
Essential Oils of Aromatic Plant Species from the Atlantic Rainforest Exhibit Extensive Chemical Diversity and Antimicrobial Activity

 , and
, and
Abstract
:1. Introduction
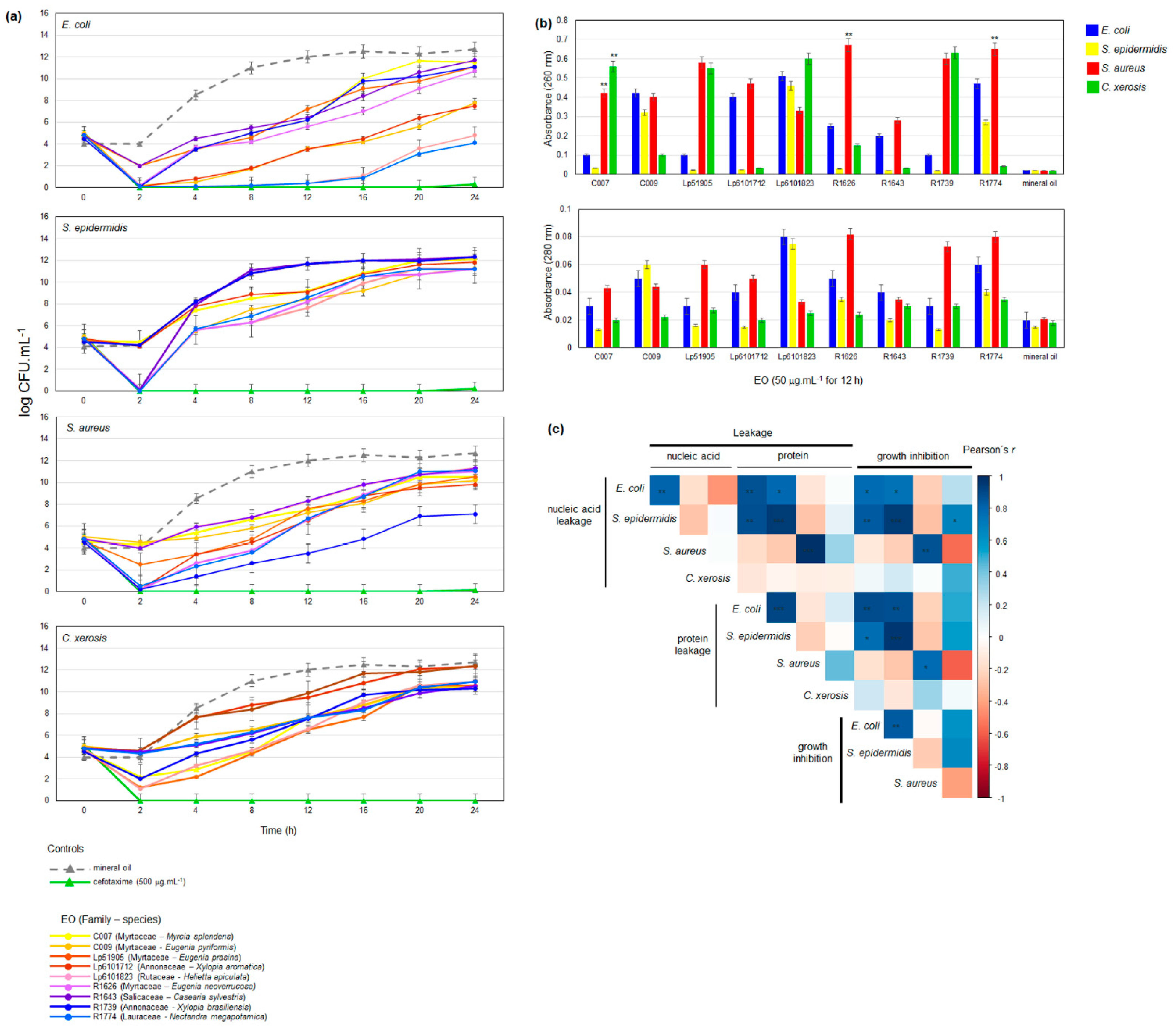
2. Results
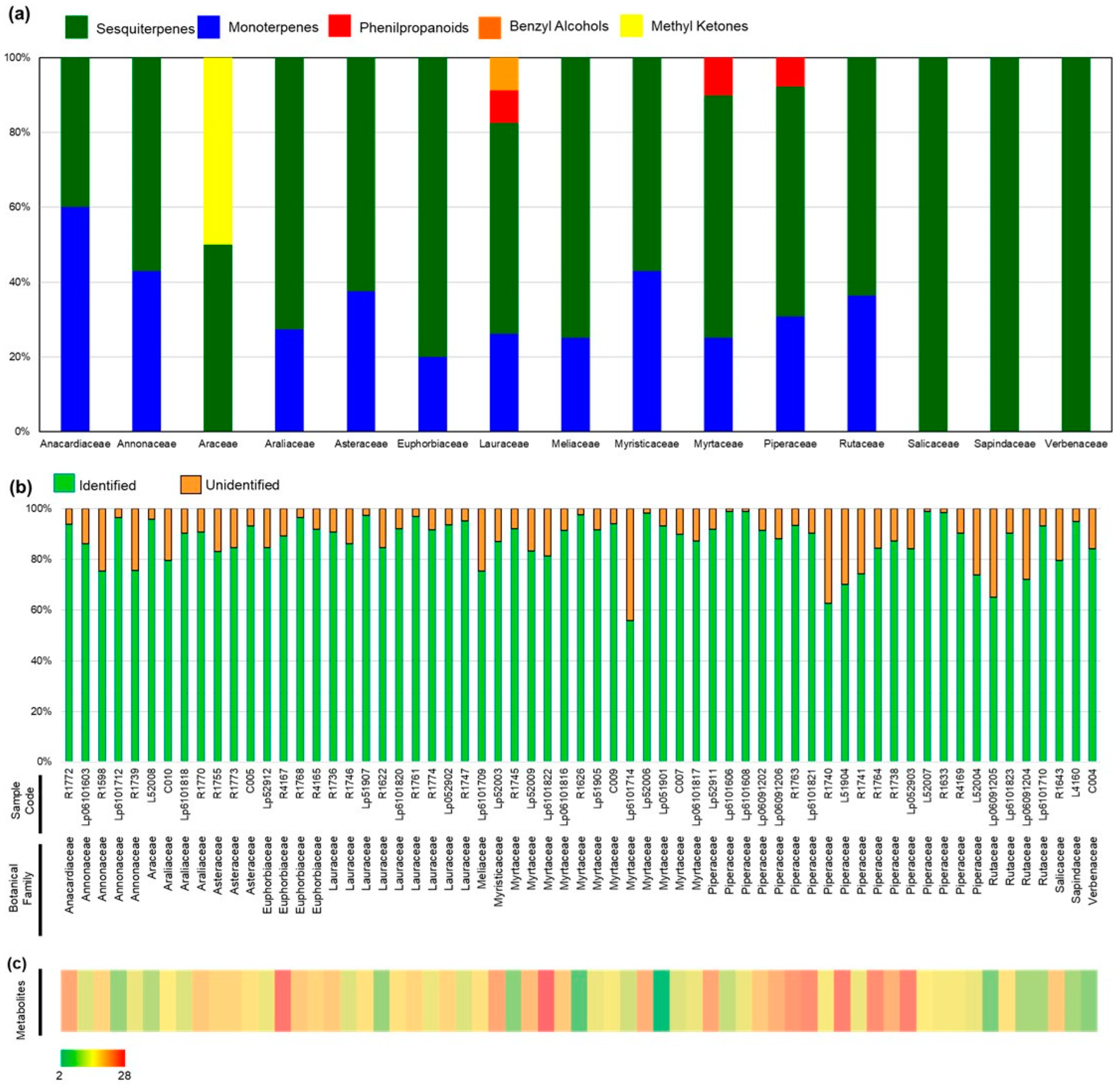
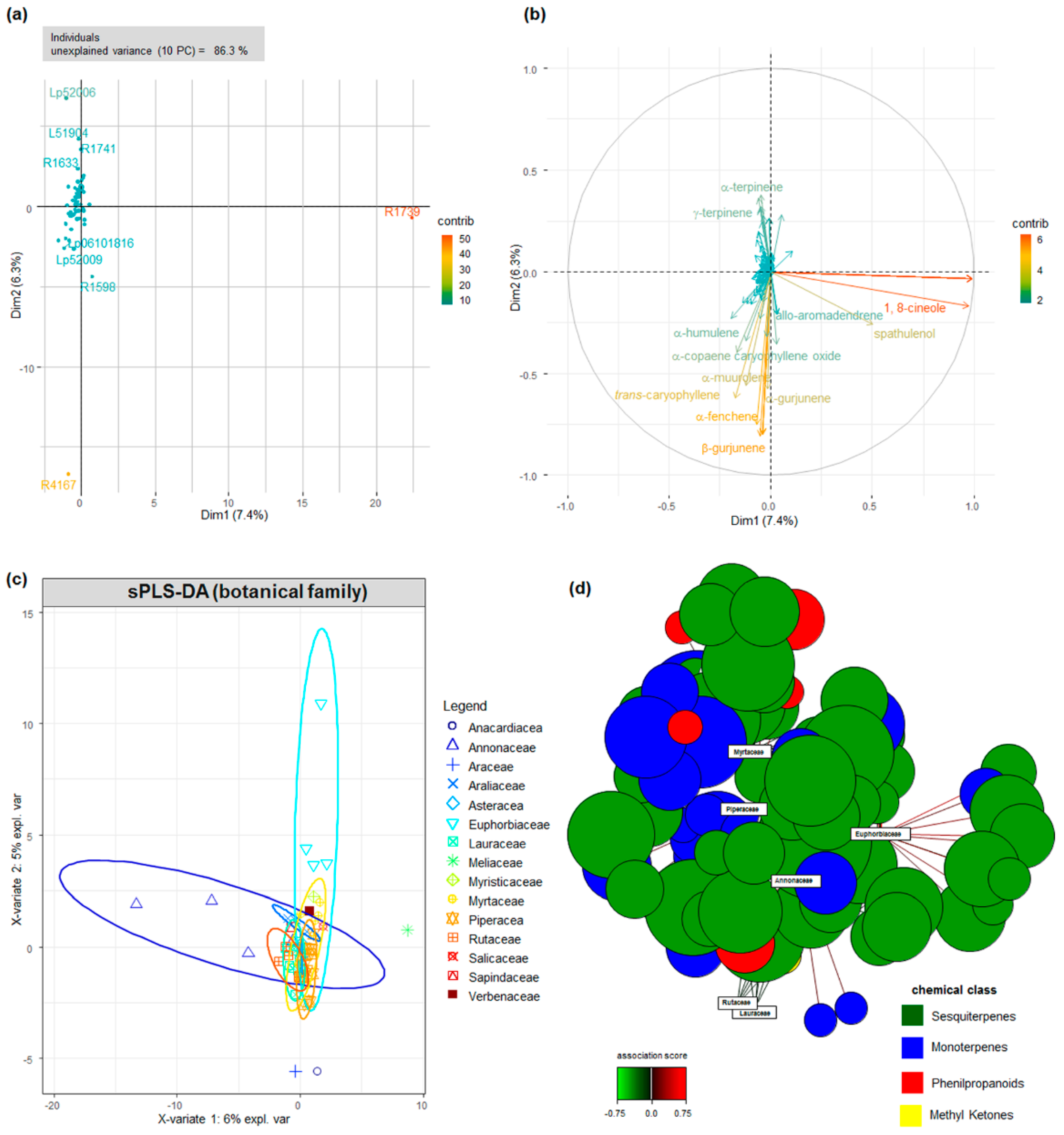
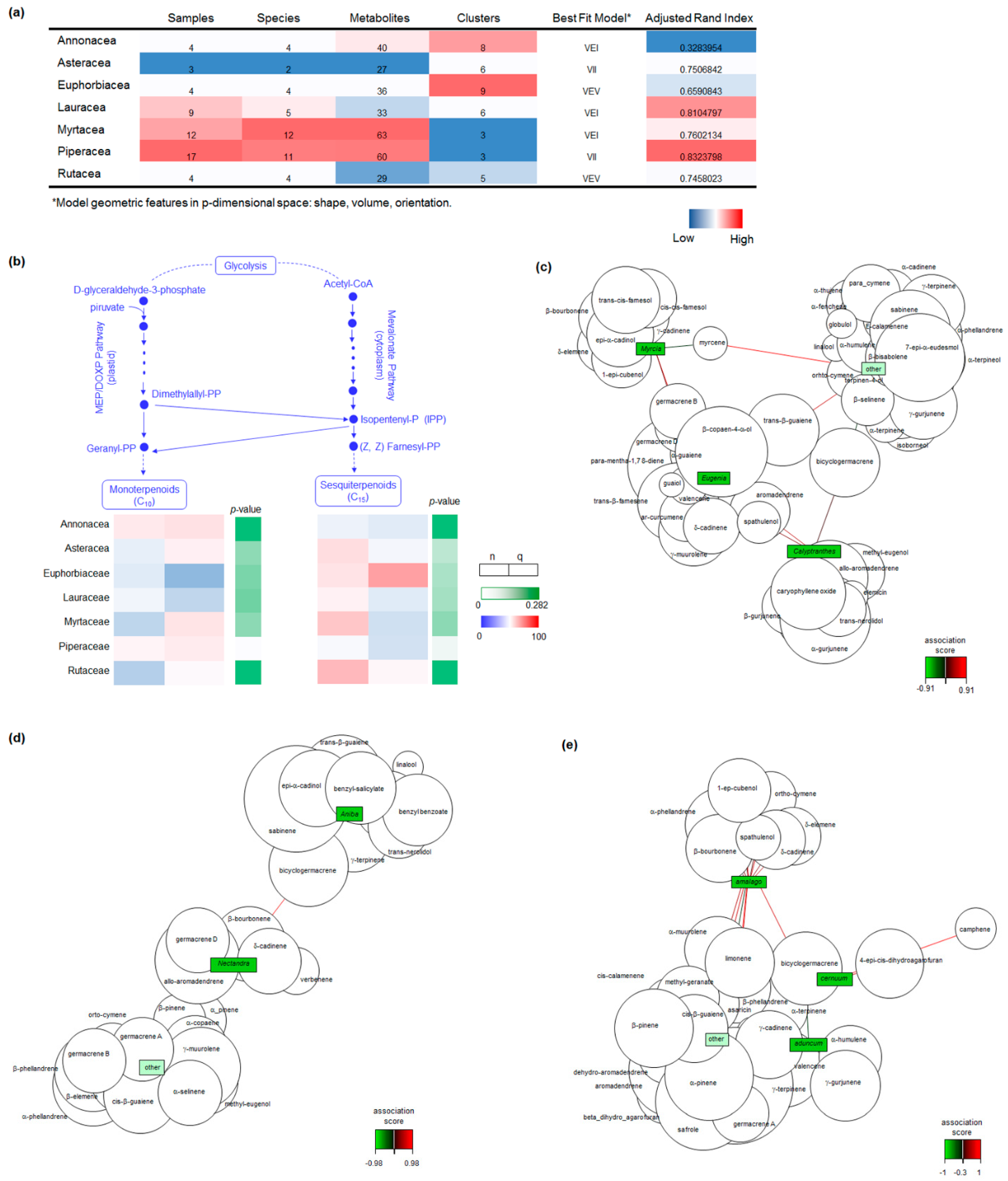
2.1. Botanical and Chemical Characterization
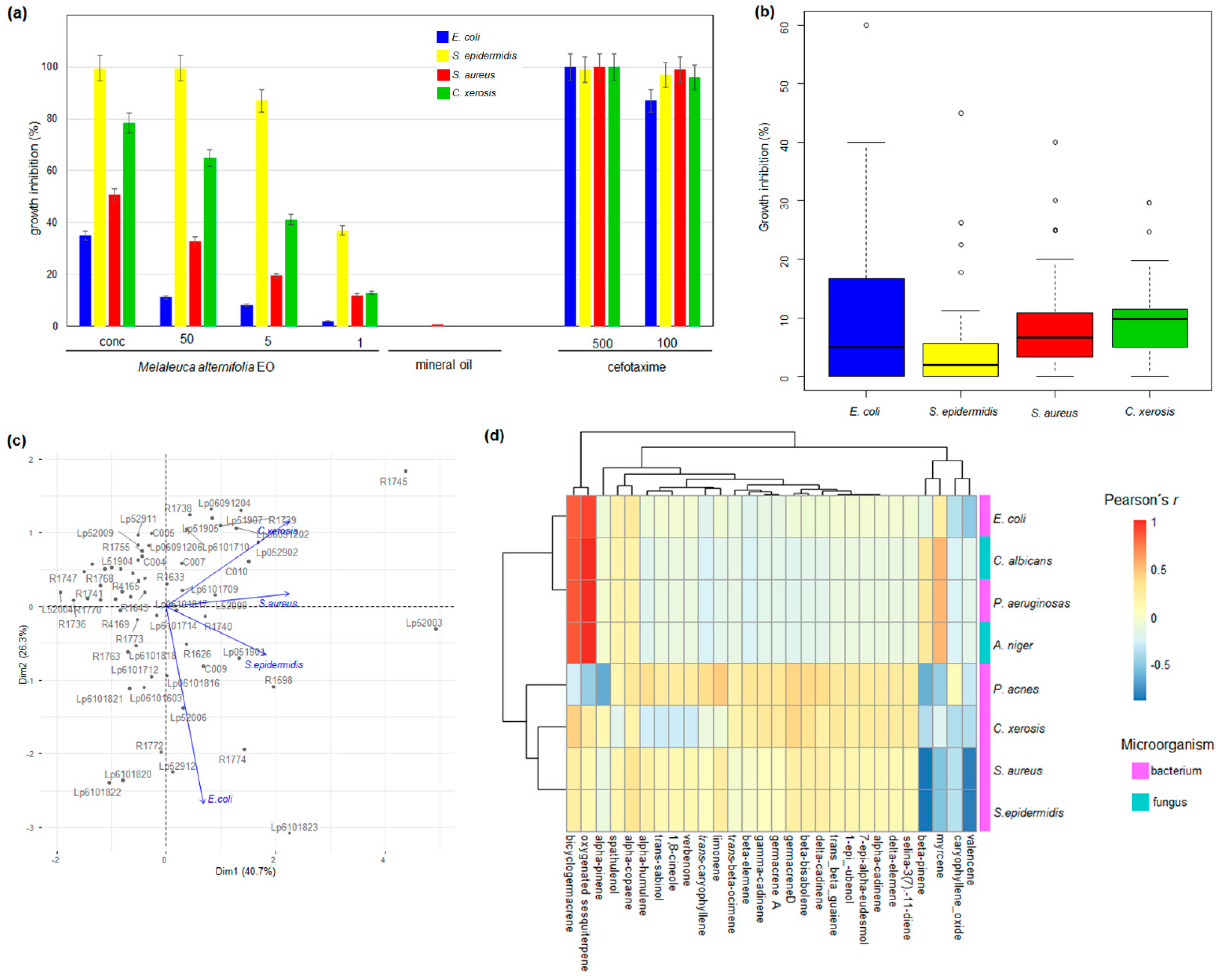
2.2. Chemical Composition and Antimicrobial Activity
3. Discussion
4. Materials and Methods
4.1. Biological Samples Collection and Environmental Data
4.2. Essential Oil Extraction
4.3. Chemical Characterization and Quantification of Essential Oils
4.4. Microbial Strains
4.5. Estimation of EO Effective Concentration for Antimicrobial Activity
4.6. Antimicrobial Activity Analyses
4.7. Mode of Action Investigations
4.8. EO Metabolite Physicochemical and Pharmacokinetic Properties
4.9. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huemer, M.; Shambat, S.M.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence—Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Pormohammad, A.; Hansen, D.; Turner, R.J. Antibacterial, Antibiofilm, and Antioxidant Activity of 15 Different Plant-Based Natural Compounds in Comparison with Ciprofloxacin and Gentamicin. Antibiotics 2022, 11, 1099. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M. The Evolution of Plant Secretory Structures and Emergence of Terpenoid Chemical Diversity. Annu. Rev. Plant Biol. 2015, 66, 139–159. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Trifan, A.; Luca, S.V.; Greige-Gerges, H.; Miron, A.; Gille, E.; Aprotosoaie, A.C. Recent advances in tackling microbial multidrug resistance with essential oils: Combinatorial and nano-based strategies. Crit. Rev. Microbiol. 2020, 46, 338–357. [Google Scholar] [CrossRef]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crops Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential oils from aromatic herbs as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef]
- Máthé, Á.; de Sales Silva, J.C. Introduction to medicinal and aromatic plants in Brazil. In Medicinal and Aromatic Plants of South America. Medicinal and Aromatic Plants of the World; Albuquerque, U., Patil, U., Máthé, Á., Eds.; Springer: Dordrecht, The Netherlands, 2018; Volume 5, pp. 47–69. [Google Scholar] [CrossRef]
- Massad, T.J.; Richards, L.A.; Philbin, C.; Yamaguchi, L.F.; Kato, M.J.; Jeffrey, C.S.; Oliveira, C., Jr.; Ochsenrider, K.; de Moraes, M.M.; Tepe, E.J.; et al. The chemical ecology of tropical forest diversity: Environmental variation, chemical similarity, herbivory, and richness. Ecology 2022, 20, e3762. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial plant compounds, extracts and essential oils: An updated review on their effects and putative mechanisms of action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef]
- Mint Evolutionary Genomics Consortium. Phylogenomic Mining of the Mints Reveals Multiple Mechanisms Contributing to the Evolution of Chemical Diversity in Lamiaceae. Mol. Plant 2018, 11, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Borges, R.S.; Ortiz, B.L.S.; Pereira, A.C.M.; Keita, H.; Carvalho, J.C.T. Rosmarinus officinalis essential oil: A review of its phytochemistry, anti-inflammatory activity, and mechanisms of action involved. J. Ethnopharmacol. 2019, 229, 29–45. [Google Scholar] [CrossRef]
- Stierlin, É.; Nicolè, F.; Costes, T.; Fernandez, X.; Michel, T. Metabolomic study of volatile compounds emitted by lavender grown under open-field conditions: A potential approach to investigate the yellow decline disease. Metabolomics 2020, 16, 31. [Google Scholar] [CrossRef]
- Alolga, R.N.; León, M.A.S.C.C.; Osei-Adjei, G.; Onoja, V. GC-MS-based metabolomics, antibacterial and anti-inflammatory investigations to characterize the quality of essential oil obtained from dried Xylopia aethiopica fruits from Ghana and Nigeria. J. Pharm. Pharmacol. 2019, 71, 1544–1552. [Google Scholar] [CrossRef]
- Perigo, C.V.; Torres, R.B.; Bernacci, L.C.; Guimarães, E.F.; Haber, L.L.; Facanali, R.; Vieira, M.A.; Quecini, V.; Marques, M.O.M. The chemical composition and antibacterial activity of eleven Piper species from distinct rainforest areas in Southeastern Brazil. Ind. Crops Prod. 2016, 94, 528–539. [Google Scholar] [CrossRef]
- Marques, M.O.; Perigo, C.V.; Haber, L.L.; Vieira, M.A.; Facanali, R.; Torres, R.B.; Bernacci, L.C.; Quecini, V. Composition and antimicrobial activity of the essential oils from a wide range of species from the Atlantic Rainforest in Brazil. Facta Univ. Ser. Phys. Chem. Technol. 2018, 16, 77. [Google Scholar]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper Species: A Comprehensive Review on Their Phytochemistry, Biological Activities and Applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Oh, D.-G.; Singh, D.; Lee, J.S.; Lee, S.; Lee, C.H. Exploring the metabolomic diversity of plant species across spatial (leaf and stem) components and phylogenic groups. BMC Plant Biol. 2020, 20, 39. [Google Scholar] [CrossRef]
- Li, D.; Gaquerel, E. Next-Generation Mass Spectrometry Metabolomics Revives the Functional Analysis of Plant Metabolic Diversity. Annu. Rev. Plant Biol. 2021, 72, 867–891. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.G.; Marquete, R.; Cruz, L.O.; Caldeira-de-Arujo, A.; Mansur, E.; de Lima Moreira, D. DNA damages promoted by the essential oil from leaves of Casearia sylvestris Sw. (Salicaceae). J. Med. Plants Res. 2016, 10, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Spósito, L.; Oda, F.B.; Vieira, J.H.; Carvalho, F.A.; Ramos, M.A.D.S.; de Castro, R.C.; Crevelin, E.J.; Crotti, A.E.M.; Santos, A.G.; da Silva, P.B.; et al. In vitro and in vivo anti-Helicobacter pylori activity of Casearia sylvestris leaf derivatives. J. Ethnopharmacol. 2018, 233, 1–12. [Google Scholar] [CrossRef] [PubMed]
- de Mesquita, M.; Grellier, P.; Mambu, L.; de Paula, J.; Espindola, L. In vitro antiplasmodial activity of Brazilian Cerrado plants used as traditional remedies. J. Ethnopharmacol. 2007, 110, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Laursen, T.; Møller, B.L.; Bassard, J.-E. Plasticity of specialized metabolism as mediated by dynamic metabolons. Trends Plant Sci. 2015, 20, 20–32. [Google Scholar] [CrossRef]
- Knudsen, C.; Gallage, N.J.; Hansen, C.C.; Møller, B.L.; Laursen, T. Dynamic metabolic solutions to the sessile life style of plants. Nat. Prod. Rep. 2018, 35, 1140–1155. [Google Scholar] [CrossRef] [Green Version]
- Valarezo, E.; Gaona-Granda, G.; Morocho, V.; Cartuche, L.; Calva, J.; Meneses, M. Chemical Constituents of the Essential Oil from Ecuadorian Endemic Species Croton ferrugineus and Its Antimicrobial, Antioxidant and α-Glucosidase Inhibitory Activity. Molecules 2021, 26, 4608. [Google Scholar] [CrossRef]
- El-Din, M.I.G.; Youssef, F.S.; Altyar, A.E.; Ashour, M.L. GC/MS Analyses of the Essential Oils Obtained from Different Jatropha Species, Their Discrimination Using Chemometric Analysis and Assessment of Their Antibacterial and Anti-Biofilm Activities. Plants 2022, 11, 1268. [Google Scholar] [CrossRef]
- Fang, C.; Luo, J.; Wang, S. The Diversity of Nutritional Metabolites: Origin, Dissection, and Application in Crop Breeding. Front. Plant Sci. 2019, 10, 1028. [Google Scholar] [CrossRef]
- Karunanithi, P.S.; Zerbe, P. Terpene Synthases as Metabolic Gatekeepers in the Evolution of Plant Terpenoid Chemical Diversity. Front. Plant Sci. 2019, 10, 1166. [Google Scholar] [CrossRef]
- Bustos-Segura, C.; Padovan, A.; Kainer, D.; Foley, W.J.; Külheim, C. Transcriptome analysis of terpene chemotypes of Melaleuca alternifolia across different tissues. Plant Cell Environ. 2017, 40, 2406–2425. [Google Scholar] [CrossRef]
- Padovan, A.; Keszei, A.; Hassan, Y.; Krause, S.T.; Köllner, T.G.; Degenhardt, J.; Gershenzon, J.; Külheim, C.; Foley, W.J. Four terpene synthases contribute to the generation of chemotypes in tea tree (Melaleuca alternifolia). BMC Plant Biol. 2017, 17, 160. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zheng, Y.; Zhong, Y.; Wu, Y.; Li, Z.; Xu, L.-A.; Xu, M. Transcriptome analysis and identification of genes related to terpenoid biosynthesis in Cinnamomum camphora. BMC Genom. 2018, 19, 550. [Google Scholar] [CrossRef] [Green Version]
- Chaw, S.-M.; Liu, Y.-C.; Wu, Y.-W.; Wang, H.-Y.; Lin, C.-Y.I.; Wu, C.-S.; Ke, H.-M.; Chang, L.-Y.; Hsu, C.-Y.; Yang, H.-T.; et al. Stout camphor tree genome fills gaps in understanding of flowering plant genome evolution. Nat. Plants 2019, 5, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Kwon, M.; Lee, A.-R.; Ro, D.-K.; Wungsintaweekul, J.; Kim, S.-U. Molecular cloning and functional characterization of three terpene synthases from unripe fruit of black pepper (Piper nigrum). Arch. Biochem. Biophys. 2018, 638, 35–40. [Google Scholar] [CrossRef]
- Sun, P.; Schuurink, R.C.; Caissard, J.-C.; Hugueney, P.; Baudino, S. My Way: Noncanonical Biosynthesis Pathways for Plant Volatiles. Trends Plant Sci. 2016, 21, 884–894. [Google Scholar] [CrossRef]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef]
- Mugula, B.B.; Kiboi, S.K.; Kanya, J.I.; Egeru, A.; Okullo, P.; Curto, M.; Meimberg, H. Knowledge Gaps in Taxonomy, Ecology, Population Distribution Drivers and Genetic Diversity of African Sandalwood (Osyris lanceolata Hochst. & Steud.): A Scoping Review for Conservation. Plants 2021, 10, 1780. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Battesti, V.; Ivorra, S.; Paradis, L.; Aberlenc, F.; Zango, O.; Zehdi-Azouzi, S.; Moussouni, S.; Naqvi, S.A.; Newton, C.; et al. On the necessity of combining ethnobotany and genetics to assess agrobiodiversity and its evolution in crops: A case study on date palms (Phoenix dactylifera L.) in Siwa Oasis, Egypt. Evol. Appl. 2020, 13, 1818–1840. [Google Scholar] [CrossRef] [Green Version]
- Funke, G.; von Graevenitz, A.; Clarridge, J.E., 3rd; Bernard, K.A. Clinical microbiology of coryneform bacteria. Clin. Microbiol. Rev. 1997, 10, 125–159. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat Rev Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poolman, J.T.; Anderson, A.S. Escherichia coli and Staphylococcus aureus: Leading bacterial pathogens of healthcare associated infections and bacteremia in older-age populations. Expert Rev. Vaccines 2018, 17, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef] [PubMed]
- Leigh-de Rapper, S.; Viljoen, A.; van Vuuren, S. Essential Oil Blends: The Potential of Combined Use for Respiratory Tract Infections. Antibiotics 2021, 10, 1517. [Google Scholar] [CrossRef]
- Abers, M.; Schroeder, S.; Goelz, L.; Sulser, A.; Rose, T.S.; Puchalski, K.; Langland, J. Antimicrobial activity of the volatile substances from essential oils. BMC Complement. Med. Ther. 2021, 21, 124. [Google Scholar] [CrossRef]
- Ergüden, B. Phenol group of terpenoids is crucial for antibacterial activity upon ion leakage. Lett. Appl. Microbiol. 2021, 73, 438–445. [Google Scholar] [CrossRef]
- Yamaguchi, T. Antibacterial effect of the combination of terpenoids. Arch. Microbiol. 2022, 204, 520. [Google Scholar] [CrossRef]
- Wijesundara, N.M.; Rupasinghe, H. Essential oils from Origanum vulgare and Salvia officinalis exhibit antibacterial and anti-biofilm activities against Streptococcus pyogenes. Microb. Pathog. 2018, 117, 118–127. [Google Scholar] [CrossRef]
- Sim, J.X.F.; Khazandi, M.; Chan, W.Y.; Trott, D.J.; Deo, P. Antimicrobial activity of thyme oil, oregano oil, thymol and carvacrol against sensitive and resistant microbial isolates from dogs with otitis externa. Vet. Dermatol. 2019, 30, 524-e159. [Google Scholar] [CrossRef]
- Scandorieiro, S.; Rodrigues, B.C.D.; Nishio, E.K.; Panagio, L.A.; de Oliveira, A.G.; Durán, N.; Nakazato, G.; Kobayashi, R.K.T. Biogenic Silver Nanoparticles Strategically Combined With Origanum vulgare Derivatives: Antibacterial Mechanism of Action and Effect on Multidrug-Resistant Strains. Front. Microbiol. 2022, 13, 842600. [Google Scholar] [CrossRef]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of Action of Melaleuca alternifolia (Tea Tree) Oil on Staphylococcus aureus Determined by Time-Kill, Lysis, Leakage, and Salt Tolerance Assays and Electron Microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [Green Version]
- Jardim Botânico do Rio de Janeiro; Lista de Espécies da Flora do Brasil. REFLORA. (In Portuguese). 2015. Available online: http://floradobrasil.jbrj.gov.br (accessed on 26 February 2022).
- Brazilian Pharmacopeia. Farmacopeia Brasileira, 6th ed.; ANVISA: Brasília, Brazil, 2019.
- Adams, R.P. Identification of Essential Oil Components by Gas Cromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; 811p. [Google Scholar]
- Van den Dool, E.; Kratz, P. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatog. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; Dezonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Determining Bactericidal Activity of Antimicrobial Agents; Approved Guideline; CLSI Document M26-A; CLSI: Wayne, PA, USA, 1999. [Google Scholar]
- Turgis, M.; Han, J.; Caillet, S.; Lacroix, M. Antimicrobial activity of mustard essential oil against Escherichia coli O157:H7 and Salmonella typhi. Food Control 2009, 20, 1073–1079. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 10 June 2022).
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Scrucca, L.; Fop, M.; Murphy, T.B.; Raftery, A.E. mclust 5: Clustering, Classification and Density Estimation Using Gaussian Finite Mixture Models. R J. 2016, 8, 289–317. [Google Scholar] [CrossRef] [Green Version]
- Harrell, F., Jr. Package ‘Hmisc’. 2019. Available online: https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (accessed on 10 June 2022).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. Version 0.84. 2017. Available online: https://github.com/taiyun/corrplot (accessed on 10 June 2022).
- Mattar, V.T.; Borioni, J.L.; Hollmann, A.; Rodriguez, S.A. Insecticidal activity of the essential oil of Schinus areira against Rhipibruchus picturatus (F.) (Coleoptera: Bruchinae), and its inhibitory effects on acetylcholinesterase. Pestic. Biochem. Physiol. 2022, 185. [Google Scholar] [CrossRef]
- De Groot, A.C.; Schmidt, E. Tea tree oil: Contact allergy and chemical composition. Contact Dermat. 2016, 75, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Moura-Costa, G.F.; Nocchi, S.R.; Ceole, L.F.; de Mello, J.C.P.; Nakamura, C.V.; Filho, B.P.D.; Temponi, L.G.; Ueda-Nakamura, T. Antimicrobial activity of plants used as medicinals on an indigenous reserve in Rio das Cobras, Paraná, Brazil. J. Ethnopharmacol. 2012, 143, 631–638. [Google Scholar] [CrossRef]
- Gehrke, I.T.; Neto, A.T.; Pedroso, M.; Mostardeiro, C.P.; Da Cruz, I.B.; Silva, U.F.; Ilha, V.; Dalcol, I.I.; Morel, A.F. Antimicrobial activity of Schinus lentiscifolius (Anacardiaceae). J. Ethnopharmacol. 2013, 148, 486–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uliana, M.P.; Fronza, M.; da Silva, A.G.; Vargas, T.S.; Andrade, T.; Scherer, R. Composition and biological activity of Brazilian rose pepper (Schinus terebinthifolius Raddi) leaves. Ind. Crops Prod. 2016, 83, 235–240. [Google Scholar] [CrossRef]
- Lima, I.M.D.S.F.; Zagmignan, A.; Santos, D.M.; Maia, H.S.; Silva, L.D.S.; Cutrim, B.D.S.; Vieira, S.L.; Filho, C.M.B.; de Sousa, E.M.; Napoleão, T.H.; et al. Schinus terebinthifolia leaf lectin (SteLL) has anti-infective action and modulates the response of Staphylococcus aureus-infected macrophages. Sci. Rep. 2019, 9, 18159. [Google Scholar] [CrossRef] [Green Version]
- Cascaes, M.M.; De Moraes, A.B.; Cruz, J.N.; Franco, C.D.J.P.; E Silva, R.C.; Nascimento, L.D.D.; Ferreira, O.O.; dos Anjos, T.O.; de Oliveira, M.S.; Guilhon, G.M.S.P.; et al. Phytochemical Profile, Antioxidant Potential and Toxicity Evaluation of the Essential Oils from Duguetia and Xylopia Species (Annonaceae) from the Brazilian Amazon. Antioxidants 2022, 11, 1709. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.S.; Esquissato, G.N.M.; Costa, E.V.; Nogueira, P.C.D.L.; de Castro-Prado, M.A.A. Mutagenic and cytostatic activities of the Xylopia laevigata essential oil in human lymphocytes. Nat. Prod. Res. 2019, 35, 1682–1685. [Google Scholar] [CrossRef]
- Moura, A.P.G.; Beltrão, D.M.; Pita, J.C.L.R.; Xavier, A.L.; Brito, M.T.; de Sousa, T.K.G.; Batista, L.M.; de Carvalho, J.E.; Ruiz, A.L.T.G.; Della Torre, A.; et al. Essential oil from fruit of Xylopia langsdorffiana: Antitumour activity and toxicity. Pharm. Biol. 2016, 54, 3093–3102. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.N.G.D.; Junqueira, J.G.M.; Terezan, A.P.; Severino, R.; Silva, T.; Martins, C.H.G.; Severino, V.G.P.; Cacuro, T.A.; Waldman, W. Chemical Composition and Antimicrobial Activity of Essential Oils from Xylopia aromatica (Annonaceae) Flowers and Leaves. Rev. Virtual Quim 2018, 10, 1578–1590. [Google Scholar] [CrossRef]
- Ricardo, L.M.; de Paula-Souza, J.; Andrade, A.; Brandão, M.G. Plants from the Brazilian Traditional Medicine: Species from the books of the Polish physician Piotr Czerniewicz (Pedro Luiz Napoleão Chernoviz, 1812–1881). Rev. Bras. de Farm. 2017, 27, 388–400. [Google Scholar] [CrossRef]
- Meneguelli, A.Z.; Camargo, E.E.S.; Buccini, D.F.; Roriz, B.C.; Cerqueira, G.R.; Moreno, S.E. Ethnopharmacological and botanical evaluation of medicinal plants used by Brazilian Amazon Indian community. Interações (Campo Grande) 2020. [Google Scholar] [CrossRef]
- Cruz-Galvez, A.M.; Gómez-Aldapa, C.A.; Villagómez-Ibarra, J.R.; Chavarría-Hernández, N.; Rodríguez-Baños, J.; Rangel-Vargas, E.; Castro-Rosas, J. Antibacterial effect against foodborne bacteria of plants used in traditional medicine in central Mexico: Studies in vitro and in raw beef. Food Control 2013, 32, 289–295. [Google Scholar] [CrossRef]
- Balakrishnan, R.; Cho, D.-Y.; Su-Kim, I.; Choi, D.-K. Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants 2020, 9, 962. [Google Scholar] [CrossRef]
- Bueno, G.; Rico, S.L.C.; Périco, L.L.; Ohara, R.; Rodrigues, V.P.; Emílio-Silva, M.T.; Assunção, R.; da Rocha, L.R.M.; Nunes, D.S.; Besten, M.A.; et al. The essential oil from Baccharis trimera (Less.) DC improves gastric ulcer healing in rats through modulation of VEGF and MMP-2 activity. J. Ethnopharmacol. 2021, 271, 113832. [Google Scholar] [CrossRef]
- Cazella, L.N.; Glamoclija, J.; Soković, M.; Gonçalves, J.E.; Linde, G.A.; Colauto, N.B.; Gazim, Z.C. Antimicrobial Activity of Essential Oil of Baccharis dracunculifolia DC (Asteraceae) Aerial Parts at Flowering Period. Front. Plant Sci. 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Zuccolotto, T.; Bressan, J.; Lourenço, A.V.F.; Bruginski, E.; Veiga, A.; Marinho, J.V.N.; Raeski, P.A.; Heiden, G.; Salvador, M.J.; Murakami, F.S.; et al. Chemical, Antioxidant, and Antimicrobial Evaluation of Essential Oils and an Anatomical Study of the Aerial Parts from Baccharis Species (Asteraceae). Chem. Biodivers. 2019, 16, e1800547. [Google Scholar] [CrossRef]
- Camargo, J.G.S.D. Desreplicação dos Extratosativos de Cyrtocymurascorpioides (Asteraceae) contra Candida spp. e Trichomonas vaginalis. 2020. MSc. Dissertation, Instituto de Biociências, Letras e CiênciasExatas (IBILCE), UNESP, São José do Rio Preto, SP, Brazil. Available online: http://hdl.handle.net/11449/194314 (accessed on 7 December 2022).
- Santos, N.C.; da Silva, J.E.; Santos, A.C.C.; Dantas, J.D.O.; Tavares, S.R.S.A.; Andrade, V.S.; Oliveira, S.D.D.S.; Blank, A.F.; Araújo, A.P.A.; Bacci, L. Bioactivity of essential oils from Croton grewioides and its major compounds: Toxicity to soybean looper Chrysodeixis includens and selectivity to the predatory stink bug Podisus nigrispinus. Environ. Sci. Pollut. Res. 2022, 1–12. [Google Scholar] [CrossRef]
- Silva, K.; Peruchetti, D.; Sirtoli, G.; Takiya, C.; Pinheiro, A.; Leal-Cardoso, J.; Caruso-Neves, C. High Doses of Essential Oil of Croton Zehntneri Induces Renal Tubular Damage. Plants 2021, 10, 1400. [Google Scholar] [CrossRef]
- Almeida-Pereira, C.S.; Nogueira, P.C.D.L.; Barbosa, A.A.T.; Nizio, D.A.D.C.; Arrigoni-Blank, M.D.F.; Sampaio, T.S.; Alves, R.P.; de Araujo-Couto, H.G.S.; Feitosa-Alcantara, R.B.; de Melo, J.O.; et al. Chemical composition and antimicrobial activity of essential oils of a Croton tetradenius Baill. germplasm. J. Essent. Oil Res. 2019, 31, 379–389. [Google Scholar] [CrossRef]
- Rocha, A.R.d.S.; Sousa, H.G.; Júnior, E.P.D.V.; de Lima, F.L.; Costa, A.S.; de Araújo, A.R.; Leite, J.R.S.; Martins, F.A.; Oliveira, M.B.P.; Plácido, A.; et al. Extracts and fractions of Croton L. (Euphorbiaceae) species with antimicrobial activity and antioxidant potential. LWT 2021, 139. [Google Scholar] [CrossRef]
- Silva, N.N.S.; Silva, J.R.A.; Alves, C.N.; Andrade, E.H.A.; da Silva, J.K.R.; Maia, J.G.S. Acetylcholinesterase Inhibitory Activity and Molecular Docking Study of 1-Nitro-2-Phenylethane, the Main Constituent of Aniba canelilla Essential Oil. Chem. Biol. Drug Des. 2014, 84, 192–198. [Google Scholar] [CrossRef]
- da Silva, J.K.R.; da Trindade, R.C.S.; Maia, J.G.S.; Setzer, W.N. Chemical Composition, Antioxidant, and Antimicrobial Activities of Essential Oils of Endlicheria arenosa (Lauraceae) from the Amazon. Nat. Prod. Commun. 2016, 11, 1934578X1601100537. [Google Scholar] [CrossRef]
- de Moura, V.M.; Guimarães, N.D.C.; Batista, L.T.; Freitas-De-Sousa, L.A.; Martins, J.D.S.; de Souza, M.C.S.; de Almeida, P.D.O.; Monteiro, W.M.; de Oliveira, R.B.; Dos-Santos, M.C.; et al. Assessment of the anti-snakebite properties of extracts of Aniba fragrans Ducke (Lauraceae) used in folk medicine as complementary treatment in cases of envenomation by Bothrops atrox. J. Ethnopharmacol. 2018, 213, 350–358. [Google Scholar] [CrossRef] [PubMed]
- da Silva, Y.C.; Silva, E.M.S.; Fernandes, N.; Lopes, N.L.; Orlandi, P.P.; Nakamura, C.V.; Costa, E.; Júnior, V.F.D.V. Antimicrobial substances from Amazonian Aniba (Lauraceae) species. Nat. Prod. Res. 2019, 35, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Longhini, R.; Lonni, A.A.; Sereia, A.L.; Krzyzaniak, L.M.; Lopes, G.C.; de Mello, J.C.P. Trichilia catigua: Therapeutic and cosmetic values. Rev. Bras. de Farm. 2017, 27, 254–271. [Google Scholar] [CrossRef] [Green Version]
- da Silva, L.L.; de Almeida, R.; e Silva, F.T.; Verícimo, M.A. Review on the therapeutic activities of the genus Trichilia. Res. Soc. Dev. 2021, 10. [Google Scholar] [CrossRef]
- Cordeiro, R.M.; Silva, A.P.D.S.E.; Pinto, R.H.H.; da Costa, W.A.; da Silva, S.H.M.; Pinheiro, W.B.D.S.; Arruda, M.S.P.; Junior, R.N.C. Supercritical CO2 extraction of ucuúba (Virola surinamensis) seed oil: Global yield, kinetic data, fatty acid profile, and antimicrobial activities. Chem. Eng. Commun. 2018, 206, 86–97. [Google Scholar] [CrossRef]
- González-Rodríguez, M.; Ruiz-Fernández, C.; Francisco, V.; Eldjoudi, D.A.; Ramadan, F.Y.; Cordero-Barreal, A.; Pino, J.; Lago, F.; Campos-Toimil, M.; Carvalho, G.R.; et al. Pharmacological Extracts and Molecules from Virola Species: Traditional Uses, Phytochemistry, and Biological Activity. Molecules 2021, 26, 792. [Google Scholar] [CrossRef]
- Ribeiro, C.L.; Silva, R.M.; Fernandes, R.D.M.N.; Araújo, M.R.; Soares, I.M.; da Silva, J.F.M.; Nascimento, G.N.L.D.; Pimenta, R.S.; Scapin, E. Chemical assessment, antioxidant and antimicrobial of leafs extracts of Virola sebifera, an Amazonian medicinal plant. Res. Soc. Dev. 2021, 10. [Google Scholar] [CrossRef]
- de Moraes, A.B.; Ferreira, O.O.; da Costa, L.S.; Almeida, L.Q.; Varela, E.L.P.; Cascaes, M.M.; Franco, C.D.J.P.; Percário, S.; Nascimento, L.D.D.; de Oliveira, M.S.; et al. Phytochemical Profile, Preliminary Toxicity, and Antioxidant Capacity of the Essential Oils of Myrciaria floribunda (H. West ex Willd.) O. Berg. and Myrcia sylvatica (G. Mey) DC. (Myrtaceae). Antioxidants 2022, 11, 2076. [Google Scholar] [CrossRef]
- Magalhães, B.Q.; Machado, F.P.; Sanches, P.S.; Lima, B.; Falcão, D.Q.; von Ranke, N.; Bello, M.L.; Rodrigues, C.R.; Santos, M.G.; Rocha, L.; et al. Eugenia sulcata (Myrtaceae) Nanoemulsion Enhances the Inhibitory Activity of the Essential Oil on P2X7R and Inflammatory Response In Vivo. Pharmaceutics 2022, 14, 911. [Google Scholar] [CrossRef]
- de Araújo, F.F.; Neri-Numa, I.A.; Farias, D.D.P.; da Cunha, G.R.M.C.; Pastore, G.M. Wild Brazilian species of Eugenia genera (Myrtaceae) as an innovation hotspot for food and pharmacological purposes. Food Res. Int. 2019, 121, 57–72. [Google Scholar] [CrossRef]
- Macedo, J.G.F.; Rangel, J.M.L.; Santos, M.D.O.; Camilo, C.J.; da Costa, J.G.M.; Souza, M.M.D.A. Therapeutic indications, chemical composition and biological activity of native Brazilian species from Psidium genus (Myrtaceae): A review. J. Ethnopharmacol. 2021, 278, 114248. [Google Scholar] [CrossRef]
- Vechi, G.; Tenfen, A.; Capusiri, E.S.; Gimenez, A.; Cechinel-Filho, V. Antiparasitic activity of two Brazilian plants: Eugenia mattosii and Marlierea eugeniopsoides. Nat. Prod. Res. 2020, 35, 4876–4880. [Google Scholar] [CrossRef]
- Maiolini, T.C.S.; Rosa, W.; Miranda, D.D.O.; Costa-Silva, T.A.; Tempone, A.G.; Bueno, P.C.P.; Dias, D.F.; de Paula, D.A.C.; Sartorelli, P.; Lago, J.H.G.; et al. Essential Oils from Different Myrtaceae Species from Brazilian Atlantic Forest Biome–Chemical Dereplication and Evaluation of Antitrypanosomal Activity. Chem. Biodivers. 2022, 19. [Google Scholar] [CrossRef]
- de Oliveira, A.C.; Simões, R.C.; Lima, C.A.P.; da Silva, F.M.A.; Nunomura, S.M.; Roque, R.A.; Tadei, W.P.; Nunomura, R.C.S. Essential oil of Piper purusanum C.DC (Piperaceae) and its main sesquiterpenes: Biodefensives against malaria and dengue vectors, without lethal effect on non-target aquatic fauna. Environ. Sci. Pollut. Res. 2022, 29, 47242–47253. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, J.; Famous, E.; Pan, S.; Peng, X.; Tian, J. Antioxidant, hepatoprotective and antifungal activities of black pepper (Piper nigrum L.) essential oil. Food Chem. 2020, 346, 128845. [Google Scholar] [CrossRef]
- Da Silva, J.K.; da Trindade, R.; Alves, N.S.; Figueiredo, P.L.; Maia, J.G.S.; Setzer, W.N. Essential Oils from Neotropical Piper Species and Their Biological Activities. Int. J. Mol. Sci. 2017, 18, 2571. [Google Scholar] [CrossRef] [Green Version]
- de Pascoli, I.C.; dos Anjos, M.M.; da Silva, A.A.; Lorenzetti, F.B.; Cortez, D.A.G.; Mikcha, J.M.G.; Nakamura, T.U.; Nakamura, C.V.; Filho, B.A.D.A. Piperaceae extracts for controlling Alicyclobacillus acidoterrestris growth in commercial orange juice. Ind. Crops Prod. 2018, 116, 224–230. [Google Scholar] [CrossRef]
- Majolo, C.; Monteiro, P.C.; Nascimento, A.V.P.D.; Chaves, F.C.M.; Gama, P.E.; Bizzo, H.R.; Chagas, E.C. Essential Oils from Five Brazilian Piper Species as Antimicrobials Against Strains of Aeromonas hydrophila. J. Essent. Oil Bear. Plants 2019, 22, 746–761. [Google Scholar] [CrossRef]
- dos Santos, A.L.M.; Araújo, F.A.M.; Matisui, D.S.; da Costa, L.A.M.A.; Macêdo, A.J.; de Lucena, J.M.V.M. Antimicrobial and antibiofilm properties of essential oils from Piper marginatum Jacq. Res. Soc. Dev. 2021, 10. [Google Scholar] [CrossRef]
- Shoorvarzi, S.N.; Shahraki, F.; Shafaei, N.; Karimi, E.; Oskoueian, E. Citrus aurantium L. bloom essential oil nanoemulsion: Synthesis, characterization, cytotoxicity, and its potential health impacts on mice. J. Food Biochem. 2020, 44, e13181. [Google Scholar] [CrossRef]
- Volpato, G.T.; Francia-Farje, L.A.; Damasceno, D.C.; Oliveira, R.V.; Hiruma-Lima, C.A.; Kempinas, W.G. Effect of essential oil from Citrus aurantium in maternal reproductive outcome and fetal anomaly frequency in rats. An. Da Acad. Bras. De Ciências 2015, 87, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlanda, J.F.; Nascimento, A. Chemical composition and antibacterial activity of Ruta graveolens L. (Rutaceae) volatile oils, from São Luís, Maranhão, Brazil. S. Afr. J. Bot. 2015, 99, 103–106. [Google Scholar] [CrossRef]
- Fernandes, T.S.; Copetti, D.; Carmo, G.D.; Neto, A.T.; Pedroso, M.; Silva, U.F.; Mostardeiro, M.A.; Burrow, R.E.; Dalcol, I.I.; Morel, A.F. Phytochemical analysis of bark from Helietta apiculata Benth and antimicrobial activities. Phytochemistry 2017, 141, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.B.; Dos Santos, N.O.; Pascon, R.C.; Vallim, M.; Figueiredo, C.R.; Martins, R.C.C.; Sartorelli, P. Chemical Composition and In Vitro Cytotoxic and Antimicrobial Activities of the Essential Oil from Leaves of Zanthoxylum monogynum St. Hill (Rutaceae). Medicines 2017, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Paula, R.C.; da Silva, S.M.; Faria, K.F.; Frézard, F.; Moreira, C.P.D.S.; Foubert, K.; Lopes, J.C.D.; Campana, P.R.V.; Rocha, M.P.; Silva, A.F.; et al. In vitro antileishmanial activity of leaf and stem extracts of seven Brazilian plant species. J. Ethnopharmacol. 2018, 232, 155–164. [Google Scholar] [CrossRef]
- Filho, D.E.S.; De Sousa, J.B.; Dos Santos, H.S.; Fontenelle, R.O.D.S. Compostos químicos isolados de extratos e óleos essenciais do gênero Zanthoxylum Linnaeus (Rutaceae) e seu potencial antimicrobiano. Hoehnea 2020, 47. [Google Scholar] [CrossRef]
- Silva, S.L.D.; Chaar, J.D.S.; Figueiredo, P.D.M.S.; Yano, T. Cytotoxic evaluation of essential oil from CaseariasylvestrisSw on human cancer cells and erythrocytes. ActaAmazonica 2008, 38, 107–112. [Google Scholar]
- Flaviane, G.P.; Ronaldo, M.; Letícia, O.C.; Adriano, C.-D.; Elisabeth, M.; Davyson, D.L.M.; Pereira, F.G.; Marquete, R.; Cruz, L.O.; Caldeira-De-Arujo, A.; et al. DNA damages promoted by the essential oil from leaves of Casearia sylvestris Sw. (Salicaceae). J. Med. Plants Res. 2016, 10, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.M.; Fratucelli, D.O.; Bueno, P.C.P.; De Castro, M.K.V.; Francisco, A.A.; Cavalheiro, A.J.; Klein, M.I. Antimicrobial and antibiofilm activities of Casearia sylvestris extracts from distinct Brazilian biomes against Streptococcus mutans and Candida albicans. BMC Complement. Altern. Med. 2019, 19, 308. [Google Scholar] [CrossRef]
- Ribeiro, I.C.D.O.; Mariano, E.G.A.; Careli, R.T.; Morais-Costa, F.; De Sant’Anna, F.M.; Pinto, M.S.; De Souza, M.R.; Duarte, E.R. Plants of the Cerrado with antimicrobial effects against Staphylococcus spp. and Escherichia coli from cattle. BMC Vet. Res. 2018, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- da Cruz, J.E.R.; Saldanha, H.C.; e Freitas, G.R.O.; Morais, E.R. A review of medicinal plants used in the Brazilian Cerrado for the treatment of fungal and bacterial infections. J. Herb. Med. 2021, 31, 100523. [Google Scholar] [CrossRef]
- Barros, L.M.; Duarte, A.E.; Morais-Braga, M.F.B.; Waczuk, E.P.; Vega, C.; Leite, N.F.; De Menezes, I.R.A.; Coutinho, H.D.M.; Rocha, J.B.T.; Kamdem, J.P. Chemical Characterization and Trypanocidal, Leishmanicidal and Cytotoxicity Potential of Lantana camara L. (Verbenaceae) Essential Oil. Molecules 2016, 21, 209. [Google Scholar] [CrossRef] [Green Version]
- Postay, L.F.; Cabral, D.S.; Heringer, O.A.; Vieira, L.V.; de Moraes, L.R.; Freitas, G.; Gomes, L.C. The effectiveness of surfactants applied with essential oil of Lippia alba in the anesthesia of Nile tilapia (Oreochromis niloticus) and their toxicity assessment for fish and mammals. Environ. Sci. Pollut. Res. 2020, 28, 10224–10233. [Google Scholar] [CrossRef]
- de Souza, R.C.; da Costa, M.M.; Baldisserotto, B.; Heinzmann, B.M.; Schmidt, D.; Caron, B.O.; Copatti, C.E. Antimicrobial and synergistic activity of essential oils of Aloysia triphylla and Lippia alba against Aeromonas spp. Microb. Pathog. 2017, 113, 29–33. [Google Scholar] [CrossRef]
- Zamora, C.M.P.; Torres, C.A.; Nuñez, M.B. Antimicrobial Activity and Chemical Composition of Essential Oils from Verbenaceae Species Growing in South America. Molecules 2018, 23, 544. [Google Scholar] [CrossRef] [Green Version]
- Nader, T.T.; Leonel, A.H.; Henrique, C.Y.; Contini, S.H.T.; Crevelin, E.J.; França, S.D.C.; Berton, B.W.; Pereira, A.M.S. Dynamized Aloysia Polystachya (Griseb.) Essential Oil: A Promising Antimicrobial Product. Homeopathy 2022. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Species | Sample/Herbarium Code | Location (Coordinates, Elevation) | Yield (%) | Major Components (%) |
|---|---|---|---|---|
| Anacardiaceae | ||||
| Schinus terebinthifolius Raddi | R1772/IAC 47521 | Ribeirão Preto (47°51′58.72″ S, 21°12′52.36″ W, 570 m) | 0.26 | α-phellandrene (23.2); α-pinene (18.2); β-phellandrene (16.8) |
| Annonaceae | ||||
| Annona dioica A.St.-Hil. | Lp06101603/IAC 47955 | Votuporanga (50°3′55.60″ S, 20°27′41.20″ W, 463 m) | 0.37 | bicyclogermacrene (30.1); germacrene D (21.0); trans-caryophyllene (12.2) |
| Guatteria australis A.St.-Hil. | R1598/ IAC 46831 | Ubatuba (45°7′39.30″ S, 23°25′18.60″ W, 26 m) | 0.14 | spathulenol (27.4); caryophyllene oxide (18.8) |
| Xylopia aromatica (Lam.) Mart. | Lp6101712/ IAC 47969 | Votuporanga (50°3′34.99″ S, 20°27′15.20″ W, 4 m) | 0.16 | limonene (71.7) |
| Xylopia brasiliensis Spreng. | R1739/IAC 47266 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 0.17 | 1,8-cineole (11.1); spathulenol (28.3) |
| Araceae | ||||
| Monstera cf. adansonii Schott | L52008/IAC 47079 | Ubatuba (45°7′47.57″ S, 23°24′49.97″ W, 180 m) | 0.14 | β-phellandrene (36.7), α-pinene (17.2), 2 tridecanone (17.0) |
| Araliaceae | ||||
| Dendropanax cuneatus (DC.) Decne. & Planch. | C010/IAC 47099 | Jundiai (46°55′40.69″ S, 23°6′42.70″ W, 770 m) | 0.12 | caryophyllene oxide (15.6), trans caryophyllene (13.2), β pinene (10.9) |
| Dendropanax cuneatus (DC.) Decne. & Planch. | Lp6101818/IAC 47975 | Adamantina (51°9′6.80″ S, 21°39′47.01″ W, 380 m) | 0.004 | spathulenol (22.1), trans caryophyllene (18.4), bicyclogermacrene (15.7), δ-3-carene (12.5) |
| Dendropanax cuneatus (DC.) Decne. & Planch. | R1770/IAC 47519 | Mococa (46°59′55.30″ S, 21°25′24.56″ W, 568 m) | 0.21 | bicyclogermacrene (32.8) |
| Asteraceae | ||||
| Baccharis dracunculifolia DC. | R1755/IAC 47282 | Pariquera-Açu (46°59′44.79″ S, 24°37′15.32″ W, 25 m) | 0.42 | trans-nerolidol (30.5), β-copaen-4-α-ol (12.0), limonene (11.6) |
| Baccharis dracunculifolia DC. | R1773/IAC 47522 | Ribeirão Preto (47°52′12.64″ S, 21°11′27.63″ W, 557 m) | 0.54 | trans-nerolidol (27.3), limonene (17.4) |
| Cyrtocymura scorpioides (Lam.) H. Rob. | C005/IAC 47097 | Jundiai (46°55′40.69″ S, 23°6′42.70″ W, 770 m) | 0.32 | germacrene D (36.1), β-pinene (26.6) |
| Euphorbiacae | ||||
| Croton celtidifolius Baill. | Lp52912/IAC 29030 | Monte Alegre do Sul (46°40′30.18″ S, 22°41′58.52″ W, 743 m) | 0.16 | cis-β-guaiene (15.8), germacrene D (11.7), trans-nerolidol (11.1) |
| Croton floribundus Spreng. | R4167/IAC 46976 | Campinas (47°4′3.36″ S, 22°51′45.72″ W, 670 m) | 0.09 | trans-caryophyllene (21.9), caryophyllene oxide (13.7) |
| Croton urucurana Baill. | R1768/IAC 47517 | Mococa (46°59′55.30″ S, 21°25′24.56″ W, 568 m) | 0.11 | bicyclogermagrene (43.4), germacrene D (24.0) |
| Croton warmingii Müll. Arg. | R4165/IAC 46974 | Campinas (47°4′3.57″ S, 22°51′45.79″ W, 670 m) | 0.15 | bicyclogermacrene (17.4), trans-caryophyllene (16.8) |
| Lauraceae | ||||
| Aiouea sp. | R1736/IAC 47263 | Pariquera-Açu (46°59′44.79″ S, 24°37′15.32″ W, 25 m) | 0.28 | α-phellandrene (24.5), trans-nerolidol (19.4) |
| Aniba viridis Mez | R1746/IAC 47273 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 0.59 | benzyl salicylate (23.4), benzyl benzoate (14.1) |
| Aniba viridis Mez | Lp51907/ AC 47071 | Ubatuba (45°7′39.29″ S, 23°25′18.59″ W, 29 m) | 0.42 | linalool (11.1), trans-nerolidol (73.1) |
| Endlicheria paniculata (Spreng.) J.F.Macbr. | R1622/IAC 46801 | Campinas (47°4′3.30″ S, 22°51′50.00″ W, 652 m) | 0.05 | α-selinene (34.5), spathulenol (15.3), γ-muurolene (11.8) |
| Nectandra megapotamica (Spreng.) Mez | Lp6101820/IAC 47986 | Adamantina (51°9′7.50″ S, 21°39′47.00″ W, 349 m) | 0.13 | cis-β-guaiene (23.4), spathulenol (15.6) |
| Nectandra megapotamica (Spreng.) Mez | R1761/IAC 47510 | Mococa (46°58′51.65″ S, 21°26′53.71″ W, 600 m) | 0.27 | α-pinene (27.1), β-pinene (28.2), bicyclogermagrene (16.4) |
| Nectandra megapotamica (Spreng.) Mez | R1774/IAC 47523 | Ribeirão Preto (47°52′12.64″ S, 21°11′27.63″ W, 557 m) | 0.10 | oxygenated sesquiterpene (28.1), α-pinene (18.7), β-pinene (17.3) |
| Nectranda megapotamica (Spreng.) Mez | Lp052902/IAC 47084 | Monte Alegre do Sul (46°39′57.60″ S, 22°42′15.12″ W, 778 m) | 0.07 | cis-β-guaiene (22.7), α-pinene (21.2), β-pinene (18.5) |
| Ocotea odorifera (Vell.) Rohwer | R1747/IAC 47274 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 2.88 | camphor (50.5), methyl-eugenol (20.0) |
| Meliaceae | ||||
| Trichilia elegans A.Juss. | Lp6101709/IAC 47961 | Votuporanga (50°3′30.60″ S, 20°27′27.50″ W, 479 m) | 0.28 | germacrene B (44.3) |
| Myristicaceae | ||||
| Virola bicuhyba (Schott ex Spreng.) Warb. | Lp52003/IAC 49465 | Ubatuba (45°7′20.86″ S, 23°24′34.06″ W, 50 m) | 0.14 | cis-β-guaiene (21.4), trans-caryophyllene (18.1) |
| Myrtaceae | ||||
| Calyptranthes lanceolata O.Berg | R1745/IAC 47272 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 0.12 | methyl eugenol (80.4) |
| Calyptranthes lucida Mart. ex DC. | Lp52009/IAC 47080 | Ubatuba (45°7′39.29″ S, 23°25′18.59″ W, 30 m) | 0.19 | caryophyllene oxide (17.3), trans-caryophyllene (16.9), bicyclogermacrene (12.4) |
| Campomanesia guavirota (DC.) Kiaersk. | Lp6101822/IAC 47988 | Adamantina (51°9′8.20″ S, 21°39′46.50″ W, 355 m) | 0.33 | α-pinene (12.3), linalool (11.8) |
| Eugenia moraviana O.Berg. | Lp06101816/ IAC 47973 | Adamantina (51°9′6.19″ S, 21°39′47.80″ W, 373 m) | 0.04 | β-pinene (16.2), trans-caryophyllene (14.2), β elemene (11.0) |
| Eugenia neoverrucosa Sobral | R1626/IAC 46825 | Campinas (47°4′0.30″ S, 22°51′52.03″ W, 650 m) | 0.42 | α-pinene (94.5) |
| Eugenia prasina O.Berg | Lp51905/IAC 47069 | Ubatuba (45°7′39.29″ S, 23°25′18.59″ W, 29 m) | 0.28 | limonene (61.4), α-pinene (12.6) |
| Eugenia pyriformis Cambess. | C009/IAC 34660 | Jundiai (46°55′40.69″ S, 23°6′42.69″ W, 770 m) | 0.17 | β-pinene (39.7), α-pinene (31.5) |
| Eugenia myrcianthes Nied. | Lp6101714/ IAC 47971 | Votuporanga (50°3′30.10″ S, 20°27′20.99″ W, 488 m) | 0.06 | β-copaen-4-α-ol (31.7) |
| Marlierea exocoriata Mart. | Lp52006/IAC 47077 | Ubatuba (45°7′8.70″ S, 23°24′32.52″ W, 38 m) | 0.28 | α-pinene (37.6), β-pinene (18.2), sabinene (11.2) |
| Myrcia spectabilis DC. | Lp051901/IAC 47045 | Ubatuba (45°7′39.29″ S, 23°25′18.59″ W, 29 m) | 0.41 | trans-cis-farnesol (52.1), cis-cis-farnesol (41.1) |
| Myrcia splendens (Sw.) DC. | C007/IAC 37365 | Jundiai (46°55′40.51″ S, 23°6′42.52″ W, 770 m) | 0.21 | α-pinene (28.1), germacrene D (20.9) |
| Myrcia tomentosa (Aubl.) DC. | Lp06101817/IAC 47974 | Adamantina (51°9′6.40″ S, 21°39′46.70″ W, 370 m) | 0.15 | germacrene D (33.09%), trans-caryophyllene (20.41%), |
| Piperaceae | ||||
| Piper aduncum L. | Lp52911/IAC 47090 | Monte Alegre do Sul (46°40′20.99″ S, 22°42′0.36″ W, 743 m) | 0.51 | spathulenol (10.6), valencene (9.7), α-pinene (6.4), asaricin (14.9), safrole (13.3) |
| Piper aduncum L. | Lp6101606/IAC 47958 | Votuporanga (50°3′53.10″ S, 20°27′46.30″ W, 458 m) | 1.52 | asaricin (80.1), safrole (10.8) |
| Piper aduncum L. | Lp6101608/IAC 47960 | Votuporanga (50°3′53.10″ S, 20°27′46.60″ W, 465 m) | 1.55 | asaricin (73.4), safrole (10.5) |
| Piper amalago L. | Lp06091202/IAC 32056 | Campinas (47°4′2.30″ S, 22°51′53.70″ W, 664 m) | 0.20 | β-phellandrene (39.3), α-pinene (14.8), germacrene D (11.7) |
| Piper amalago L. | Lp06091206/IAC 46823 | Campinas (50°3′53.09″ S, 20°27′46.30″ W, 458 m) | 0.36 | β-phellandrene (15.9), α-pinene (6.7), sabinene (6.3), bicyclogermagrene (20.8), spathulenol (9.1) |
| Piper amalago L. | R1763/IAC 47512 | Mococa (46°58′51.65″ S, 21°26′53.71″ W, 600 m) | 0.26 | β-phellandrene (33.1), α-pinene (11.7), bicyclogermagrene (15.0) |
| Piper amalago L. | Lp6101821/IAC 47987 | Adamantina (51°9′7.89″ S, 21°39′47.19″ W, 349 m) | 0.23 | β-phellandrene (12.3), sabinene (8.2), myrcene (6.8), bicyclogermagrene (19.4); γ-muurolene (5.9), spathulenol (5.6) |
| Piper amplum Kunth. | R1740/IAC 7267 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 0.38 | α-pinene (18.1), cis-β-ocimene (10.5), limonene (8.6), trans-caryophyllene (8.8), germacrene D (5,5) |
| Piper cernuum Vell. | L51904/IAC 7068 | Ubatuba (45°7′39.04″ S, 23°25′18.52″ W, 30 m) | 0.32 | α-pinene (10.0), camphene (6.3), dihydro-β-agarofuran (28.7), 10-epi γ-eudesmol (13.5), 4-epi-cis-dihydro-agarofuran (10.8) |
| Piper cernuum Vell. | R1741/IAC 7268 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 1.84 | dihydro-β-agarofuran (33.8), 10-epi-γ-eudesmol (12.2), α-pinene (11.8), camphene (8.7) |
| Piper crassinervium Kunth. | R1764/IAC 7513 | Mococa (46°59′55.30″ S, 21°25′24.56″ W, 568 m) | 0.53 | β-pinene (11.6), α-pinene (11.5), germacrene D (9,2), trans-caryophyllene (7.8), guaiol (5.5), bicyclogermacrene (5.1) |
| Piper gaudichaudianum Kunth. | R1738/IAC 7265 | Pariquera-Açu (47°52′48.76″ S, 24°36′48.42″ W, 25 m) | 0.16 | trans-nerolidol (17.5), α-pinene (12.2), caryophyllene oxide (8.5), trans-caryophyllene (8.2), β-pinene (7.0), trans-β-guaiene (6.9) |
| Piper leptorum Kunth. | Lp052903/IAC 7085 | Monte Alegre do Sul (46°39′53.99″ S, 22°42′13.32″ W, 778 m) | 0.60 | seychellene (34.7), caryophyllene oxide (12.5) |
| Piper rivinoides Kunth. | L52007/IAC 47078 | Ubatuba (45°7′16.03″ S, 23°25′16.36″ W, 30 m) | 0.63 | α-pinene (73.2), β-pinene (5.2) |
| Piper solmsianum C.DC. | R1633/IAC 46832 | Ubatuba (45°7′8.79″ S, 23°24′32.47″ W, 40 m) | 0.39 | δ-3-carene (66.9), myrcene (26.1), α-pinene (22.7), α-selinene (5.5) |
| Piper umbellatum (L.) | R4169/IAC 46978 | Campinas (47°4′4.69″ S, 22°51′54.60″ W, 667 m) | 0.18 | germacrene D (55.8), bicyclogermacrene (11.8), trans-caryophyllene (6.3) |
| Piper xylosteoides (Kunth.) Steud. | L52004/IAC 47075 | Ubatuba (45°7′37.64″ S, 23°25′16.03″ W, 30 m) | 1.04 | spathulenol (12.3), germacrene B (10.6), β-copaen-4-α-ol (9.4), trans-nerolidol (8.2), trans-β-guaiene; (7.8) |
| Rutaceae | ||||
| Esenbeckia febrifuga (A.St.-Hil.) A.Juss. ex Mart | Lp06091205/IAC 44591 | Campinas (47°4′3.49″ S, 22°51′47.19″ W, 672 m) | 0.14 | caryophyllene oxide (46.7) |
| Helietta apiculata Benth. | Lp6101823/IAC 47989 | Adamantina (51°9′12.19″ S, 21°39′41.90″ W, 365 m) | 0.16 | limonene (42.3) |
| Metrodorea nigra A.St.-Hil. | Lp06091204/IAC 46826 | Campinas (47°4′0.52″ S, 22°51′52.24″ W, 650 m) | 0.05 | spathulenol (23.6), bicyclogermacrene (16.6), germacrene D (15.3) |
| Zanthoxylum petiolare A.St.-Hil. & Tul. | Lp6101710/IAC 47962 | Votuporanga (50°3′29.80″ S, 20°27′27.40″ W, 479 m) | 0.18 | β-phellandrene (40.7), germacrene D (22.0) |
| Salicaceae | ||||
| Casearia sylvestris Sw. | R1643/IAC 46842 | Ubatuba (45°7′26.44″ S, 23°24′37.87″ W, 50 m) | 0.16 | trans-β-guaiene (12.2), 1,10-di-epi-cubenol (12.1) |
| Sapindaceae | ||||
| Cupania vernalis Cambess. | L4160/IAC 46969 | Campinas (47°4′1.63″ S, 22°51′47.24″ W, 670 m) | 0.20 | bicyclogermacrene (35.9), germacrene D (21.4), trans-caryophyllene (16.1) |
| Verbenaceae | ||||
| Aloysia virgata (Ruiz & Pav.) Juss. | C004/IAC 4614 | Jundiai (46°55′40.45″ S, 23°6′42.48″ W, 770 m) | 0.22 | γ-muurolene (32.7), trans-β-guaiene (24.6) |
| Microorganism | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Botanical Family | Bacteria | Filamentous Fungus | Yeast | ||||||
| Plant Species (Location) | Sample Code | C. xerosis | E. coli | P. acnes | P. aeruginosa | S. aureus | S. epidermidis | A. niger | C. albicans |
| Annonaceae | |||||||||
| Xylopia brasiliensis (Pariquera-Açu) | R1739 | 50 ± 2.2 | 50 ± 2.5 | 25 ± 1.3 | 25 ± 1.1 | 50 ± 2.5 | 50 ± 2.4 | 25 ± 1.2 | 25 ± 1.3 |
| Xylopia aromatica (Votuporanga) | Lp6101712 | 6.2 ± 0.1 | 25 ± 1.2 | 12.5 ± 0.6 | 25 ± 1.2 | 25 ± 1.3 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 |
| Lauraceae | |||||||||
| Nectandra megapotamica (Ribeirão Preto) | R1774 | 6.2 ± 0.1 | 12.5 ± 0.6 | 25 ± 1.2 | 25 ± 1.1 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.1 | 25 ± 1.2 |
| Myrtaceae | |||||||||
| Eugenia neoverrucosa (Campinas) | R1626 | 6.2 ± 0.1 | 25 ± 1.2 | 25 ±1.2 | 25 ± 1.2 | 25 ± 1.1 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 |
| Eugenia prasina (Ubatuba) | Lp51905 | 12.5 ± 0.6 | 25 ± 1.2 | 25 ± 1.2 | 12.5 ± 0.6 | 25 ± 1.1 | 25 ± 1.1 | 25 ± 1.2 | 25 ± 1.2 |
| Eugenia pyriformis (Jundiai) | C009 | 50 ± 2.3 | 50 ± 2.2 | 25 ± 1.2 | 25 ± 1.2 | 50 ± 2.2 | 50 ± 2.3 | 25 ± 1.2 | 25 ± 1.2 |
| Myrcia splendens (Jundiai) | C007 | 12.5 ± 0.6 | 25 ± 1.2 | 12.5 ± 0.6 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 |
| Rutaceae | |||||||||
| Helietta apiculata (Adamantina) | Lp6101823 | 12.5 ± 0.6 | 25 ± 1.2 | 12.5 ± 0.6 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 |
| Salicaceae | |||||||||
| Casearia sylvestris (Ubatuba) | R1643 | 12.5 ± 0.6 | 25 ± 1.2 | 12.5 ± 0.5 | 25 ± 1.2 | 25 ± 1.3 | 25 ± 1.2 | 25 ± 1.2 | 25 ± 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perigo, C.V.; Haber, L.L.; Facanali, R.; Vieira, M.A.R.; Torres, R.B.; Bernacci, L.C.; Guimarães, E.F.; Baitello, J.B.; Sobral, M.E.G.; Quecini, V.; et al. Essential Oils of Aromatic Plant Species from the Atlantic Rainforest Exhibit Extensive Chemical Diversity and Antimicrobial Activity. Antibiotics 2022, 11, 1844. https://doi.org/10.3390/antibiotics11121844
Perigo CV, Haber LL, Facanali R, Vieira MAR, Torres RB, Bernacci LC, Guimarães EF, Baitello JB, Sobral MEG, Quecini V, et al. Essential Oils of Aromatic Plant Species from the Atlantic Rainforest Exhibit Extensive Chemical Diversity and Antimicrobial Activity. Antibiotics. 2022; 11(12):1844. https://doi.org/10.3390/antibiotics11121844
Chicago/Turabian StylePerigo, Crislene V., Lenita L. Haber, Roselaine Facanali, Maria A. R. Vieira, Roseli B. Torres, Luís C. Bernacci, Elsie F. Guimarães, João B. Baitello, Marcos E. G. Sobral, Vera Quecini, and et al. 2022. "Essential Oils of Aromatic Plant Species from the Atlantic Rainforest Exhibit Extensive Chemical Diversity and Antimicrobial Activity" Antibiotics 11, no. 12: 1844. https://doi.org/10.3390/antibiotics11121844
APA StylePerigo, C. V., Haber, L. L., Facanali, R., Vieira, M. A. R., Torres, R. B., Bernacci, L. C., Guimarães, E. F., Baitello, J. B., Sobral, M. E. G., Quecini, V., & Marques, M. O. M. (2022). Essential Oils of Aromatic Plant Species from the Atlantic Rainforest Exhibit Extensive Chemical Diversity and Antimicrobial Activity. Antibiotics, 11(12), 1844. https://doi.org/10.3390/antibiotics11121844