


Synergistic Inhibition of MRSA by Chenodeoxycholic Acid and Carbapenem Antibiotics

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Minimum Inhibitory Concentration (MIC) Assays
2.3. Checkerboard Synergy Assays
2.4. Time–Kill Curve Assay
2.5. Structured Illumination Microscopy (SIM)
2.6. Proteomic Analysis
3. Results
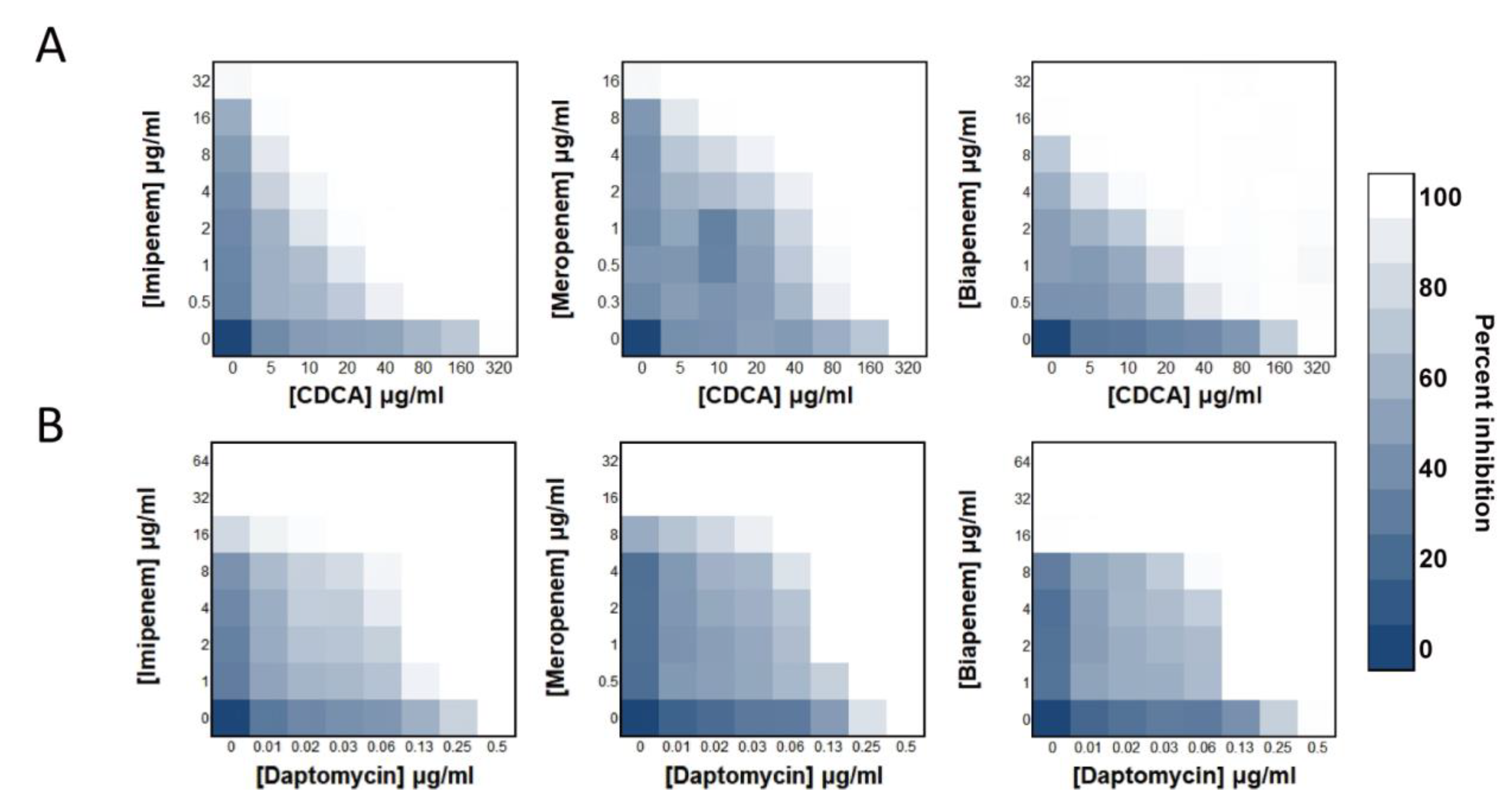
3.1. CDCA Sensitizes MRSA to Carbapenem Killing
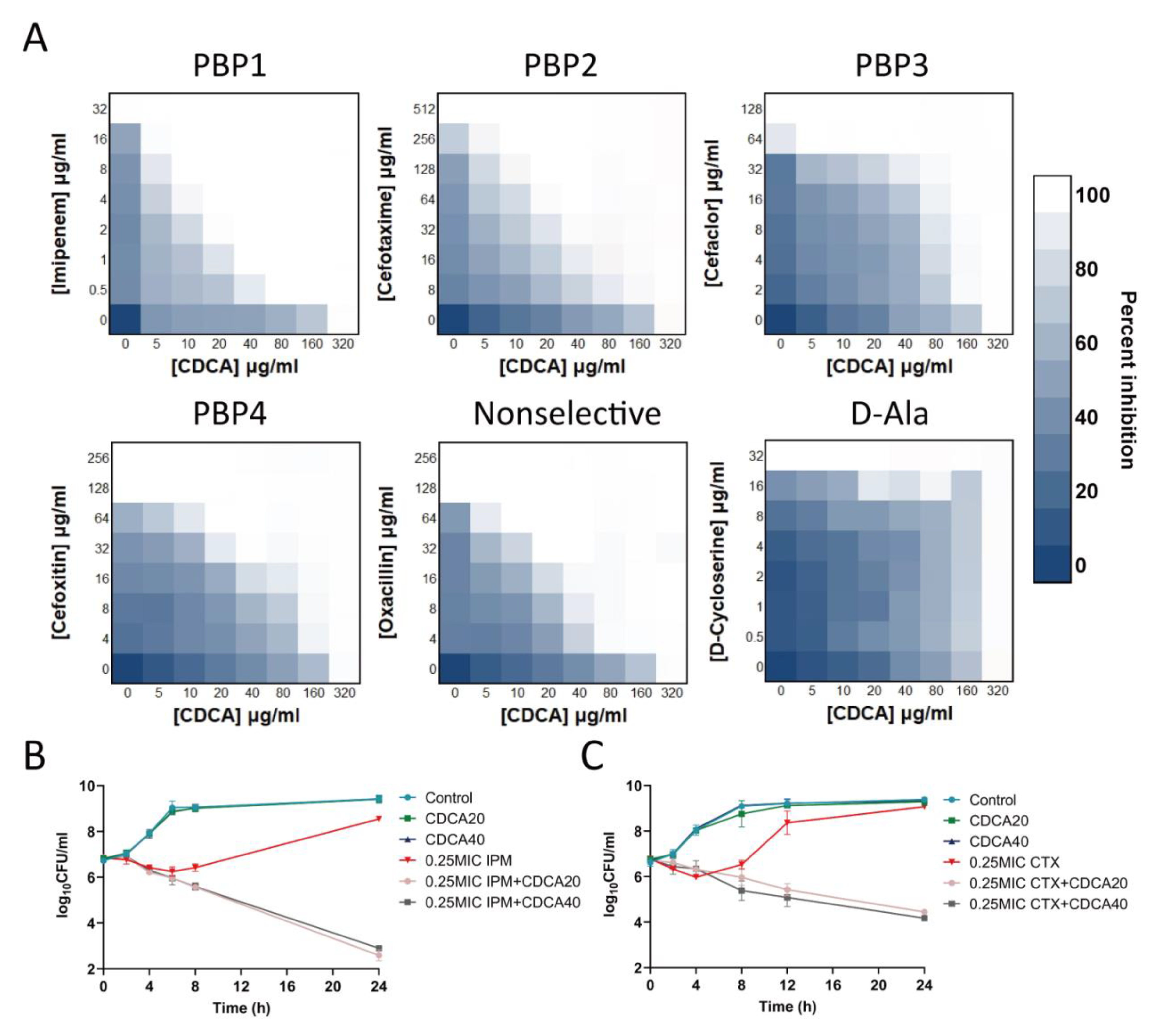
3.2. CDCA Improves the Bactericidal Effect of Antibiotics Targeting PBP1 and PBP2
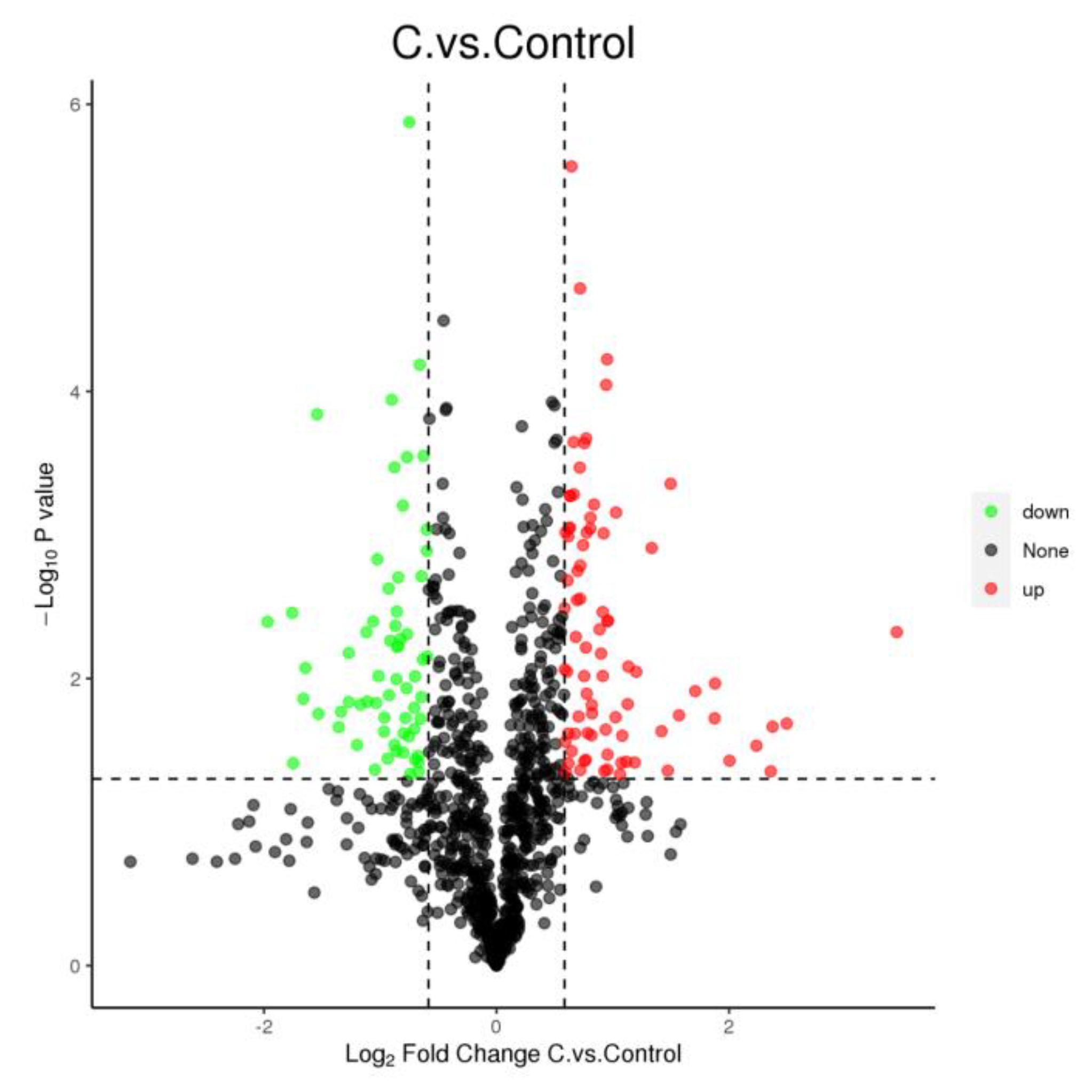
3.3. Proteome Analysis Revealed That CDCA Perturbed the Bacterial Cytoplasmic Membrane
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knox, J.; Uhlemann, A.C.; Lowy, F.D. Staphylococcus aureus infections: Transmission within households and the community. Trends Microbiol. 2015, 23, 437–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Boldock, E.; Surewaard, B.G.J.; Shamarina, D.; Na, M.; Fei, Y.; Ali, A.; Williams, A.; Pollitt, E.J.G.; Szkuta, P.; Morris, P.; et al. Human skin commensals augment Staphylococcus aureus pathogenesis. Nat. Microbiol. 2018, 3, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 2008, 46 (Suppl. S5), S350–S359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomer, L.; Schneewind, O.; Missiakas, D. Pathogenesis of Staphylococcus aureus bloodstream infections. Annu. Rev. Pathol. 2016, 11, 343–364. [Google Scholar] [CrossRef] [Green Version]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Peacock, S.J.; Paterson, G.K. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef]
- Kos, V.N.; Desjardins, C.A.; Griggs, A.; Cerqueira, G.; Van Tonder, A.; Holden, M.T.; Godfrey, P.; Palmer, K.L.; Bodi, K.; Mongodin, E.F.; et al. Comparative genomics of vancomycin-resistant Staphylococcus aureus strains and their positions within the clade most commonly associated with Methicillin-resistant S. aureus hospital-acquired infection in the United States. mBio 2012, 3, e00112-12. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical practice guidelines by the infectious diseases society of america for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: Executive summary. Clin. Infect. Dis. 2011, 52, 285–292. [Google Scholar] [CrossRef]
- Gould, I.M.; David, M.Z.; Esposito, S.; Garau, J.; Lina, G.; Mazzei, T.; Peters, G. New insights into meticillin-resistant Staphylococcus aureus (MRSA) pathogenesis, treatment and resistance. Int. J. Antimicrob. Agents 2012, 39, 96–104. [Google Scholar] [CrossRef]
- Rodvold, K.A.; McConeghy, K.W. Methicillin-resistant Staphylococcus aureus therapy: Past, present, and future. Clin. Infect. Dis. 2014, 58 (Suppl. S1), S20–S27. [Google Scholar] [CrossRef]
- Stryjewski, M.E.; Corey, G.R. Methicillin-resistant Staphylococcus aureus: An evolving pathogen. Clin. Infect. Dis. 2014, 58 (Suppl. S1), S10–S19. [Google Scholar] [CrossRef] [Green Version]
- El-Gamal, M.I.; Brahim, I.; Hisham, N.; Aladdin, R.; Mohammed, H.; Bahaaeldin, A. Recent updates of carbapenem antibiotics. Eur. J. Med. Chem. 2017, 131, 185–195. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [Green Version]
- Garidel, P.; Hildebrand, A.; Knauf, K.; Blume, A. Membranolytic activity of bile salts: Influence of biological membrane properties and composition. Molecules 2007, 12, 2292–2326. [Google Scholar] [CrossRef] [Green Version]
- Patra, D.; Ahmadieh, D.; Aridi, R. Study on interaction of bile salts with curcumin and curcumin embedded in dipalmitoyl-sn-glycero-3-phosphocholine liposome. Colloids Surf. B Biointerfaces 2013, 110, 296–304. [Google Scholar] [CrossRef]
- Moghimipour, E.; Ameri, A.; Handali, S. Absorption-enhancing effects of bile salts. Molecules 2015, 20, 14451–14473. [Google Scholar] [CrossRef] [Green Version]
- Moghimipour, E.; Tabassi, S.A.; Ramezani, M.; Handali, S.; Lobenberg, R. Brush border membrane vesicle and Caco-2 cell line: Two experimental models for evaluation of absorption enhancing effects of saponins, bile salts, and some synthetic surfactants. J. Adv. Pharm. Technol. Res. 2016, 7, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Cui, K.; Tong, Q.; Ma, S.; Sun, Y.; He, G.; Li, D.; Lin, L.; Blazekovic, B.; Chevalier, S.; et al. Traditional Chinese medicine Tanreqing targets both cell division and virulence in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2022, 12, 884045. [Google Scholar] [CrossRef]
- Yang, W.; Wei, Q.; Tong, Q.; Cui, K.; He, G.; Lin, L.; Ma, L.Z.; Cornelis, P.; Wang, Y. Traditional Chinese medicine Tanreqing inhibits quorum sensing systems in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 517462. [Google Scholar] [CrossRef]
- Yang, W.; Liu, J.; Blazekovic, B.; Sun, Y.; Ma, S.; Ren, C.; Vladimir-Knezevic, S.; Li, C.; Xing, Y.; Tian, G.; et al. In vitro antibacterial effects of Tanreqing injection combined with vancomycin or linezolid against methicillin-resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2018, 18, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, T.; Hu, J.; Ren, C.; Lei, H.; Hou, Y.; Brantner, A.H. Anti-biofilm activity of TanReQing, a Traditional Chinese Medicine used for the treatment of acute pneumonia. J. Ethnopharmacol. 2011, 134, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Jian, C.; Peng, Q.; Hou, T.; Wu, K.; Shang, B.; Zhao, M.; Wang, Y.; Zheng, W.; Ma, Q.; et al. Prohibitin 2 deficiency impairs cardiac fatty acid oxidation and causes heart failure. Cell Death Dis. 2020, 11, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panesso, D.; Planet, P.J.; Diaz, L.; Hugonnet, J.E.; Tran, T.T.; Narechania, A.; Munita, J.M.; Rincon, S.; Carvajal, L.P.; Reyes, J.; et al. Methicillin-susceptible, vancomycin-resistant Staphylococcus aureus, Brazil. Emerg. Infect. Dis. 2015, 21, 1844–1848. [Google Scholar] [CrossRef] [PubMed]
- Assis, L.R.; Theodoro, R.D.S.; Costa, M.B.S.; Nascentes, J.A.S.; Rocha, M.D.D.; Bessa, M.A.S.; Menezes, R.P.; Dilarri, G.; Hypolito, G.B.; Santos, V.R.D.; et al. Antibacterial activity of isobavachalcone (IBC) is associated with membrane disruption. Membranes 2022, 12, 269. [Google Scholar] [CrossRef]
- Munita, J.M.; Bayer, A.S.; Arias, C.A. Evolving resistance among Gram-positive pathogens. Clin. Infect. Dis. 2015, 61 (Suppl. S2), S48–S57. [Google Scholar] [CrossRef] [Green Version]
- Berti, A.D.; Theisen, E.; Sauer, J.D.; Nonejuie, P.; Olson, J.; Pogliano, J.; Sakoulas, G.; Nizet, V.; Proctor, R.A.; Rose, W.E. Penicillin binding protein 1 is important in the compensatory response of Staphylococcus aureus to daptomycin-induced membrane damage and is a potential target for beta-lactam-daptomycin synergy. Antimicrob. Agents Chemother. 2016, 60, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.F.; Henriques, A.O.; Pinho, M.G.; de Lencastre, H.; Tomasz, A. Role of PBP1 in cell division of Staphylococcus aureus. J. Bacteriol. 2007, 189, 3525–3531. [Google Scholar] [CrossRef] [Green Version]
- Berti, A.D.; Sakoulas, G.; Nizet, V.; Tewhey, R.; Rose, W.E. beta-lactam antibiotics targeting PBP1 selectively enhance daptomycin activity against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.F.; Henriques, A.O.; Pinho, M.G.; de Lencastre, H.; Tomasz, A. Evidence for a dual role of PBP1 in the cell division and cell separation of Staphylococcus aureus. Mol. Microbiol. 2009, 72, 895–904. [Google Scholar] [CrossRef]
- Reed, P.; Atilano, M.L.; Alves, R.; Hoiczyk, E.; Sher, X.; Reichmann, N.T.; Pereira, P.M.; Roemer, T.; Filipe, S.R.; Pereira-Leal, J.B.; et al. Staphylococcus aureus survives with a minimal peptidoglycan synthesis machine but sacrifices virulence and antibiotic resistance. PLoS Pathog. 2015, 11, e1004891. [Google Scholar] [CrossRef] [Green Version]
- Pinho, M.G.; de Lencastre, H.; Tomasz, A. An acquired and a native penicillin-binding protein cooperate in building the cell wall of drug-resistant staphylococci. Proc. Natl. Acad. Sci. USA 2001, 98, 10886–10891. [Google Scholar] [CrossRef] [Green Version]
- Lutkenhaus, J.; Pichoff, S.; Du, S. Bacterial cytokinesis: From Z ring to divisome. Cytoskeleton 2012, 69, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.F.; Mobashery, S. β-Lactams against the fortress of the Gram-positive Staphylococcus aureus bacterium. Chem. Rev. 2021, 121, 3412–3463. [Google Scholar] [CrossRef]
- Garcia-Fernandez, E.; Koch, G.; Wagner, R.M.; Fekete, A.; Stengel, S.T.; Schneider, J.; Mielich-Suss, B.; Geibel, S.; Markert, S.M.; Stigloher, C.; et al. Membrane microdomain disassembly inhibits MRSA antibiotic resistance. Cell 2017, 171, 1354–1367. [Google Scholar] [CrossRef]
- Pereira, A.R.; Reed, P.; Veiga, H.; Pinho, M.G. The Holliday junction resolvase RecU is required for chromosome segregation and DNA damage repair in Staphylococcus aureus. BMC Microbiol. 2013, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Janion, C. Inducible SOS response system of DNA repair and mutagenesis in Escherichia coli. Int. J. Biol. Sci. 2008, 4, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Bojer, M.S.; Wacnik, K.; Kjelgaard, P.; Gallay, C.; Bottomley, A.L.; Cohn, M.T.; Lindahl, G.; Frees, D.; Veening, J.W.; Foster, S.J.; et al. SosA inhibits cell division in Staphylococcus aureus in response to DNA damage. Mol. Microbiol. 2019, 112, 1116–1130. [Google Scholar] [CrossRef] [Green Version]
- Bojer, M.S.; Frees, D.; Ingmer, H. SosA in Staphylococci: An addition to the paradigm of membrane-localized, SOS-induced cell division inhibition in bacteria. Curr. Genet. 2020, 66, 495–499. [Google Scholar] [CrossRef]
- Ortwine, J.K.; Werth, B.J.; Sakoulas, G.; Rybak, M.J. Reduced glycopeptide and lipopeptide susceptibility in Staphylococcus aureus and the “seesaw effect”: Taking advantage of the back door left open? Drug Resist. Updates 2013, 16, 73–79. [Google Scholar] [CrossRef]
- Renzoni, A.; Kelley, W.L.; Rosato, R.R.; Martinez, M.P.; Roch, M.; Fatouraei, M.; Haeusser, D.P.; Margolin, W.; Fenn, S.; Turner, R.D.; et al. Molecular bases determining daptomycin resistance-mediated resensitization to beta-Lactams (Seesaw Effect) in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2017, 61, e01634-16. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FICI | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.09375 | 0.125 | 0.1875 | 0.25 | 0.3125 | 0.375 | 0.5 | 0.625 | 0.75 | |
| IPM | 1 | 1 | 6 | 4 | 1 | 5 | 0 | 1 | 6 |
| MEM | 0 | 0 | 2 | 7 | 3 | 5 | 1 | 2 | 5 |
| BPM | 0 | 1 | 5 | 7 | 1 | 3 | 1 | 0 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, K.; Yang, W.; Liu, S.; Li, D.; Li, L.; Ren, X.; Sun, Y.; He, G.; Ma, S.; Zhang, J.; et al. Synergistic Inhibition of MRSA by Chenodeoxycholic Acid and Carbapenem Antibiotics. Antibiotics 2023, 12, 71. https://doi.org/10.3390/antibiotics12010071
Cui K, Yang W, Liu S, Li D, Li L, Ren X, Sun Y, He G, Ma S, Zhang J, et al. Synergistic Inhibition of MRSA by Chenodeoxycholic Acid and Carbapenem Antibiotics. Antibiotics. 2023; 12(1):71. https://doi.org/10.3390/antibiotics12010071
Chicago/Turabian StyleCui, Kaiyu, Weifeng Yang, Shuang Liu, Dongying Li, Lu Li, Xing Ren, Yanan Sun, Gaiying He, Shuhua Ma, Jidan Zhang, and et al. 2023. "Synergistic Inhibition of MRSA by Chenodeoxycholic Acid and Carbapenem Antibiotics" Antibiotics 12, no. 1: 71. https://doi.org/10.3390/antibiotics12010071
APA StyleCui, K., Yang, W., Liu, S., Li, D., Li, L., Ren, X., Sun, Y., He, G., Ma, S., Zhang, J., Wei, Q., & Wang, Y. (2023). Synergistic Inhibition of MRSA by Chenodeoxycholic Acid and Carbapenem Antibiotics. Antibiotics, 12(1), 71. https://doi.org/10.3390/antibiotics12010071