
Apt (Adenine Phosphoribosyltransferase) Mutation in Laboratory-Selected Vancomycin-Intermediate Staphylococcus aureus




Abstract
:1. Introduction
2. Results and Discussion
2.1. Mutations Identified in MM66 VISA Mutants
2.2. General Aspects of the MM66-3 and MM66-4 Transcriptome
2.3. Altered Cell Wall Metabolism Genes in MM66 VISA
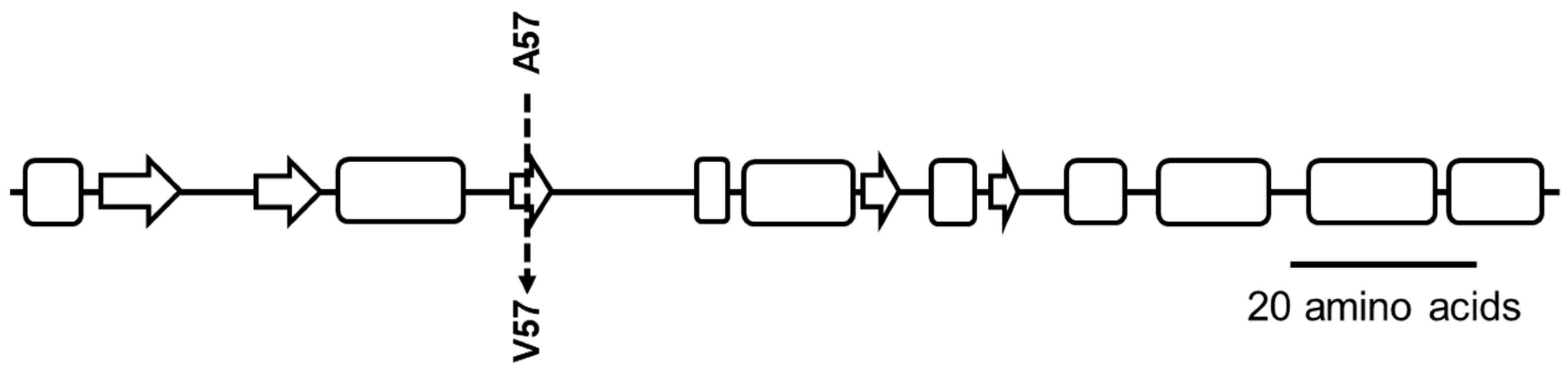
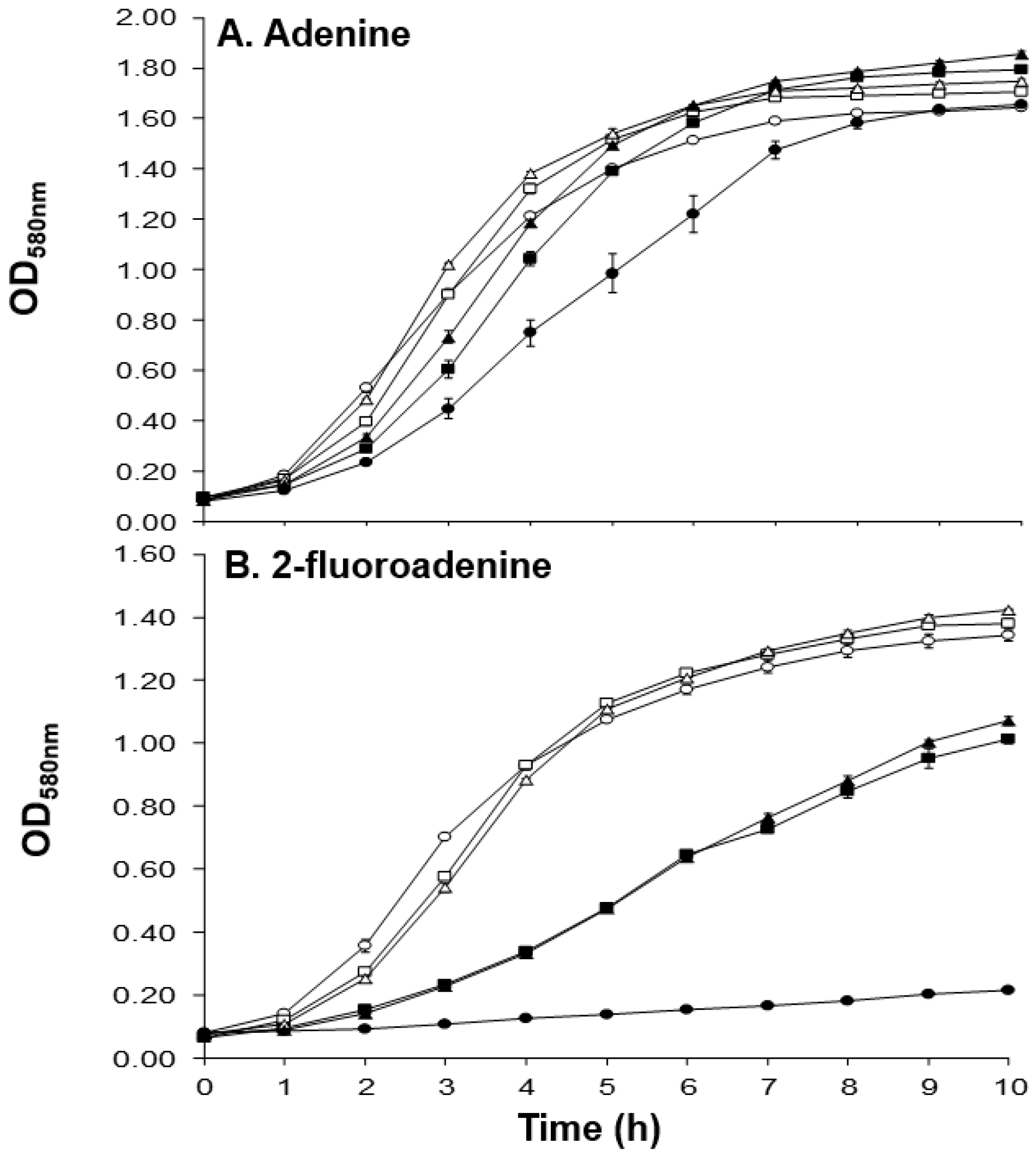
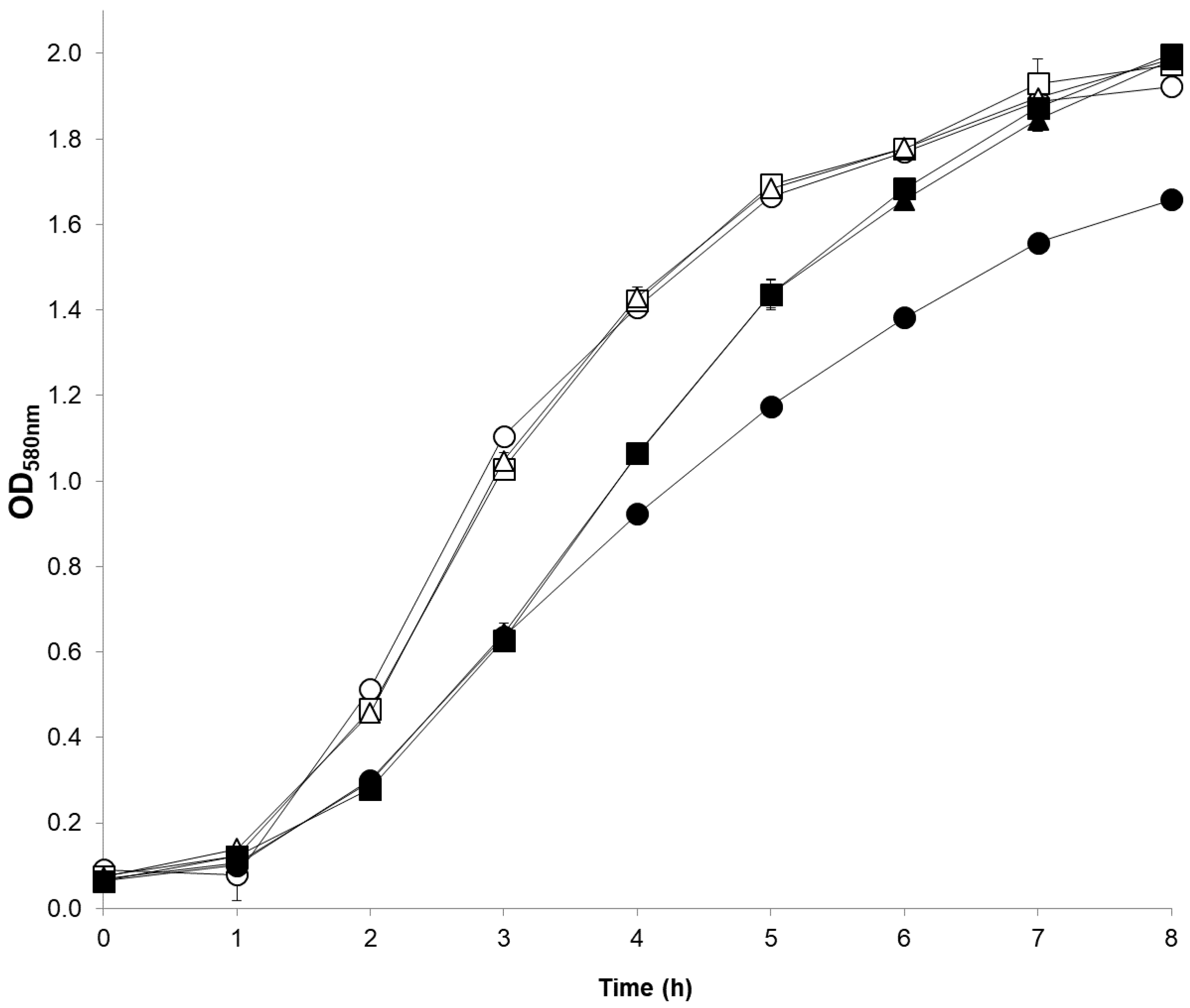
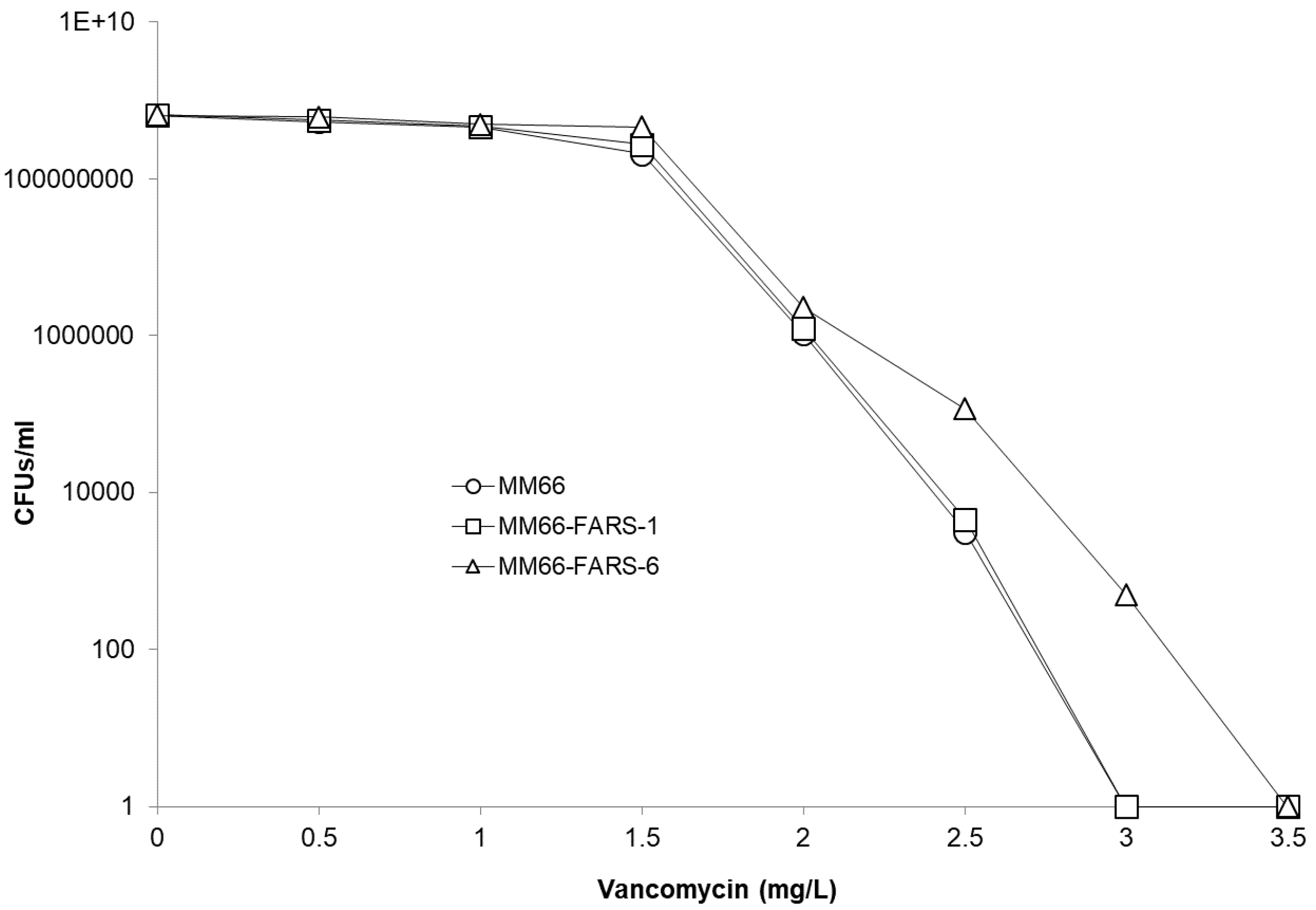
2.4. Physiological Corroboration of apt Mutation in MM66 VISA Mutants and Characterization of MM66 apt Mutants
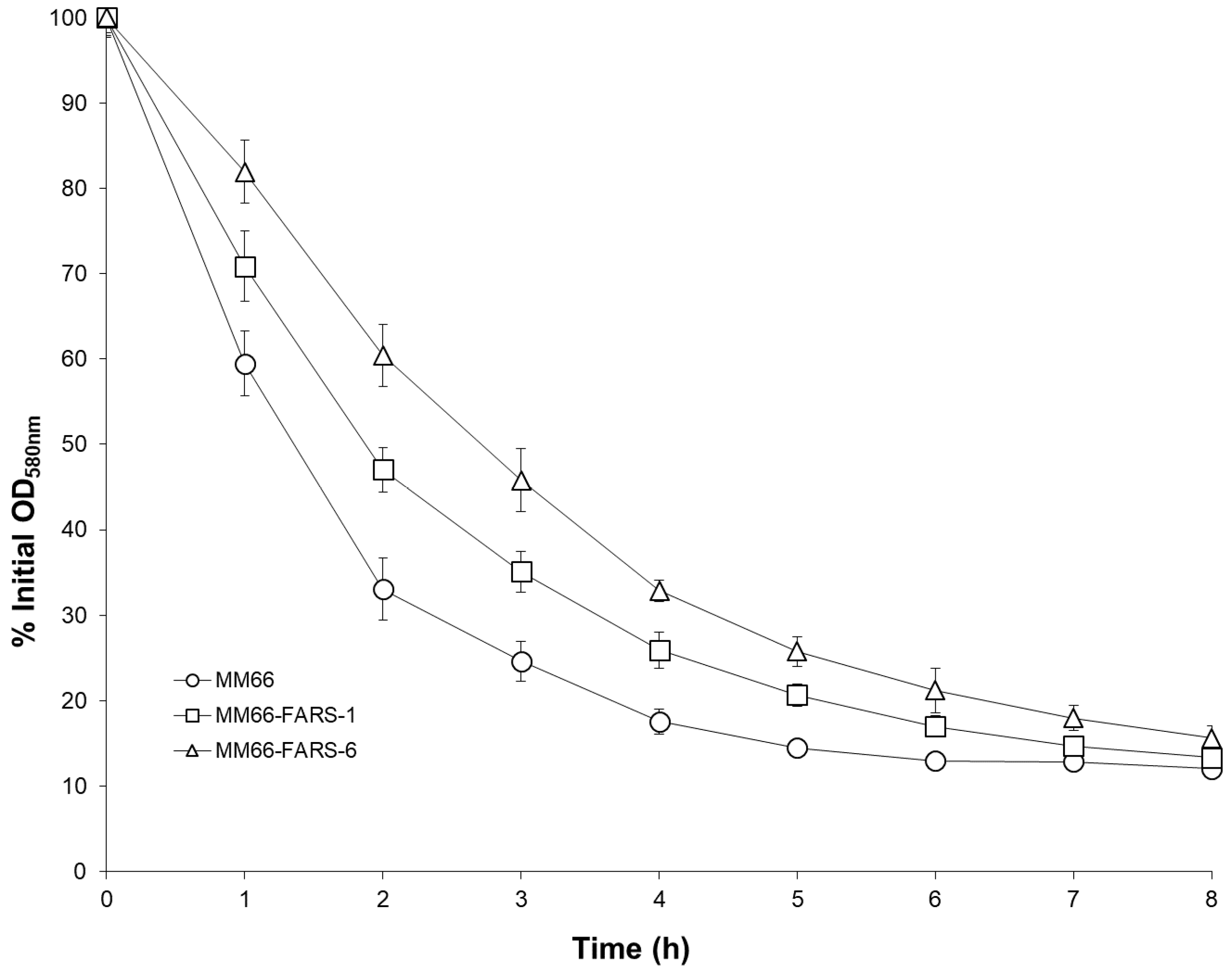
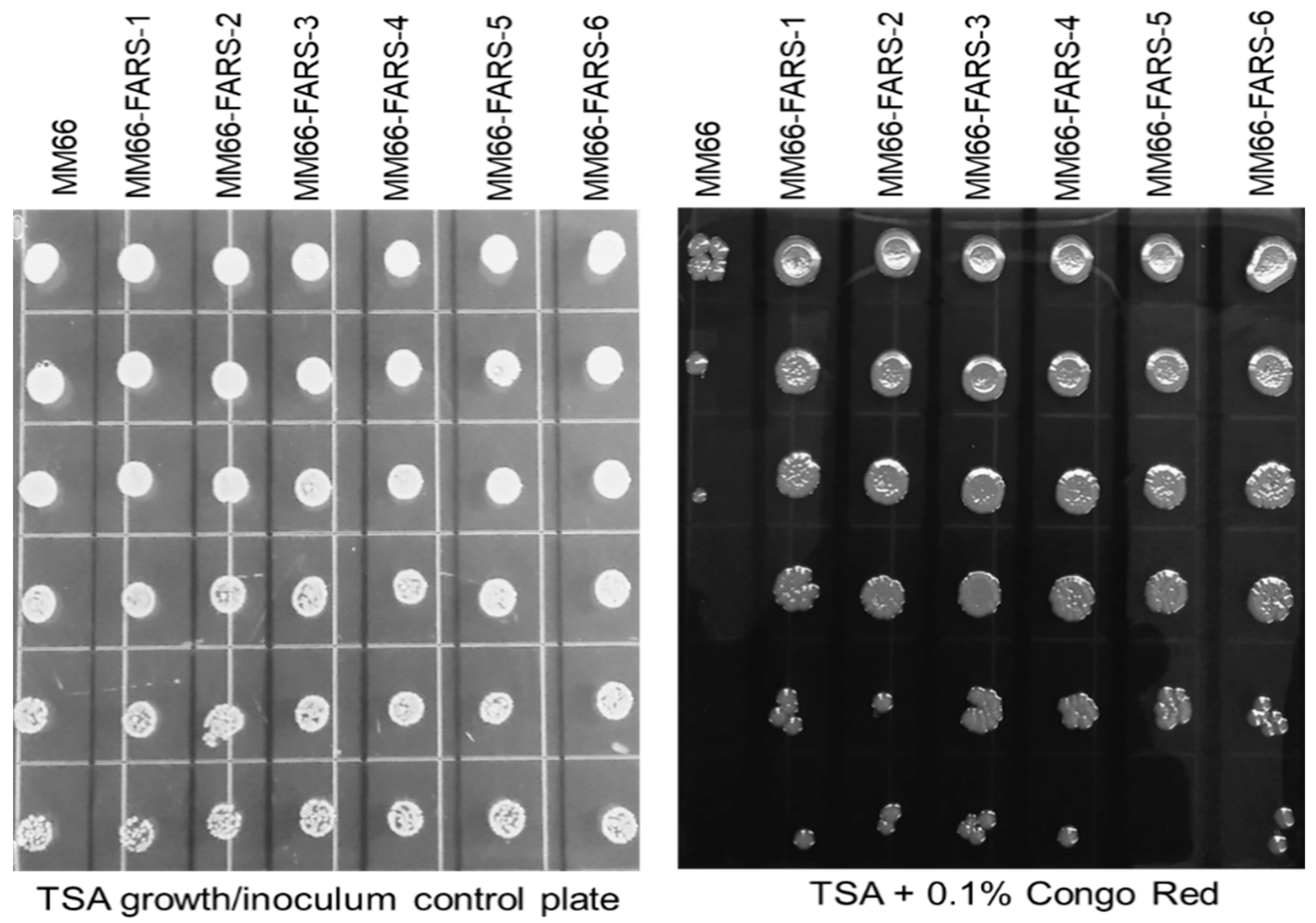
2.5. Alteration in Virulence Factor Gene Expression and Virulence Phenotypes of MM66 VISA Mutants
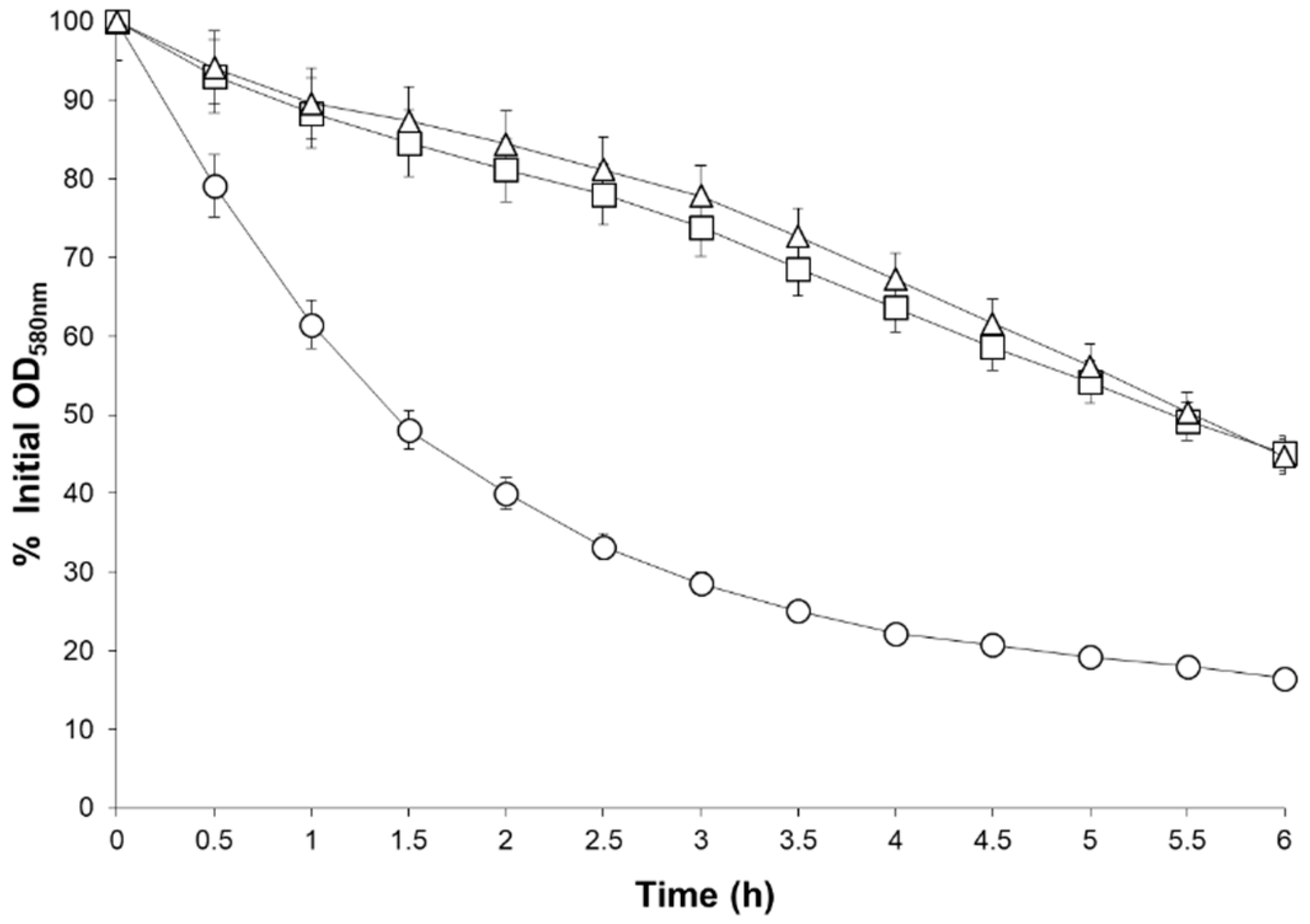
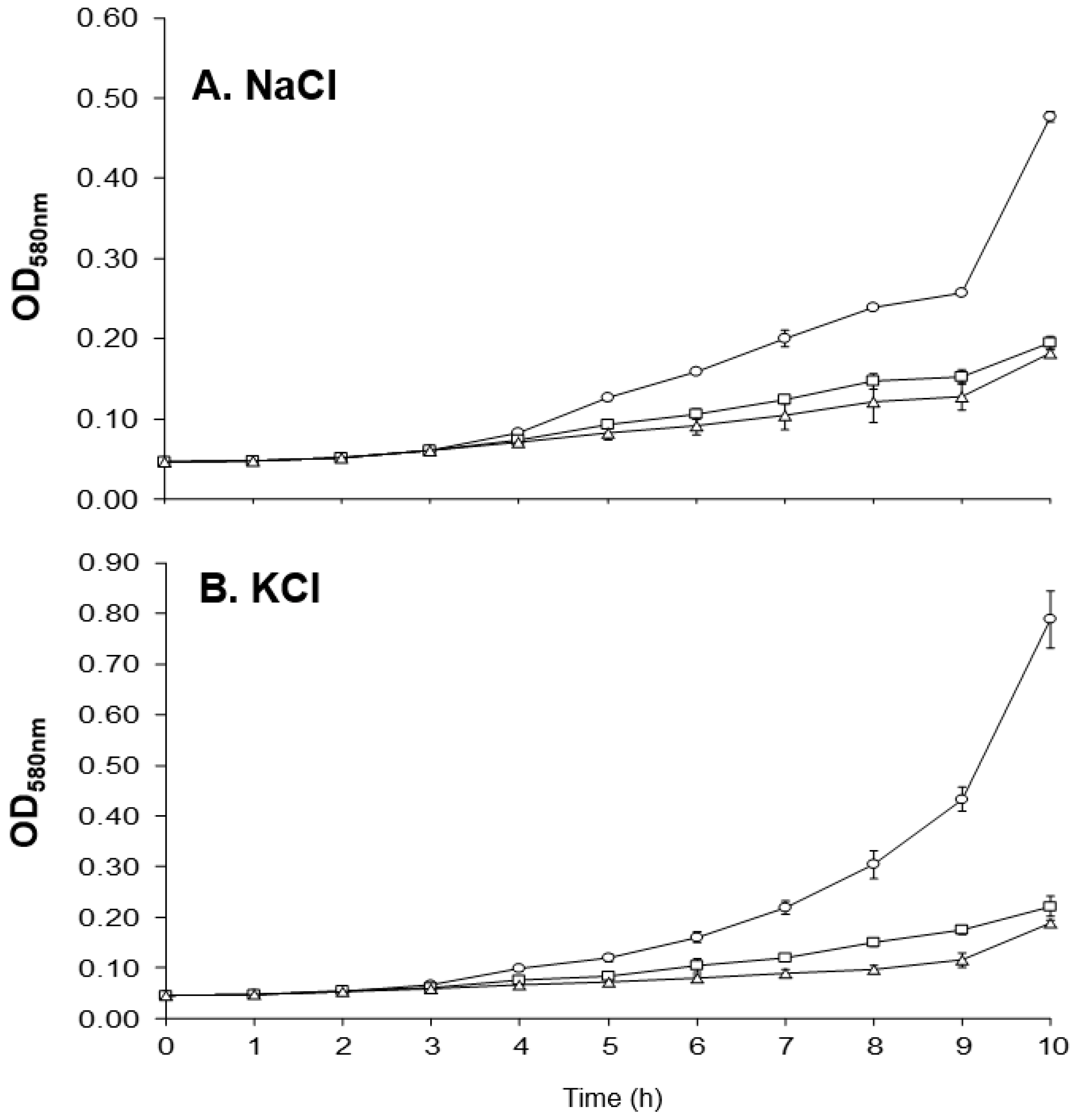
2.6. Altered Growth of MM66 VISA Mutants in the Presence of Salt
3. Conclusions
4. Materials and Methods
4.1. Bacterial Strains, Culture Conditions, and Chemicals
4.2. DNA and RNA Purification and cDNA Synthesis
4.3. Comparative Genome Sequencing
4.4. S. aureus DNA Microarray and Quantitative Real-Time PCR Analyses
4.5. Isolation and Characterization of MM66 FARS Mutants
4.6. Triton-X-100-Induced Whole Cell Autolysis, Coagulase Time and Hemolysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.; Stinear, T.P.; Grayson, M.L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: Resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef] [PubMed]
- Mongodin, E.; Finan, J.; Climo, M.W.; Rosato, A.; Gill, S.; Archer, G.L. Microarray transcription analysis of clinical Staphylococcus aureus isolates resistant to vancomycin. J. Bacteriol. 2003, 185, 4638–4643. [Google Scholar] [CrossRef]
- Fox, P.M.; Climo, M.W.; Archer, G.L. Lack of relationship between purine biosynthesis and vancomycin resistance in Staphylococcus aureus: A cautionary tale for microarray interpretation. Antimicrob. Agents Chemother. 2007, 51, 1274–1280. [Google Scholar] [CrossRef]
- Hershey, H.V.; Taylor, M.W. Nucleotide sequence and deduced amino acid sequence of Escherichia coli adenine phosphoribosyltransferase and comparison with other analogous enzymes. Gene 1986, 43, 287–293. [Google Scholar] [CrossRef]
- Hochstadt-Ozer, J.; Stadtman, E.R. The regulation of purine utilization in bacteria. I. Purification of adenine phosphoribosyltransferase from Escherichia coli K12 and control of activity by nucleotides. J. Biol. Chem. 1971, 246, 5294–5303. [Google Scholar] [CrossRef]
- Hochstadt-Ozer, J.; Stadtman, E.R. The regulation of purine utilization in bacteria. II. Adenine phosphoribosyltransferase in isolated membrane preparations and its role in transport of adenine across the membrane. J. Biol. Chem. 1971, 246, 5304–5311. [Google Scholar] [CrossRef]
- Hochstadt-Ozer, J.; Stadtman, E.R. The regulation of purine utilization in bacteria. III. The involvement of purine phosphoribosyltransferases in the uptake of adenine and other nucleic acid precursors by intact resting cells. J. Biol. Chem. 1971, 246, 5312–5320. [Google Scholar] [CrossRef]
- Hattangady, D.S.; Singh, A.K.; Muthaiyan, A.; Jayaswal, R.K.; Gustafson, J.E.; Ulanov, A.V.; Li, Z.; Wilkinson, B.J.; Pfeltz, R.F. Genomic, transcriptomic and metabolomic studies of two well-characterized, laboratory-derived vancomycin-intermediate Staphylococcus aureus strains derived from the same parent strain. Antibiotics 2015, 4, 76–112. [Google Scholar] [CrossRef] [PubMed]
- Dengler, V.; Foulston, L.; DeFrancesco, A.S.; Losick, R. An electrostatic net model for the role of extracellular DNA in biofilm formation by Staphylococcus aureus. J. Bacteriol. 2015, 197, 3779–3787. [Google Scholar] [CrossRef]
- DeFrancesco, A.S.; Masloboeva, N.; Syed, A.K.; DeLoughery, A.; Bradshaw, N.; Li, G.W.; Gilmore, M.S.; Walker, S.; Losick, R. Genome-wide screen for genes involved in eDNA release during biofilm formation by Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2017, 114, E5969–E5978. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, F.C.; Baddiley, J. A continuum of anionic charge: Structures and functions of D-alanyl-teichoic acids in gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Campbell, J.; Kim, Y.; Swoboda, J.G.; Mylonakis, E.; Walker, S.; Gilmore, M.S. Wall teichoic acid protects Staphylococcus aureus from inhibition by Congo red and other dyes. J. Antimicrob. Chemother. 2012, 67, 2143–2151. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Riordan, J.T.; Lamichhane-Khadka, R.; Winnett, D.C.; Jimenez, J.; Robinson, K.; O’Brien, F.G.; Cantore, S.A.; Gustafson, J.E. Hetero-vancomycin-intermediate methicillin-resistant Staphylococcus aureus isolate from a medical center in Las Cruces, New Mexico. J. Clin. Microbiol. 2007, 45, 1325–1329. [Google Scholar] [CrossRef]
- Jensen, K.F.; Hansen, M.R.; Jensen, K.S.; Christoffersen, S.; Poulsen, J.C.; Molgaard, A.; Kadziola, A. Adenine phosphoribosyltransferase from Sulfolobus solfataricus is an enzyme with unusual kinetic properties and a crystal structure that suggests it evolved from a 6-oxopurine phosphoribosyltransferase. Biochemistry 2015, 54, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Zaman, S.; Muthaiyan, A.; Nagarajan, V.; Elasri, M.O.; Wilkinson, B.J.; Gustafson, J.E. The fusidic acid stimulon of Staphylococcus aureus. J. Antimicrob. Chemother. 2008, 62, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Rawlings, N.D. The CHAP domain: A large family of amidases including GSP amidase and peptidoglycan hydrolases. Trends Biochem. Sci. 2003, 28, 234–237. [Google Scholar] [CrossRef]
- Dubrac, S.; Boneca, I.G.; Poupel, O.; Msadek, T. New insights into the WalK/WalR (YycG/YycF) essential signal transduction pathway reveal a major role in controlling cell wall metabolism and biofilm formation in Staphylococcus aureus. J. Bacteriol. 2007, 189, 8257–8269. [Google Scholar] [CrossRef]
- Dubrac, S.; Msadek, T. Identification of genes controlled by the essential YycG/YycF two-component system of Staphylococcus aureus. J. Bacteriol. 2004, 186, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.; Dubrac, S.; Andersen, K.K.; Noone, D.; Fert, J.; Msadek, T.; Devine, K. Genes controlled by the essential YycG/YycF two-component system of Bacillus subtilis revealed through a novel hybrid regulator approach. Mol. Microbiol. 2003, 49, 1639–1655. [Google Scholar] [CrossRef] [PubMed]
- Multari, R.A.; Cremers, D.A.; Bostian, M.L.; Dupre, J.M.; Gustafson, J.E. Proof of principle for a real-time pathogen isolation media diagnostic: The use of laser-induced breakdown spectroscopy to discriminate bacterial pathogens and antimicrobial-resistant Staphylococcus aureus strains grown on blood agar. J. Pathog. 2013, 2013, 898106. [Google Scholar] [CrossRef]
- Matyi, S.A.; Ramaraj, T.; Sundararajan, A.; Lindquist, I.E.; Devitt, N.P.; Schilkey, F.D.; Lamichhane0Khadka, R.; Hoyt, P.R.; Mudge, J.; Gustafson, J.E. Draft genomes of heterogeneous vancomycin-intermediate Staphylococcus aureus strain MM66 and MM66 Derivatives with Altered Vancomycin Resistance Levels. Genome Announc. 2014, 2. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.; Xu, B.; Chen, L.; Wu, Y.; Yang, X.; Shen, X.; Molin, S.; Danchin, A.; Jiang, H.; et al. Structure-based discovery of inhibitors of the YycG histidine kinase: New chemical leads to combat Staphylococcus epidermidis infections. BMC Microbiol. 2006, 6, 96. [Google Scholar]
- Howden, B.P.; McEvoy, C.R.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.F.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; et al. Evolution of multidrug resistance during Staphylococcus aureus infection involves mutation of the essential two component regulator WalKR. PLoS Pathog. 2011, 7, e1002359. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.; Turck, M.; Szekat, C.; Nagel, M.; Clever, I.; Bierbaum, G. Role of insertion elements and yycFG in the development of decreased susceptibility to vancomycin in Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 205–215. [Google Scholar] [CrossRef]
- Peng, H.; Hu, Q.; Shang, W.; Yuan, J.; Zhang, X.; Liu, H.; Zheng, Y.; Hu, Z.; Yang, Y.; Tan, L.; et al. WalK(S221P), a naturally occurring mutation, confers vancomycin resistance in VISA strain XN108. J. Antimicrob. Chemother. 2017, 72, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Shoji, M.; Cui, L.; Iizuka, R.; Komoto, A.; Neoh, H.M.; Watanabe, Y.; Hishinuma, T.; Hiramatsu, H. walK and clpP mutations confer reduced vancomycin susceptibility in Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3870–3881. [Google Scholar] [CrossRef] [PubMed]
- Rosey, E.L.; Oskouian, B.; Stewart, G.C. Lactose metabolism by Staphylococcus aureus: Characterization of lacABCD, the structural genes of the tagatose 6-phosphate pathway. J. Bacteriol. 1991, 173, 5992–5998. [Google Scholar] [CrossRef] [PubMed]
- Miallau, L.; Hunter, W.N.; McSweeney, S.M.; Leonard, G.A. Structures of Staphylococcus aureus D-tagatose-6-phosphate kinase implicate domain motions in specificity and mechanism. J. Biol. Chem. 2007, 282, 19948–19957. [Google Scholar] [CrossRef] [PubMed]
- Oskouian, B.; Stewart, G.C. Cloning and characterization of the repressor gene of the Staphylococcus aureus lactose operon. J. Bacteriol. 1987, 169, 5459–5465. [Google Scholar] [CrossRef] [PubMed]
- McAleese, F.; Wu, S.W.; Sieradzki, K.; Dunman, P.; Murphy, E.; Projan, S.; Tomasz, A. Overexpression of genes of the cell wall stimulon in clinical isolates of Staphylococcus aureus exhibiting vancomycin-intermediate–S. aureus-type resistance to vancomycin. J. Bacteriol. 2006, 188, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; de Lencastre, H.; Tomasz, A. Sigma-B, a putative operon encoding alternate sigma factor of Staphylococcus aureus RNA polymerase: Molecular cloning and DNA sequencing. J. Bacteriol. 1996, 178, 6036–6042. [Google Scholar] [CrossRef]
- Senn, M.M.; Giachino, P.; Homerova, D.; Steinhuber, A.; Strassner, J.; Kormanec, J.; Fluckiger, U.; Berger-Bachi, B.; Bischoff, M. Molecular analysis and organization of the sigmaB operon in Staphylococcus aureus. J. Bacteriol. 2005, 187, 8006–8019. [Google Scholar] [CrossRef] [PubMed]
- Pane-Farre, J.; Jonas, B.; Forstner, K.; Engelmann, S.; Hecker, M. The sigmaB regulon in Staphylococcus aureus and its regulation. Int. J. Med. Microbiol. 2006, 296, 237–258. [Google Scholar] [CrossRef]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bachi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus sigmaB regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef]
- Keinhorster, D.; Salzer, A.; Duque-Jaramillo, A.; George, S.E.; Marincola, G.; Lee, J.C.; Weidenmaier, C.; Wolz, C. Revisiting the regulation of the capsular polysaccharide biosynthesis gene cluster in Staphylococcus aureus. Mol. Microbiol. 2019, 112, 1083–1099. [Google Scholar] [CrossRef]
- Weigel, L.M.; Clewell, D.B.; Gill, S.R.; Clark, N.C.; McDougal, L.K.; Flannagan, S.E.; Kolonay, J.F.; Shetty, J.; Killgore, G.E.; Tenover, F.C. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef]
- Hirschhausen, N.; Schlesier, T.; Peters, G.; Heilmann, C. Characterization of the modular design of the autolysin/adhesin Aaa from Staphylococcus aureus. PLoS ONE 2012, 7, e40353. [Google Scholar] [CrossRef]
- Liang, X.; Zheng, L.; Landwehr, C.; Lunsford, D.; Holmes, D.; Ji, Y. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol. 2005, 187, 5486–5492. [Google Scholar] [CrossRef]
- Fournier, B.; Hooper, D.C. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proteolytic activity of Staphylococcus aureus. J. Bacteriol. 2000, 182, 3955–3964. [Google Scholar] [CrossRef]
- Crosby, H.A.; Tiwari, N.; Kwiecinski, J.M.; Xu, Z.; Dykstra, A.; Jenul, C.; Fuentes, E.J.; Horswill, A.R. The Staphylococcus aureus ArlRS two-component system regulates virulence factor expression through MgrA. Mol. Microbiol. 2020, 113, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Luong, T.T.; Dunman, P.M.; Murphy, E.; Projan, S.J.; Lee, C.Y. Transcription profiling of the mgrA regulon in Staphylococcus aureus. J. Bacteriol. 2006, 188, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Memmi, G.; Nair, D.R.; Cheung, A. Role of ArlRS in autolysis in methicillin-sensitive and methicillin-resistant Staphylococcus aureus strains. J. Bacteriol. 2012, 194, 759–767. [Google Scholar] [CrossRef]
- Blake, K.L.; O’Neill, A.J.; Mengin-Lecreulx, D.; Henderson, P.J.; Bostock, J.M.; Dunsmore, C.J.; Simmons, K.J.; Fishwick, C.W.G.; Leeds, J.A.; Chopra, I. The nature of Staphylococcus aureus MurA and MurZ and approaches for detection of peptidoglycan biosynthesis inhibitors. Mol. Microbiol. 2009, 72, 335–343. [Google Scholar] [CrossRef]
- Stapleton, M.R.; Horsburgh, M.J.; Hayhurst, E.J.; Wright, L.; Jonsson, I.M.; Tarkowski, A.; Kokai-Kun, J.F.; Mond, J.J.; Foster, S.J. Characterization of IsaA and SceD, two putative lytic transglycosylases of Staphylococcus aureus. J. Bacteriol. 2007, 189, 7316–7325. [Google Scholar] [CrossRef]
- Saxild, H.H.; Nygaard, P. Genetic and physiological characterization of Bacillus subtilis mutants resistant to purine analogs. J. Bacteriol. 1987, 169, 2977–2983. [Google Scholar] [CrossRef]
- de Repentigny, J.; Grimard, S.; Turgeon, P.; Sonea, S. Inhibition by adenine of Staphylococcus aureus growth in a nutrient medium free from guanine, guanosine, or hypoxanthine. J. Bacteriol. 1966, 91, 2099–2100. [Google Scholar] [CrossRef]
- Phonimdaeng, P.; O’Reilly, M.; O’Toole, P.W.; Foster, T.J. Molecular cloning and expression of the coagulase gene of Staphylococcus aureus 8325–4. J. Gen. Microbiol. 1988, 134, 75–83. [Google Scholar] [CrossRef]
- Cooney, J.; Kienle, Z.; Foster, T.J.; O’Toole, P.W. The gamma-hemolysin locus of Staphylococcus aureus comprises three linked genes, two of which are identical to the genes for the F and S components of leukocidin. Infect. Immun. 1993, 61, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Pattee, P.A. Chromosomal map location of the alpha-hemolysin structural gene in Staphylococcus aureus NCTC 8325. Infect. Immun. 1986, 54, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Wilkinson, B.J. Staphylococcus aureus osmoregulation: Roles for choline, glycine betaine, proline, and taurine. J. Bacteriol. 1992, 174, 2711–2716. [Google Scholar] [CrossRef]
- Roesser, M.; Muller, V. Osmoadaptation in bacteria and archaea: Common principles and differences. Environ. Microbiol. 2001, 3, 743–754. [Google Scholar] [CrossRef]
- Kempf, B.; Bremer, E. OpuA, an osmotically regulated binding protein-dependent transport system for the osmoprotectant glycine betaine in Bacillus subtilis. J. Biol. Chem. 1995, 270, 16701–16713. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Peleg, A.Y.; Stinear, T.P. The evolution of vancomycin intermediate Staphylococcus aureus (VISA) and heterogenous-VISA. Infect. Genet. Evol. 2014, 21, 575–582. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Spina, D.; Purrello, S.; Campanile, F.; Di Pietro, C.; Purrello, M.; Stefani, S. Modulating activity of vancomycin and daptomycin on the expression of autolysis cell-wall turnover and membrane charge genes in hVISA and VISA strains. PLoS ONE 2012, 7, e29573. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Lecca, D.; Abbracchio, M.P.; Ceruti, S. Pathophysiological role of purines and pyrimidines in neurodevelopment: Unveiling new pharmacological approaches to congenital brain diseases. Front. Pharmacol. 2017, 8, 941. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Koebmann, B.J.; Jensen, P.R.; Nilsson, D. Increasing acidification of nonreplicating Lactococcus lactis deltathyA mutants by incorporating ATPase activity. Appl. Environ. Microbiol. 2002, 68, 5249–5257. [Google Scholar] [CrossRef]
- Sieradzki, K.; Tomasz, A. Suppression of glycopeptide resistance in a highly teicoplanin-resistant mutant of Staphylococcus aureus by transposon inactivation of genes involved in cell wall synthesis. Microb. Drug Resist. 1998, 4, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Schmidt, J.L.; Jayaswal, R.K.; Wilkinson, B.J. Impact of sigB mutation on Staphylococcus aureus oxacillin and vancomycin resistance varies with parental background and method of assessment. Int. J. Antimicrob. Agents 2003, 21, 256–261. [Google Scholar] [CrossRef]
- Marquis, R.E. Salt-induced contraction of bacterial cell walls. J. Bacteriol. 1968, 95, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Vijaranakul, U.; Nadakavukaren, M.J.; de Jonge, B.L.; Wilkinson, B.J.; Jayaswal, R.K. Increased cell size and shortened peptidoglycan interpeptide bridge of NaCl-stressed Staphylococcus aureus and their reversal by glycine betaine. J. Bacteriol. 1995, 177, 5116–5121. [Google Scholar] [CrossRef]
- Brown, D.F.; Edwards, D.I.; Hawkey, P.M.; Morrison, D.; Ridgway, G.L.; Towner, K.J.; Wren, Joint Working Party of the British Society for Antimicrobial Chemotherapy; Hospital Infection Society; Infection Control Nurses Association. Guidelines for the laboratory diagnosis and susceptibility testing of methicillin-resistant Staphylococcus aureus (MRSA). J. Antimicrob. Chemother. 2005, 56, 1000–1018. [Google Scholar] [CrossRef]
- Madiraju, M.V.; Brunner, D.P.; Wilkinson, B.J. Effects of temperature, NaCl, and methicillin on penicillin-binding proteins, growth, peptidoglycan synthesis, and autolysis in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 1987, 31, 1727–1733. [Google Scholar] [CrossRef]
- Riordan, J.T.; O’Leary, J.O.; Gustafson, J.E. Contributions of sigB and sarA to distinct multiple antimicrobial resistance mechanisms of Staphylococcus aureus. Int. J. Antimicrob. Agents 2006, 28, 54–61. [Google Scholar] [CrossRef]
- Riordan, J.T.; Muthaiyan, A.; Van Voorhies, W.; Price, C.T.; Graham, J.E.; Wilkinson, B.J.; Gustafson, J.E. Response of Staphylococcus aureus to salicylate challenge. J. Bacteriol. 2007, 189, 220–227. [Google Scholar] [CrossRef]
- Albert, T.J.; Dailidiene, D.; Dailide, G.; Norton, J.E.; Kalia, A.; Richmond, T.A.; Molla, M.; Singh, J.; Green, R.D.; Berg, D.E. Mutation discovery in bacterial genomes: Metronidazole resistance in Helicobacter pylori. Nat. Methods 2005, 2, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Price, C.T.; O’Brien, F.G.; Shelton, B.P.; Warmington, J.R.; Grubb, W.B.; Gustafson, J.E. Effects of salicylate and related compounds on fusidic acid MICs in Staphylococcus aureus. J. Antimicrob. Chemother. 1999, 44, 57–64. [Google Scholar] [CrossRef]
- O’Brien, F.G.; Botterill, C.I.; Endersby, T.G.; Lim, R.L.; Grubb, W.B.; Gustafson, J.E. Heterogeneous expression of fusidic acid resistance in Staphylococcus aureus with plasmid or chromosomally encoded fusidic acid resistance genes. Pathology 1998, 30, 299–303. [Google Scholar] [CrossRef]
- Gustafson, J.E.; Berger-Bachi, B.; Strassle, A.; Wilkinson, B.J. Autolysis of methicillin-resistant and -susceptible Staphylococcus aureus. Antimicrob. Agents Chemother. 1992, 36, 566–572. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | SACOL Loci a | Gene | Function | SNP Position a | Amino Acid Change | |
|---|---|---|---|---|---|---|
| MM66-3 | ||||||
| Intergenic | ||||||
| SACOL1918 | IS1181 transposase | T1974334→A1974334 | ||||
| SACOL1919 | Fur family transcriptional regulator | |||||
| SACOL2608 | Hypothetical protein | T2667188 → C2667188 | ||||
| SACOL2609 | Hypothetical protein | |||||
| Intragenic | ||||||
| SACOL1690 | apt | Adenine phosphoribosyltransferase | C1721075 → T1721075 | A57 → V57 | ||
| SACOL1576 | ssaA6 | CHAP domain containing protein | A1610400 → C1610400 | H297 → G297 | ||
| T1610401 → C1610401 | ||||||
| G1610402 → C1610402 | ||||||
| SACOL1836 | Hypothetical protein | T1891424 → C1891424 | I7 →V7 | |||
| SACOL2639 | cysJ | Sulfite reductase flavoprotein subunit | G2699218 → A2699218 | P458 →S458 |
| SACOL Locus a | Gene | Function | Fold Change in Gene Expression | |
|---|---|---|---|---|
| MM66-3 (RT-qPCR) | MM66-4 (RT-qPCR) | |||
| Upregulated genes | ||||
| SACOL0136 | cap5A | Capsular polysaccharide biosynthesis | 2.9 | 4 |
| SACOL0137 | cap5B | Capsular polysaccharide biosynthesis | ND | 2.7 |
| SACOL0660 | adhP | Alcohol dehydrogenase | 2.1 (1.2) | 2.3 (3.9) |
| SACOL0781 | opuAC | Osmoprotectant ABC transporter component | ND | 2.3 |
| SACOL0856 | clfA | Clumping factor A | 2.1 | ND |
| SACOL1078 | purL | Phosphoribosylformylglycinamidine synthase II | 2.2 | 2.2 |
| SACOL1079 | purF | Amidophosphoribosyltransferase | 4.2 | 3.7 |
| SACOL1080 | purM | Phosphoribosylaminoimidazole synthetase | 4.5 (4.5) | 4.0 (3.3) |
| SACOL1081 | purN | Phosphoribosylglycinamide formyltransferase | 3.9 | 3.8 |
| SACOL1082 | purH | Phosphoribosylaminoimidazolecarboxamide | 2.1 | 2.1 |
| SACOL1083 | purD | Phosphoribosylamine-glycine ligase | 2.9 | 2.8 |
| SACOL1215 | pyrAB | Carbamoyl-phosphate synthase large subunit | 3.1 | 2.5 |
| SACOL1216 | pyrF | Orotidine 5-phosphate decarboxylase | 2.3 | 2.3 |
| SACOL1217 | pyrE | Orotate phosphoribosyltransferase | 3.1 | 2.8 |
| SACOL1328 | glnR | Glutamine synthetase repressor | 2 | 3.6 |
| SACOL1450 | arlS | Sensor histidine kinase | 2 | 2.1 |
| SACOL1451 | arlR | DNA-binding response regulator | 2.2 | 3.2 |
| SACOL2054 | sigB | RNA polymerase sigma factor sigB | ND (2.0) | 2.3 (1.4) |
| SACOL2055 | rsbW | Anti-sigma B factor | ND | 2.9 |
| SACOL2056 | rsbV | Anti-anti-sigma factor | ND | 2.2 |
| SACOL2088 | sceD | sceD autolysin | 2.7 | 3.7 |
| SACOL2092 | murA | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | ND | 2.7 |
| SACOL2176 | opuD1 | BCCT family osmoprotectant transporter | 2.2 | 2.3 |
| SACOL2181 | lacE | PTS system, lactose-specific IIBC components | 12.7 | 6.1 |
| SACOL2182 | lacF | PTS system, lactose-specific IIA component | 10.8 (4.0) | 9.0 (3.2) |
| SACOL2187 | lacA1 | Hypothetical protein | 14 | 9.4 |
| SACOL2671 | secA | secA preprotein translocase subunit | 2.7 | 3.1 |
| Downregulated genes | ||||
| SACOL0209 | coa | Staphylocoagulase precursor | −3.0 (−1.1) | −3.3 (−2.1) |
| SACOL0478 | ssl11 | Superantigen-like protein | ND | −2.1 |
| SACOL0507 | aaa | Multifunctional autolysin | ND | −3.2 |
| SACOL0723 | aaa1 | aaa homolog (38 % identity) | ND | −2.9 |
| SACOL1062 | atl | Bifunctional autolysin | ND | −2 |
| SACOL1164 | Fibrinogen binding-related protein | −3.9 | −3.9 | |
| SACOL1169 | Fibrinogen-binding protein precursor-related protein | −2.5 | −3.4 | |
| SACOL1173 | hly | Alpha-hemolysin precursor | −2.4 (−1.9) | −5.0 (−2.6) |
| SACOL2291 | ssaA1 | Secretory antigen precursor ssaA | ND | −2.2 |
| SACOL2295 | ssaA3 | ssaA homolog | ND | −2.2 |
| SACOL2418 | sbi | IgG-binding protein | −2.3 (−1.2) | −3.3 (−3.0) |
| SACOL2421 | hlgC | Gamma-hemolysin, component C | ND | −2.5 |
| SACOL2422 | hlgB | Gamma hemolysin, component B | ND | −2.2 |
| SACOL2581 | ssaA2 | ssaA homolog | ND | −3.1 |
| SACOL2627 | betA | Choline dehydrogenase | ND | −7.4 |
| SACOL2628 | betB | Glycine betaine aldehyde dehydrogenase | ND (−2.2) | −9.2 (−7.0) |
| SACOL2632 | opuD2 | BCCT family osmoprotectant transporter | ND | −4.6 |
| Strain | Parent Strain | 2-FA Selection Conc. (mM) | 2-FA MIC (mM) | References |
|---|---|---|---|---|
| MM66 | 5 | [15] | ||
| MM66-FA-1 | MM66 | 5 | >7 | This study |
| MM66-FA-2 | MM66 | 5 | >7 | This study |
| MM66-FA-3 | MM66 | 5 | >7 | This study |
| MM66-FA-4 | MM66 | 5 | >7 | This study |
| MM66-FA-5 | MM66 | 5 | >7 | This study |
| MM66-FA-6 | MM66 | 5 | >7 | This study |
| 2-FARS Mutant(s) | apt Mutation * | Effect of Mutation on Apt |
|---|---|---|
| MM66-FA-1 | Contiguous 45 bp deletion between A1720937—A1720891 | M103 →I103 and H104—D118 (HKDAIKPGQRVLITD) internal deletion |
| MM66-FA-2 | Contiguous 45 bp deletion between A1720937—A1720891 | M103 →I103 and H104—D118 (HKDAIKPGQRVLITD) internal deletion |
| MM66-FA-3 | C insertion after GCACC1721018 | Frameshift after P76 |
| MM66-FA-4 | Noncontiguous 15 bp deletion between C1721039—A1721020 | M70—F74 (MGIGF) internal deletion |
| MM66-FA-5 | ATGGG insertion after TGGGG1701031 | Frameshift after G71 |
| MM66-FA-6 | T1720975 →A1720975 | Y90 →stop codon (TAA) |
| Strain | Vancomycin Gradients | |
|---|---|---|
| 0–2 mg/L | 0–2.5 mg/L | |
| MM66 | 26 ± 2.00 | 17 ± 1.00 |
| MM66-FA-1 | 47 ± 0.58 * | 33 ± 2.52 * |
| MM66-FA-2 | 48 ± 3.05 * | 33 ± 2.65 * |
| MM66-FA-3 | 49 ± 3.06 * | 36 ± 1.00 * |
| MM66-FA-4 | 52 ± 0.58 * | 37 ± 0.00 * |
| MM66-FA-5 | 55 ± 2.52 * | 38 ± 1.00 * |
| MM66-FA-6 | 53 ± 1.53 * | 36 ± 0.58 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamichhane-Khadka, R.; Dulal, S.; Cuaron, J.A.; Pfeltz, R.; Gupta, S.K.; Wilkinson, B.J.; Gustafson, J.E. Apt (Adenine Phosphoribosyltransferase) Mutation in Laboratory-Selected Vancomycin-Intermediate Staphylococcus aureus. Antibiotics 2021, 10, 583. https://doi.org/10.3390/antibiotics10050583
Lamichhane-Khadka R, Dulal S, Cuaron JA, Pfeltz R, Gupta SK, Wilkinson BJ, Gustafson JE. Apt (Adenine Phosphoribosyltransferase) Mutation in Laboratory-Selected Vancomycin-Intermediate Staphylococcus aureus. Antibiotics. 2021; 10(5):583. https://doi.org/10.3390/antibiotics10050583
Chicago/Turabian StyleLamichhane-Khadka, Reena, Santosh Dulal, Jesus A. Cuaron, Richard Pfeltz, Sushim Kumar Gupta, Brian J. Wilkinson, and John E. Gustafson. 2021. "Apt (Adenine Phosphoribosyltransferase) Mutation in Laboratory-Selected Vancomycin-Intermediate Staphylococcus aureus" Antibiotics 10, no. 5: 583. https://doi.org/10.3390/antibiotics10050583
APA StyleLamichhane-Khadka, R., Dulal, S., Cuaron, J. A., Pfeltz, R., Gupta, S. K., Wilkinson, B. J., & Gustafson, J. E. (2021). Apt (Adenine Phosphoribosyltransferase) Mutation in Laboratory-Selected Vancomycin-Intermediate Staphylococcus aureus. Antibiotics, 10(5), 583. https://doi.org/10.3390/antibiotics10050583