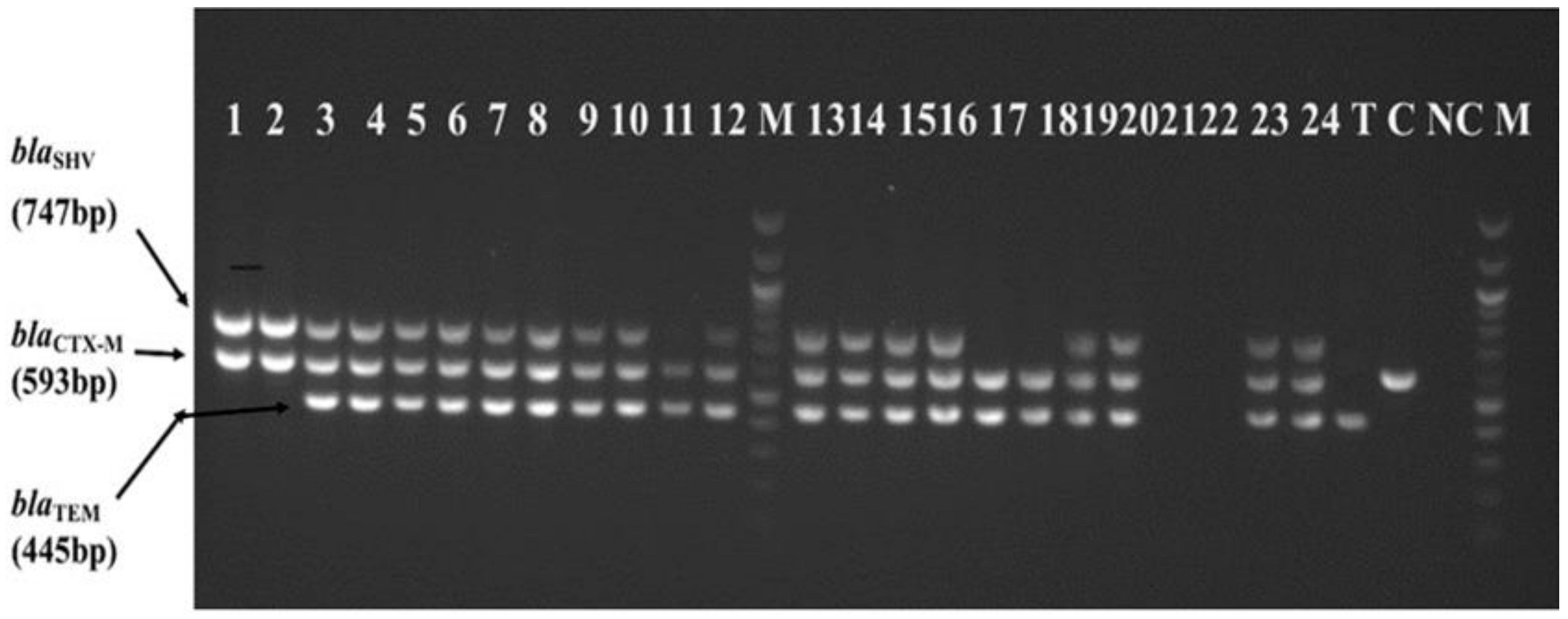
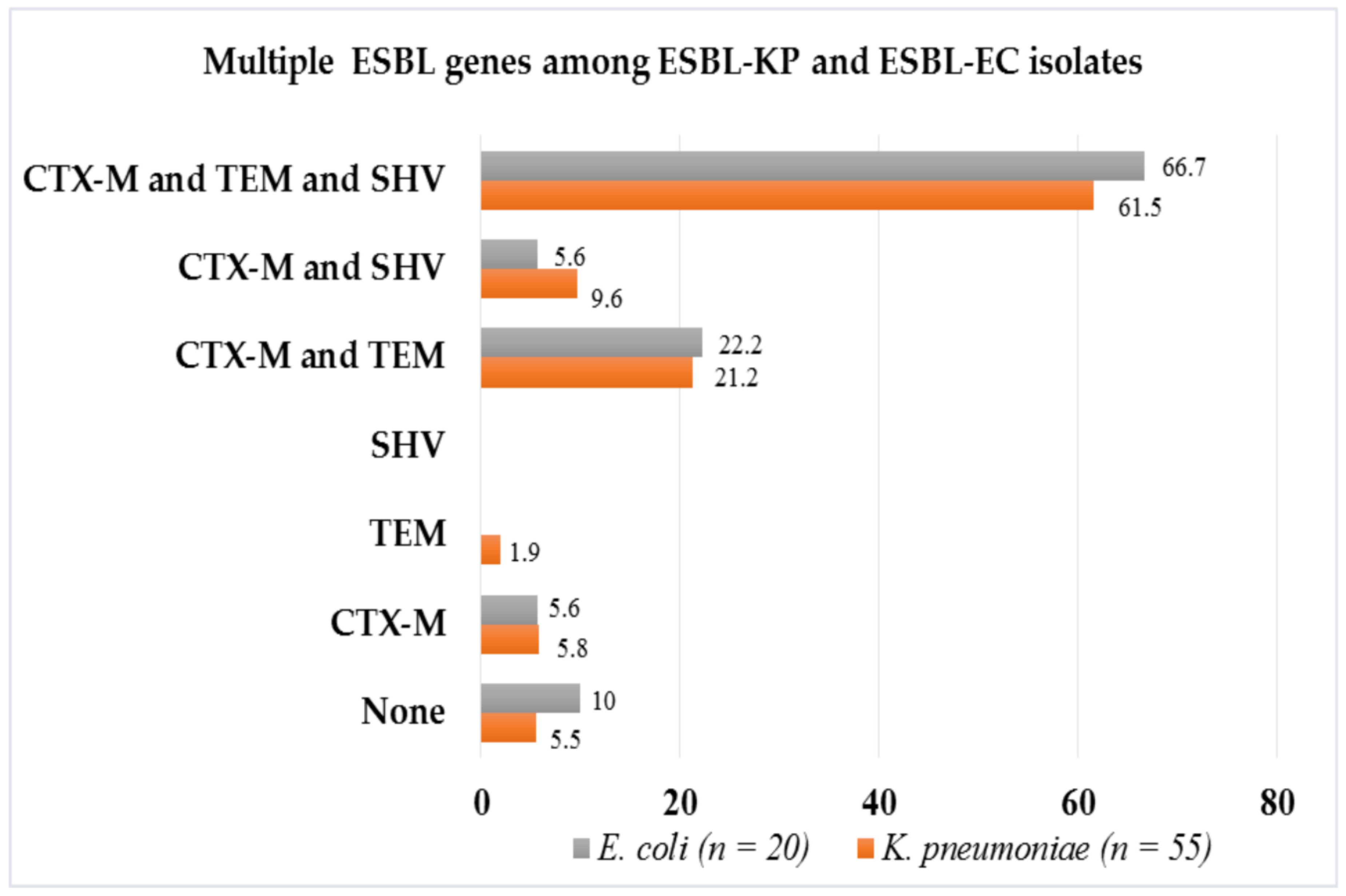
2.2. Multiple ESBL Genes Harbored by Phenotypically Confirmed ESBL-KP and ESBL-EC
2.3. Combination of ESBL Genes in Association with Resistance to None β-Lactam Antibiotics and Meropenem
2.4. Pairs of ESBL Isolates Showing Similar Patterns of Antibiotics Susceptibility and ESBL Genes
3. Discussion
4. Materials and Methods
4.1. Study Design, Duration, Setting and Population
4.2. Isolates Recovery and Phenotypic Screening of ESBL Production
4.3. Phenotypic Confirmation of ESBL Production
4.3.1. CLSI Combination Disc Diffusion (CLSI-CDD) Method
4.3.2. Modified Double Disc Synergy (MDDS) Method
4.4. Antibiotic Susceptibility Testing (AST)
4.5. Molecular Characterization of ESBL Genes
4.5.1. DNA Extraction
4.5.2. Multiplex PCR Amplification
4.6. Quality Control
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in Gram negative bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 25 January 2021).
- Global Action Plan on Antimicrobial Resistance. Available online: https://apps.who.int/iris/bitstream/handle/10665/193736/9789241509763_eng.pdf;jsessionid=BF97F32FB0787262F4A0194D56EA2C0A?sequence=1 (accessed on 25 January 2021).
- Deane, J.; Rea, M.C.; Fouhy, F.; Stanton, C.; Ross, R.P.; Plant, B.J. Long-Term Implications of Antibiotic Use on Gut Health and Microbiota in Populations Including Patients With Cystic Fibrosis. In The Gut-Brain Axis; Elsevier: Amsterdam, The Netherlands, 2016; pp. 223–259. [Google Scholar]
- Lee, N.-Y.; Lee, C.-C.; Huang, W.-H.; Tsui, K.-C.; Hsueh, P.-R.; Ko, W.-C. Carbapenem therapy for bacteremia due to extended-spectrum-β-lactamase-producing Escherichia coli or Klebsiella pneumoniae: Implications of ertapenem susceptibility. Antimicrob. Agents Chemother. 2012, 56, 2888–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, P.A. Extended-spectrum β-lactamases in the 21st century: Characterization, epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, Y.; Archin, T.; Kargar, M.; Mohkam, M. A simple multiplex PCR for assessing prevalence of extended-spectrum β-lactamases producing Klebsiella pneumoniae in Intensive Care Units of a referral hospital in Shiraz, Iran. Asian Pac. J. Trop. Med. 2013, 6, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Jacoby, G.A. Updated functional classification of β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. In Clinical and Laboratory Standards Institute; CLSI: Wyne, PA, USA, 2018. [Google Scholar]
- Krishnamurthy, V.; Vijaykumar, G.S.; Kumar, S.; Prashanth, H.V.; Prakash, R.; Nagaraj, E.R. Phenotypic and genotypic methods for detection of extended spectrum β lactamase producing Escherichia coli and Klebsiella pneumoniae isolated from ventilator associated pneumonia. J. Clin. Diagn. Res. 2013, 7, 1975–1978. [Google Scholar] [CrossRef]
- Kayange, N.; Kamugisha, E.; Mwizamholya, D.L.; Jeremiah, S.; Mshana, S.E. Predictors of positive blood culture and deaths among neonates with suspected neonatal sepsis in a tertiary hospital, Mwanza-Tanzania. BMC Pediatrics 2010, 10, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marando, R.; Seni, J.; Mirambo, M.M.; Falgenhauer, L.; Moremi, N.; Mushi, M.F.; Kayange, N.; Manyama, F.; Imirzalioglu, C.; Chakraborty, T. Predictors of the extended-spectrum-beta lactamases producing Enterobacteriaceae neonatal sepsis at a tertiary hospital, Tanzania. Int. J. Med. Microbiol. 2018, 308, 803–811. [Google Scholar] [CrossRef]
- Mshana, S.E.; Falgenhauer, L.; Mirambo, M.M.; Mushi, M.F.; Moremi, N.; Julius, R.; Seni, J.; Imirzalioglu, C.; Matee, M.; Chakraborty, T. Predictors of bla CTX-M-15 in varieties of Escherichia coli genotypes from humans in community settings in Mwanza, Tanzania. BMC Infect. Dis. 2016, 16, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moremi, N.; Manda, E.V.; Falgenhauer, L.; Ghosh, H.; Imirzalioglu, C.; Matee, M.; Chakraborty, T.; Mshana, S.E. Predominance of CTX-M-15 among ESBL producers from environment and fish gut from the shores of Lake Victoria in Mwanza, Tanzania. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Moremi, N.; Claus, H.; Vogel, U.; Mshana, S.E. Faecal carriage of CTX-M extended-spectrum beta-lactamase-producing Enterobacteriaceae among street children dwelling in Mwanza city, Tanzania. PLoS ONE 2017, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mshana, S.; Imirzalioglu, C.; Hain, T.; Domann, E.; Lyamuya, E.; Chakraborty, T. Multiple ST clonal complexes, with a predominance of ST131, of Escherichia coli harbouring blaCTX-M-15 in a tertiary hospital in Tanzania. Clin. Microbiol. Infect. 2011, 17, 1279–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassen, B.; Sghaier, S.; Abbassi, M.S.; Ferjani, M.A.; Said, M.B.; Hassen, A.; Hammami, S. Multidrug resistance and the predominance of blaCTX-M in extended spectrum beta-lactamase-producing Enterobacteriaceae of animal and water origin. J. Mol. Microbiol. Biotechnol. 2018, 28, 201–206. [Google Scholar] [CrossRef]
- Park, H.; Kim, J.; Ryu, S.; Jeon, B. Predominance of blaCTX-M-65 and blaCTX-M-55 in extended-spectrum β-lactamase-producing Escherichia coli from raw retail chicken in South Korea. J. Glob. Antimicrob. Resist. 2019, 17, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Diab, A.M.; Abul-Aziz, M.; El-Kholy, I.; Rezk, M.A. Modified double-disc synergy test (MDDST) for detection of extended-spectrum beta-lactamases in AmpC beta-lactamase-producing Klebsiella clinical isolates. Eur. Chem. Bull. 2018, 7, 89–92. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI. Supplement M100; Clinical and Laboratory Standards Institute: Wyne, PA, USA, 2020. [Google Scholar]
- Ho, P.; Shek, R.H.; Chow, K.; Duan, R.; Mak, G.C.; Lai, E.L.; Yam, W.; Tsang, K.W.; Lai, W. Detection and characterization of extended-spectrum β-lactamases among bloodstream isolates of Enterobacter spp. in Hong Kong, 2000. J. Antimicrob. Chemother. 2005, 55, 326–332. [Google Scholar] [CrossRef]
- Asir, J.; Nair, S.; Devi, S.; Prashanth, K.; Saranathan, R.; Kanungo, R. Simultaneous gut colonisation and infection by ESBL-producing Escherichia coli in hospitalised patients. Australas. Med. J. 2015, 8, 200–207. [Google Scholar] [CrossRef]
- Ndugulile, F.; Jureen, R.; Harthug, S.; Urassa, W.; Langeland, N. Extended Spectrum β-Lactamases among Gram-negative bacteria of nosocomial origin from an Intensive Care Unit of a tertiary health facility in Tanzania. BMC Infect. Dis. 2005, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Seni, J.; Falgenhauer, L.; Simeo, N.; Mirambo, M.M.; Imirzalioglu, C.; Matee, M.; Rweyemamu, M.; Chakraborty, T.; Mshana, S.E. Multiple ESBL-producing Escherichia coli sequence types carrying quinolone and aminoglycoside resistance genes circulating in companion and domestic farm animals in Mwanza, Tanzania, harbor commonly occurring plasmids. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Paterson, D. Recommendation for treatment of severe infections caused by Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs). Clin. Microbiol. Infect. 2000, 6, 460–463. [Google Scholar] [CrossRef] [Green Version]
- Hawkey, P.M.; Jones, A.M. The changing epidemiology of resistance. J. Antimicrob. Chemother. 2009, 64, i3–i10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, S.; Fan, X.; Huang, Z.; Xia, L.; Xiao, M.; Chen, R.; Xu, Y.; Zhuo, C. Dominance of CTX-M-type extended-spectrum β-lactamase (ESBL)-producing Escherichia coli isolated from patients with community-onset and hospital-onset infection in China. PLoS ONE 2014, 9, e100707. [Google Scholar] [CrossRef]
- Rossolini, G.; D’Andrea, M.; Mugnaioli, C. The spread of CTX-M-type extended-spectrum β-lactamases. Clin. Microbiol. Infect. 2008, 14, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, C.; Li, X.-Q.; Zong, Z.-Y.; Zhong, N.-S. Epidemic plasmid carrying bla CTX-M-15 in Klebsiella penumoniae in China. PLoS ONE 2013, 8, e52222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, T.; Ali, I.; Khan, N.A.; Han, B.; Gao, J. The growing genetic and functional diversity of extended spectrum beta-lactamases. BioMed Res. Int. 2018, 2018, 1–14. [Google Scholar]
- Gómez-Zorrilla, S.; Camoez, M.; Tubau, F.; Cañizares, R.; Periche, E.; Dominguez, M.A.; Ariza, J.; Peña, C. Prospective observational study of prior rectal colonization status as a predictor for subsequent development of Pseudomonas aeruginosa clinical infections. Antimicrob. Agents Chemother. 2015, 59, 5213–5219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierro, A.; Van Saene, H.K.; Donnell, S.C.; Hughes, J.; Ewan, C.; Nunn, A.J.; Lloyd, D.A. Microbial translocation in neonates and infants receiving long-term parenteral nutrition. Arch. Surg. 1996, 131, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Pultz, N.J.; Gries, D.M.; Ray, A.J.; Eckstein, E.C.; Aron, D.C.; Donskey, C.J. Acquisition of nosocomial pathogens on hands after contact with environmental surfaces near hospitalized patients. Infect. Control Hosp. Epidemiol. 2004, 25, 164–167. [Google Scholar] [CrossRef]
- Mathur, P. Hand hygiene: Back to the basics of infection control. Indian J. Med. Res. 2011, 134, 611–620. [Google Scholar] [CrossRef]
- Silago, V.; Kovacs, D.; Msanga, D.R.; Seni, J.; Matthews, L.; Oravcová, K.; Zadoks, R.N.; Lupindu, A.M.; Hoza, A.S.; Mshana, S.E. Bacteremia in critical care units at Bugando Medical Centre, Mwanza, Tanzania: The role of colonization and contaminated cots and mothers’ hands in cross-transmission of multidrug resistant Gram-negative bacteria. Antimicrob. Resist. Infect. Control 2020, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Polsfuss, S.; Bloemberg, G.V.; Giger, J.; Meyer, V.; Böttger, E.C.; Hombach, M. Practical approach for reliable detection of AmpC beta-lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2011, 49, 2798–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monstein, H.J.; Östholm-Balkhed, Å.; Nilsson, M.V.; Nilsson, M.; Dornbusch, K.; Nilsson, L.E. Multiplex PCR amplification assay for the detection of blaSHV, blaTEM and blaCTX-M genes in Enterobacteriaceae. Apmis 2007, 115, 1400–1408. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic Agent | Patterns of ESBL Genes Combinations | |||||
|---|---|---|---|---|---|---|
| None | CTX-M or TEM | CTX-M and TEM | CTX-M and SHV | CTX-M, TEM and SHV | p Value | |
| SXT (n = 75) | 5 (6.7%) | 5 (6.7%) | 15 (20%) | 6 (8.0%) | 44 (58.7%) | - |
| CN (n = 59) | 4 (6.8%) | 5 (8.5%) | 10 (16.9%) | 5 (8.5%) | 35 (59.3%) | 0.601 |
| CIP (n = 26) | 2 (6.7%) | 2 (6.7%) | 7 (24.1%) | 3 (10.3%) | 15 (51.7%) | 0.836 |
| MEM (n = 4) | - | - | - | - | 4 (100%) | 0.562 |
| ID | Isolate | Source | MDDS | SXT | CN | CIP | MEM | Presence of bla Alleles |
|---|---|---|---|---|---|---|---|---|
| 070CL CL053 | K. pneumoniae | rectal swab | positive | 6 (R) | 8 (R) | 32 (S) | 30 (S) | CTX-M + TEM |
| K. pneumoniae | HCW hand | positive | 6 (R) | 6 (R) | 32 (S) | 34 (S) | CTX-M + TEM | |
| 231CL 249CL | E. coli | rectal swab | positive | 6 (R) | 14 (I) | 30 (S) | 30 (S) | CTX-M + TEM + SHV |
| K. pneumoniae | cot | positive | 6 (R) | 14 (I) | 30 (S) | 29 (S) | CTX-M + TEM + SHV | |
| 249CL | E. coli | rectal swab | positive | 6 (R) | 14 (I) | 28 (S) | 28 (S) | CTX-M + TEM + SHV |
| K. pneumoniae | cot | positive | 6 (R) | 14 (I) | 30 (S) | 29 (S) | CTX-M + TEM + SHV | |
| 249CL 250CL | K. pneumoniae | blood | positive | 6 (R) | 14 (I) | 28 (S) | 12 (R) | CTX-M + TEM + SHV |
| K. pneumoniae | blood | positive | 6 (R) | 14 (I) | 27 (S) | 11 (R) | CTX-M + TEM + SHV | |
| 275CL | K. pneumoniae | blood | positive | 6 (R) | 6 (R) | 17 (I) | 28 (S) | CTX-M + TEM |
| K. pneumoniae | rectal swab | positive | 6 (R) | 8 (R) | 20 (I) | 32 (S) | CTX-M + TEM | |
| 282CL | K. pneumoniae | blood | positive | 6 (R) | 6 (R) | 6 (R) | 30 (S) | CTX-M + TEM + SHV |
| K. pneumoniae | rectal swab | positive | 6 (R) | 6 (R) | 6 (R) | 30 (S) | CTX-M + TEM + SHV | |
| 285CL | K. pneumoniae | blood | positive | 12(R) | 6 (R) | 30 (S) | 30 (S) | CTX-M + TEM + SHV |
| K. pneumoniae | rectal swab | positive | 6 (R) | 6 (R) | 28 (S) | 30 (S) | CTX-M + TEM + SHV | |
| 387CL | K. pneumoniae | blood | positive | 6 (R) | 6 (R) | 19 (I) | 30 (S) | CTX-M + TEM + SHV |
| K. pneumoniae | rectal swab | positive | 6 (R) | 6 (R) | 15 (R) | 30 (S) | CTX-M + TEM + SHV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silago, V.; Kovacs, D.; Samson, H.; Seni, J.; Matthews, L.; Oravcová, K.; Lupindu, A.M.; Hoza, A.S.; Mshana, S.E. Existence of Multiple ESBL Genes among Phenotypically Confirmed ESBL Producing Klebsiella pneumoniae and Escherichia coli Concurrently Isolated from Clinical, Colonization and Contamination Samples from Neonatal Units at Bugando Medical Center, Mwanza, Tanzania. Antibiotics 2021, 10, 476. https://doi.org/10.3390/antibiotics10050476
Silago V, Kovacs D, Samson H, Seni J, Matthews L, Oravcová K, Lupindu AM, Hoza AS, Mshana SE. Existence of Multiple ESBL Genes among Phenotypically Confirmed ESBL Producing Klebsiella pneumoniae and Escherichia coli Concurrently Isolated from Clinical, Colonization and Contamination Samples from Neonatal Units at Bugando Medical Center, Mwanza, Tanzania. Antibiotics. 2021; 10(5):476. https://doi.org/10.3390/antibiotics10050476
Chicago/Turabian StyleSilago, Vitus, Dory Kovacs, Happyness Samson, Jeremiah Seni, Louise Matthews, Katarina Oravcová, Athumani M. Lupindu, Abubakar S. Hoza, and Stephen E. Mshana. 2021. "Existence of Multiple ESBL Genes among Phenotypically Confirmed ESBL Producing Klebsiella pneumoniae and Escherichia coli Concurrently Isolated from Clinical, Colonization and Contamination Samples from Neonatal Units at Bugando Medical Center, Mwanza, Tanzania" Antibiotics 10, no. 5: 476. https://doi.org/10.3390/antibiotics10050476
APA StyleSilago, V., Kovacs, D., Samson, H., Seni, J., Matthews, L., Oravcová, K., Lupindu, A. M., Hoza, A. S., & Mshana, S. E. (2021). Existence of Multiple ESBL Genes among Phenotypically Confirmed ESBL Producing Klebsiella pneumoniae and Escherichia coli Concurrently Isolated from Clinical, Colonization and Contamination Samples from Neonatal Units at Bugando Medical Center, Mwanza, Tanzania. Antibiotics, 10(5), 476. https://doi.org/10.3390/antibiotics10050476