
Studies on the Reactions of Biapenem with VIM Metallo β-Lactamases and the Serine β-Lactamase KPC-2

 ,
, 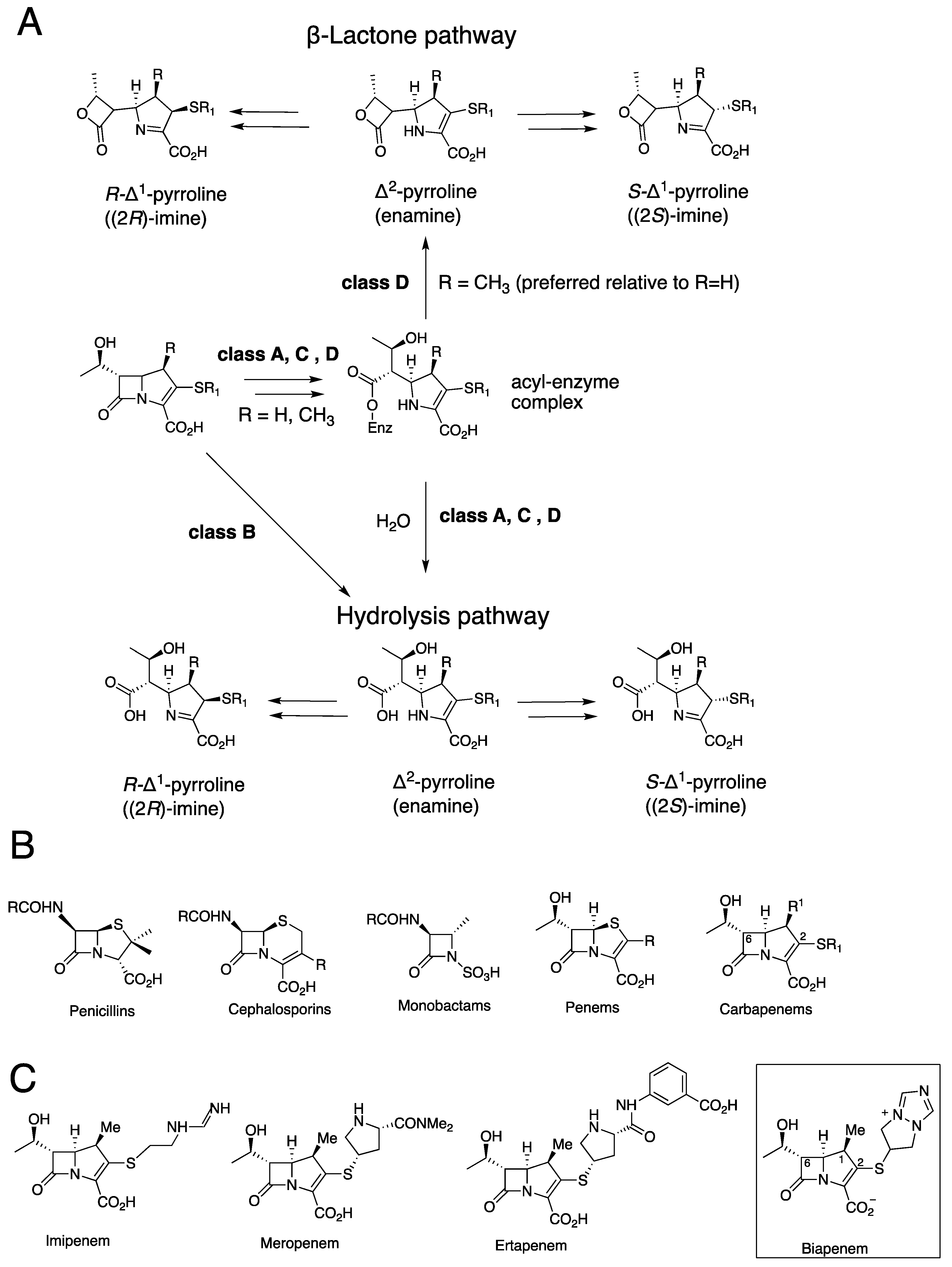{kind=link}
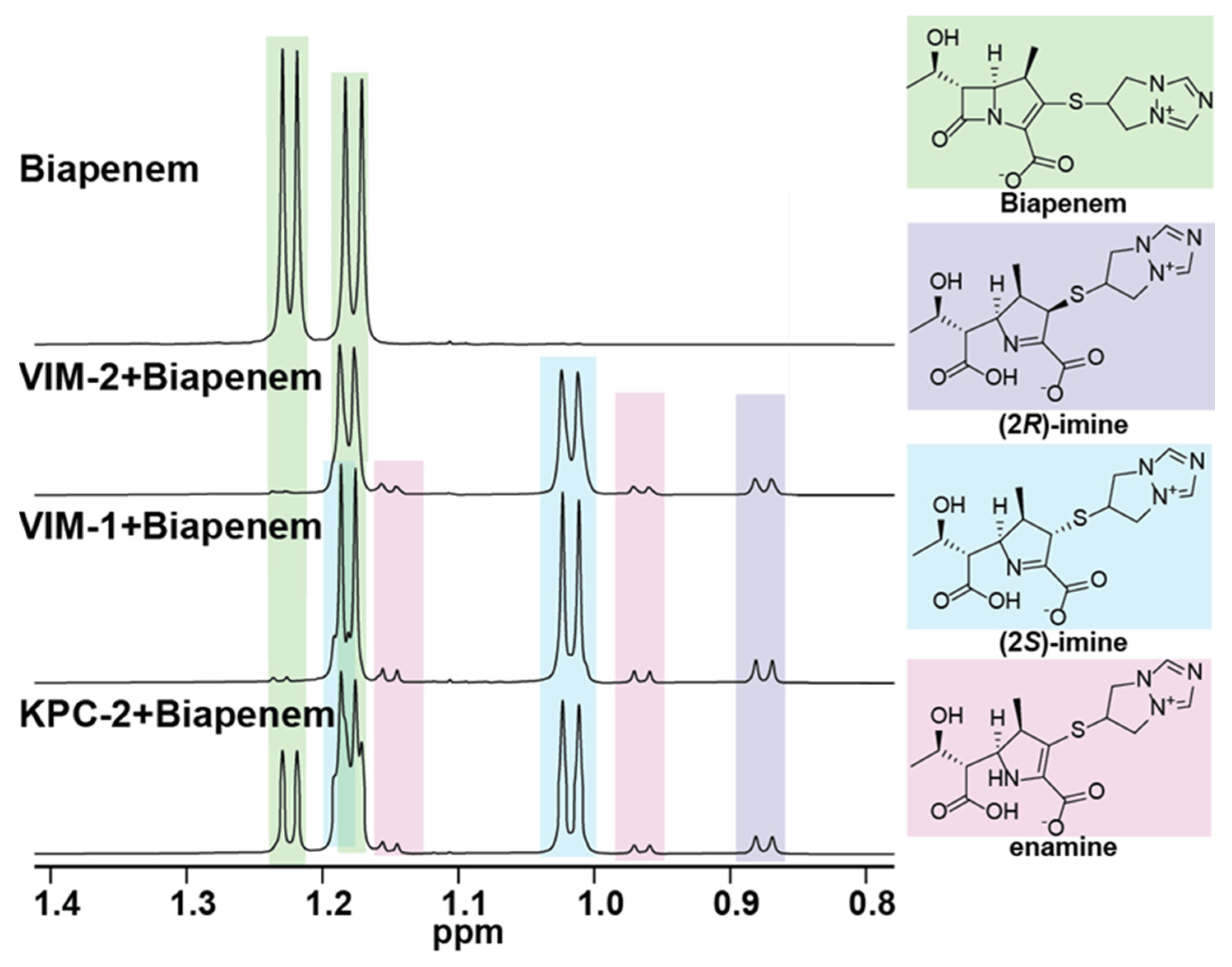{kind=link}
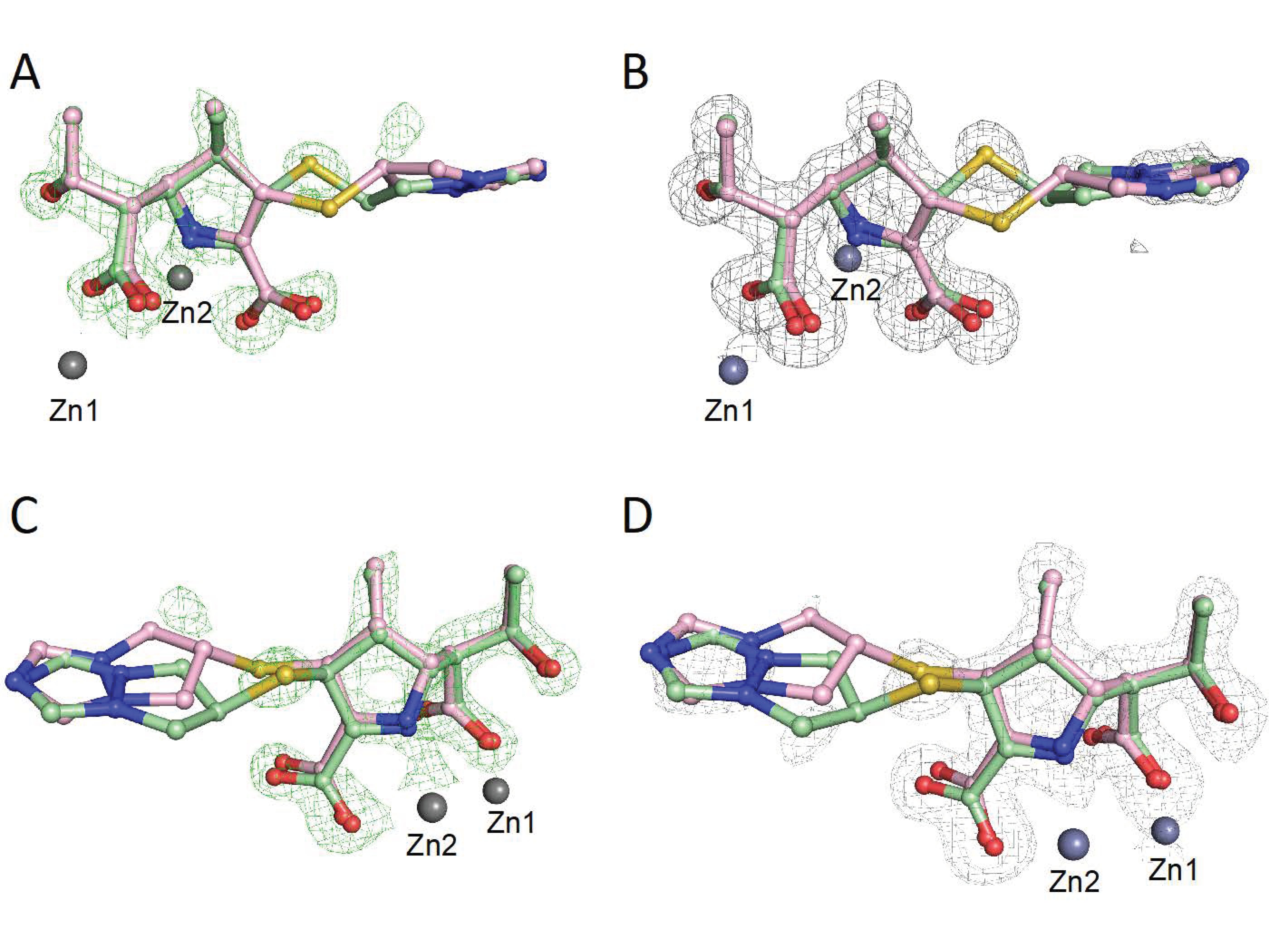{kind=link}
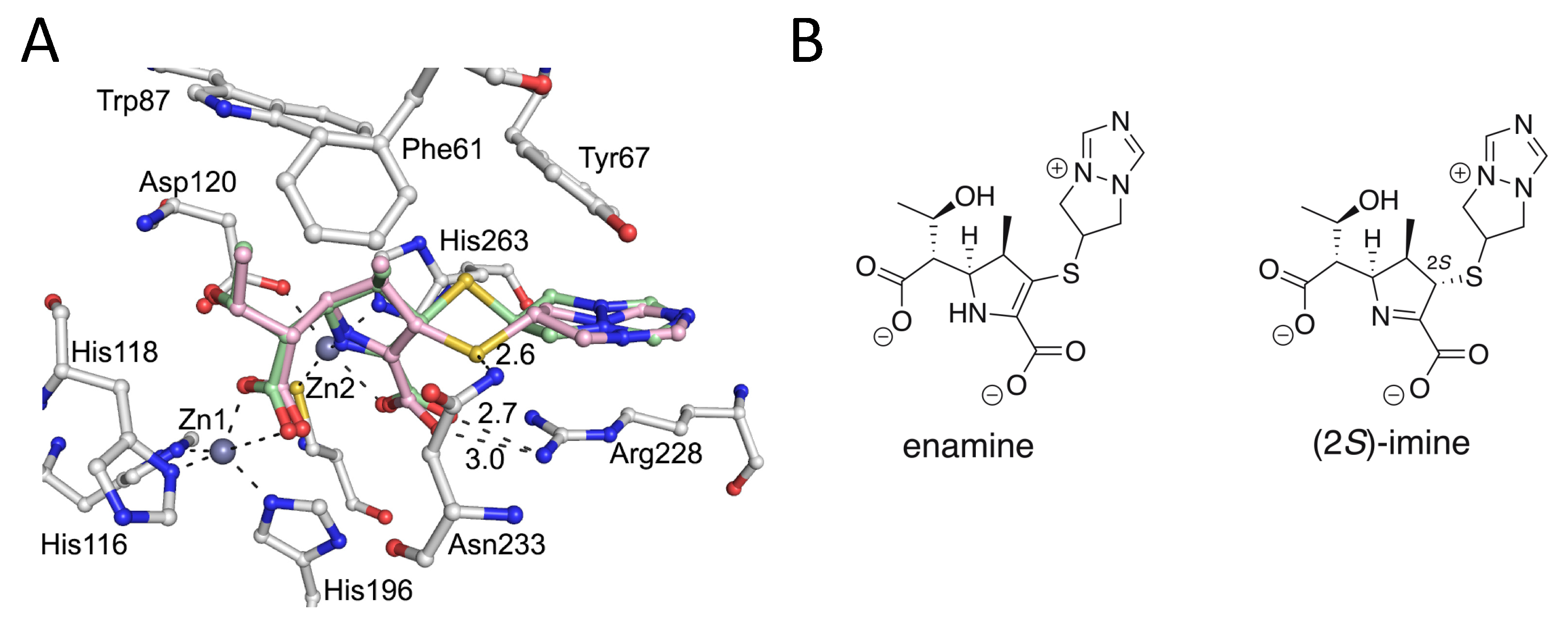{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Protein Purification and X-ray Crystallography
4.2. Data Collection and Processing
4.3. 1H NMR Studies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K. Bench-to-Bedside Review: The Role of Beta-Lactamases in Antibiotic-Resistant Gram-Negative Infections. Crit. Care. 2010, 14, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K. Metallo-Beta-Lactamases: A Class Apart. Clin. Infect. Dis. 1998, 27, S48–S53. [Google Scholar] [CrossRef] [Green Version]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum β-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meini, M.-R.; Llarrull, L.I.; Vila, A.J. Evolution of Metallo-β-lactamases: Trends Revealed by Natural Diversity and in vitro Evolution. Antibiotics 2014, 3, 285–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohans, C.T.; Van Groesen, E.; Kumar, K.; Tooke, C.L.; Spencer, J.; Paton, R.S.; Brem, J.; Schofield, C.J. A New Mechanism for β-Lactamases: Class D Enzymes Degrade 1β-Methyl Carbapenems through Lactone Formation. Angew. Chem. 2018, 130, 1296–1299. [Google Scholar] [CrossRef] [Green Version]
- Aertker, K.M.J.; Chan, H.T.H.; Lohans, C.T.; Schofield, C.J. Analysis of β-Lactone Formation by Clinically Observed Carbapenemases Informs on a Novel Antibiotic Resistance Mechanism. J. Biol. Chem. 2020, 295, 16604–16613. [Google Scholar] [CrossRef] [PubMed]
- Lohans, C.T.; Freeman, E.I.; van Groesen, E.; Tooke, C.L.; Hinchliffe, P.; Spencer, J.; Brem, J.; Schofield, C.J. Mechanistic Insights into β-Lactamase-Catalysed Carbapenem Degradation Through Product Characterisation. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- van Groesen, E.; Lohans, C.T.; Brem, J.; Aertker, K.M.; Claridge, T.D.W.; Schofield, C.J. 19F-NMR Monitoring of Reversible Protein Post-Translational Modifications: Class D Β-Lactamase Carbamylation and Inhibition. Chem. Eur. J. 2019, 25, 11837–11841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisa, M.-N.; Palacios, A.R.; Aitha, M.; González, M.M.; Moreno, D.M.; Crowder, M.W.; Bonomo, R.A.; Spencer, J.; Tierney, D.L.; Llarrull, L.I.; et al. A General Reaction Mechanism for Carbapenem Hydrolysis by Mononuclear and Binuclear Metallo-β-Lactamases. Nat. Comm. 2017, 8, 538. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, G.; Maccarone, G.; Mezzatesta, M.L.; Privitera, A.; Carciotto, V.; Santagati, M.; Stefani, S.; Nicoletti, G. In vitro Activity of Biapenem against Recent Gram-Negative and Gram-Positive Clinical Isolates. Chemotherapy 1997, 43, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Tamai, S.; Abe, T.; Matsunaga, H.; Hayashi, K.; Kishi, I.; Shiro, M.; Nagao, Y. New Straightforward Synthesis and Characterization of a Unique 1β-Methylcarbapenem Antibiotic Biapenem Bearing a σ-Symmetric Bicyclotriazoliumthio Group as the Pendant Moiety. J. Org. Chem. 1998, 63, 8145–8149. [Google Scholar] [CrossRef]
- Perry, C.M.; Ibbotson, T. Biapenem. Drugs 2002, 62, 2221–2234. [Google Scholar] [CrossRef] [PubMed]
- Lepsanovic, Z.; Libisch, B.; Tomanovic, B.; Nonkovic, Z.; Balogh, B.; Füzi, M. Characterisation of the First VIM Metallo- β -Lactamase-Producing Pseudomonas Aeruginosa Clinical Isolate in Serbia. Acta Microbiol. Immunol. Hung. 2008, 55, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Mavroidi, A.; Tsakris, A.; Tzelepi, E.; Pournaras, S.; Loukova, V.; Tzouvelekis, L.S. Carbapenem-hydrolysing VIM-2 metallo- beta-lactamase in Pseudomonas aeruginosa from Greece. J. Antimicrob. Chemother. 2000, 46, 1041–1043. [Google Scholar] [CrossRef] [Green Version]
- Docquier, J.-D.; Lamotte-Brasseur, J.; Galleni, M.; Amicosante, G.; Frère, J.-M.; Rossolini, G.M. On functional and structural heterogeneity of VIM-type metallo-beta-lactamases. J. Antimicrob. Chemother. 2003, 51, 257–266. [Google Scholar] [CrossRef]
- Makena, A.; Düzgün, A.Ö.; Brem, J.; Mcdonough, M.A.; Rydzik, A.M.; Abboud, M.I.; Saral, A.; Çiçek, Ç.A.; Sandalli, C.; Schofield, C.J. Comparison of Verona Integron-Borne Metallo-Lactamase (VIM) Variants Reveals Differences in Stability and Inhibition Profiles. Antimicrob. Agents Chemother. 2016, 60, 1377–1384. [Google Scholar] [CrossRef] [Green Version]
- Salimraj, R.; Hinchliffe, P.; Kosmopoulou, M.; Tyrrell, J.M.; Brem, J.; van Berkel, S.S.; Verma, A.; Owens, R.J.; McDonough, M.A.; Walsh, T.R.; et al. Crystal Structures of VIM-1 Complexes Explain Active Site Heterogeneity in VIM-Class Metallo-β-Lactamases. FEBS J. 2019, 286, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Lucic, A.; Hinchliffe, P.; Malla, T.R.; Tooke, C.L.; Brem, J.; Calvopiña, K.; Lohans, C.T.; Rabe, P.; McDonough, M.A.; Armistead, T.; et al. Faropenem Reacts with Serine and Metallo-β-Lactamases to Give Multiple Products. Eur. J. Med. Chem. 2021, 215, 1–10. [Google Scholar] [CrossRef]
- Mehta, S.C.; Furey, I.M.; Pemberton, O.A.; Boragine, D.M.; Chen, Y.; Palzkill, T. KPC-2 β-lactamase enables carbapenem antibiotic resistance through fast deacylation of the covalent intermediate. J. Biol. Chem. 2021, 296, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Galdadas, I.; Lovera, S.; Pérez-Hernández, G.; Barnes, M.D.; Healy, J.; Afsharikho, H.; Woodford, N.; Bonomo, R.A.; Gervasio, F.L.; Haider, S. Defining the Architecture of KPC-2 Carbapenemase: Identifying Allosteric Networks to Fight Antibiotics Resistance. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Garau, G.; Bebrone, C.; Anne, C.; Galleni, M.; Frère, J.-M.; Dideberg, O. A Metallo-β-lactamase Enzyme in Action: Crystal Structures of the Monozinc Carbapenemase CphA and its Complex with Biapenem. J. Mol. Biol. 2005, 345, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Watanabe, T.; Baba, N.; Takeuchi, Y.; Ohsawa, F.; Gomi, S. Crystal Structures of Biapenem and Tebipenem Complexed with Penicillin-Binding Proteins 2X and 1A from Streptococcus Pneumoniae. Antimicrob. Agents Chemother. 2008, 52, 2053–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchet, M.A.; Pan, Y.H.; Basta, L.A.B.; Saavedra, H.; Lloyd, E.P.; Kumar, P.; Mattoo, R.; Townsend, C.A.; Lamichhane, G. Structural insight into the inactivation of Mycobacterium tuberculosis non-classical transpeptidase LdtMt2 by biapenem and tebipenem. BMC Biochem. 2017, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Jin, W.; Matsunaga, K.; Ikemizu, S.; Yamagata, Y.; Wachino, J.; Shibata, N.; Arakawa, Y.; Kurosaki, H. Crystallographic Investigation of the Inhibition Mode of a VIM-2 Metallo-β-Lactamase from Pseudomonas Aeruginosa by a Mercaptocarboxylate Inhibitor. J. Med. Chem. 2007, 50, 6647–6653. [Google Scholar] [CrossRef] [PubMed]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y. Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef] [PubMed]
- Brem, J.; Panduwawala, T.; Hansen, J.U.; Hewitt, J.; Liepins, E.; Donets, P.; Espina, L.; Farley, A.J.M.; Shubin, K.; Campillos, G.G.; et al. Imitation of β-lactam binding enables broad-spectrum metallo-β-lactamase inhibitors. Nat. Chem. 2022, 14, 15–24. [Google Scholar] [CrossRef]
- Davies, D.T.; Leiris, S.; Sprynski, N.; Castandet, J.; Lozano, C.; Bousquet, J.; Zalacain, M.; Vasa, S.; Dasari, P.K.; Pattipati, R.; et al. ANT2681: SAR Studies Leading to the Identification of a Metallo-β-Lactamase Inhibitor with Potential for Clinical Use in Combination with Meropenem for the Treatment of Infections Caused by NDM-Producing Enterobacteriaceae. ACS Infect. Dis. 2020, 6, 2419–2430. [Google Scholar] [CrossRef]
- Cahill, S.; Tarhonskaya, H.; Rydzik, A.; Flashman, E.; McDonough, M.A.; Schofield, C.J.; Brem, J. Use of ferrous iron by metallo-β-lactamases. J. Inorg. Biochem. 2016, 163, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkova, A.; Lucic, A.; Krajnc, A.; Brem, J.; Calvopiña, K.; Langley, G.W.; McDonough, M.A.; Trapencieris, P.; Schofield, C.J. Broad Spectrum β-Lactamase Inhibition by a Thioether Substituted Bicyclic Boronate. ACS Infect. Dis. 2020, 6, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Waterman, D.G.; Parkhurst, J.M.; Brewster, A.S.; Gildea, R.J.; Gerstel, M.; Fuentes-Montero, L.; Vollmar, M.; Michels-Clark, T.; Young, I.; et al. DIALS: Implementation and evaluation of a new integration package. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brem, J.; van Berkel, S.S.; Zollman, D.; Lee, S.Y.; Gileadi, O.; McHugh, P.J.; Walsh, T.R.; McDonough, M.A.; Schofield, C.J. Structural Basis of Metallo-β-Lactamase Inhibition by Captopril Stereoisomers. Antimicrob. Agents Chemother. 2016, 60, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser Crystallographic Software. J. App. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A Comprehensive Python-Based System for Macromolecular Structure Solution. Acta Crystallogr. D. Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Struct. Biol. 2004, D60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Willcott, M.R. MestRe Nova. J. Am. Chem. Soc. 2009, 131, 13180. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucic, A.; Malla, T.R.; Calvopiña, K.; Tooke, C.L.; Brem, J.; McDonough, M.A.; Spencer, J.; Schofield, C.J. Studies on the Reactions of Biapenem with VIM Metallo β-Lactamases and the Serine β-Lactamase KPC-2. Antibiotics 2022, 11, 396. https://doi.org/10.3390/antibiotics11030396
Lucic A, Malla TR, Calvopiña K, Tooke CL, Brem J, McDonough MA, Spencer J, Schofield CJ. Studies on the Reactions of Biapenem with VIM Metallo β-Lactamases and the Serine β-Lactamase KPC-2. Antibiotics. 2022; 11(3):396. https://doi.org/10.3390/antibiotics11030396
Chicago/Turabian StyleLucic, Anka, Tika R. Malla, Karina Calvopiña, Catherine L. Tooke, Jürgen Brem, Michael A. McDonough, James Spencer, and Christopher J. Schofield. 2022. "Studies on the Reactions of Biapenem with VIM Metallo β-Lactamases and the Serine β-Lactamase KPC-2" Antibiotics 11, no. 3: 396. https://doi.org/10.3390/antibiotics11030396
APA StyleLucic, A., Malla, T. R., Calvopiña, K., Tooke, C. L., Brem, J., McDonough, M. A., Spencer, J., & Schofield, C. J. (2022). Studies on the Reactions of Biapenem with VIM Metallo β-Lactamases and the Serine β-Lactamase KPC-2. Antibiotics, 11(3), 396. https://doi.org/10.3390/antibiotics11030396