
Performance of Two Extracts Derived from Propolis on Mature Biofilm Produced by Candida albicans

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
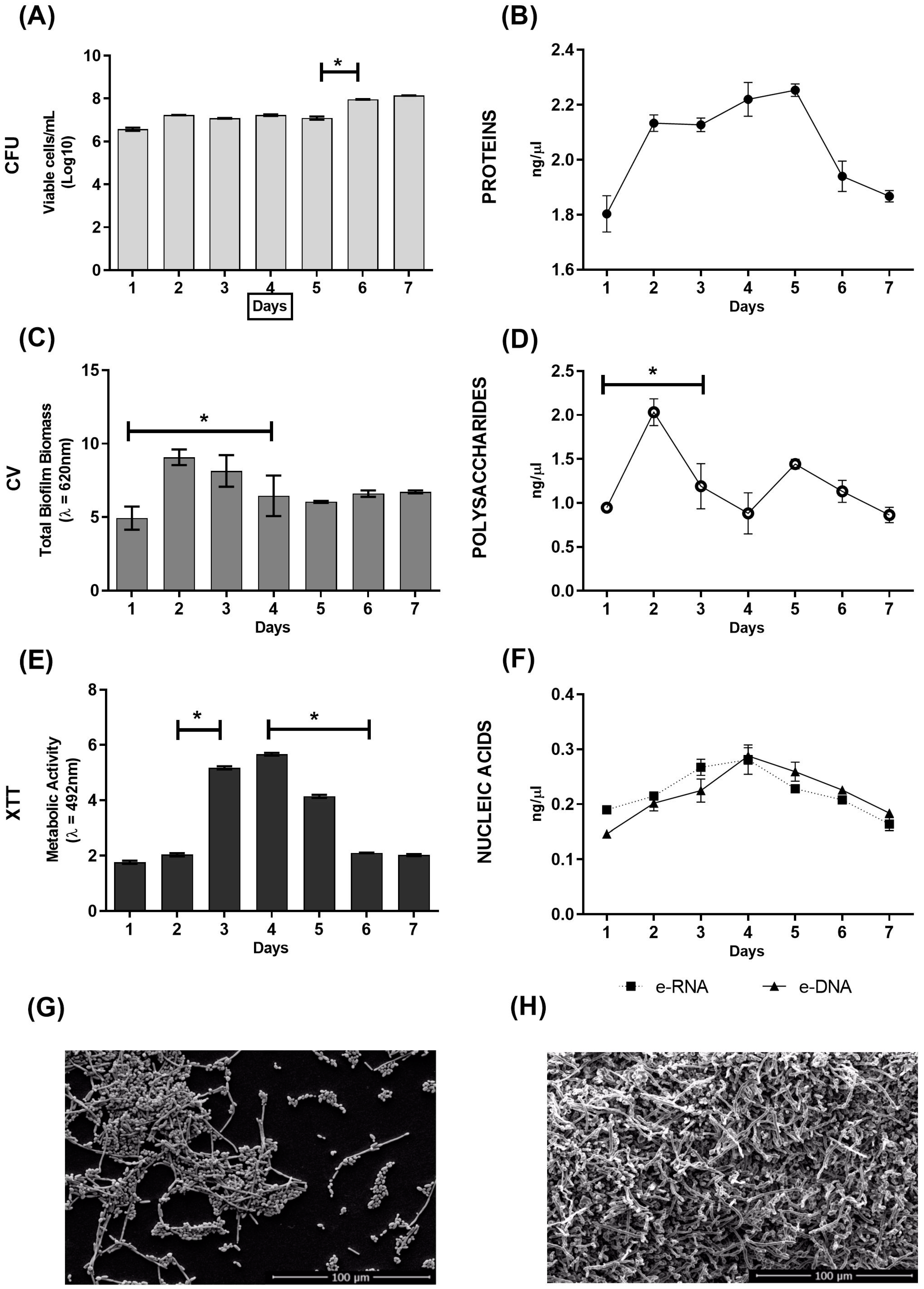
2.1. In Vitro Evaluation of C. albicans Biofilm Formation over Seven Days
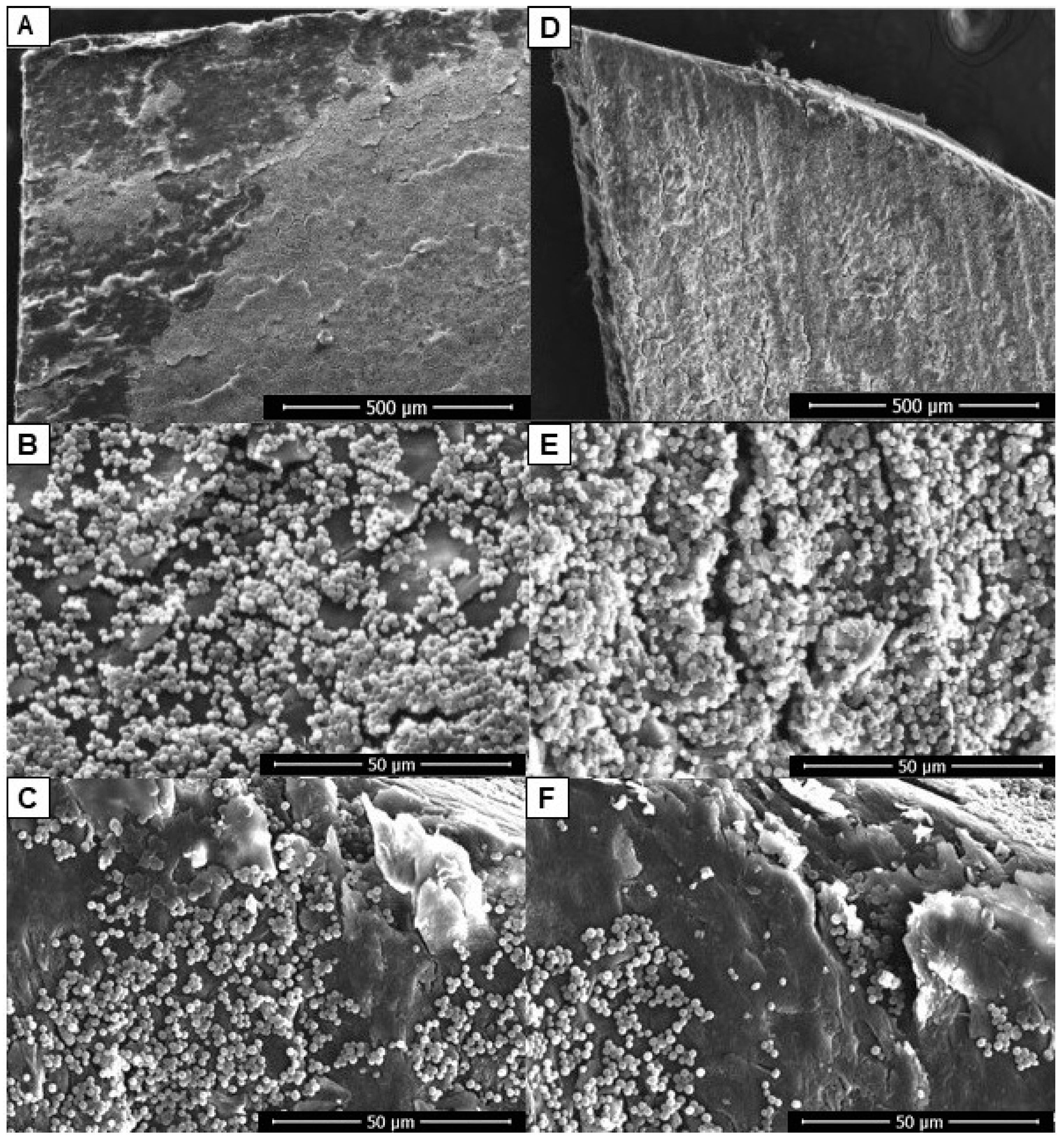
2.2. Assessment of the Ability of C. albicans to Form Biofilm in an Ex Vivo Model of Onychomycosis
2.3. Antibiofilm Activity of PE and WPE against C. albicans
2.4. Cytotoxicity of PE and WPE
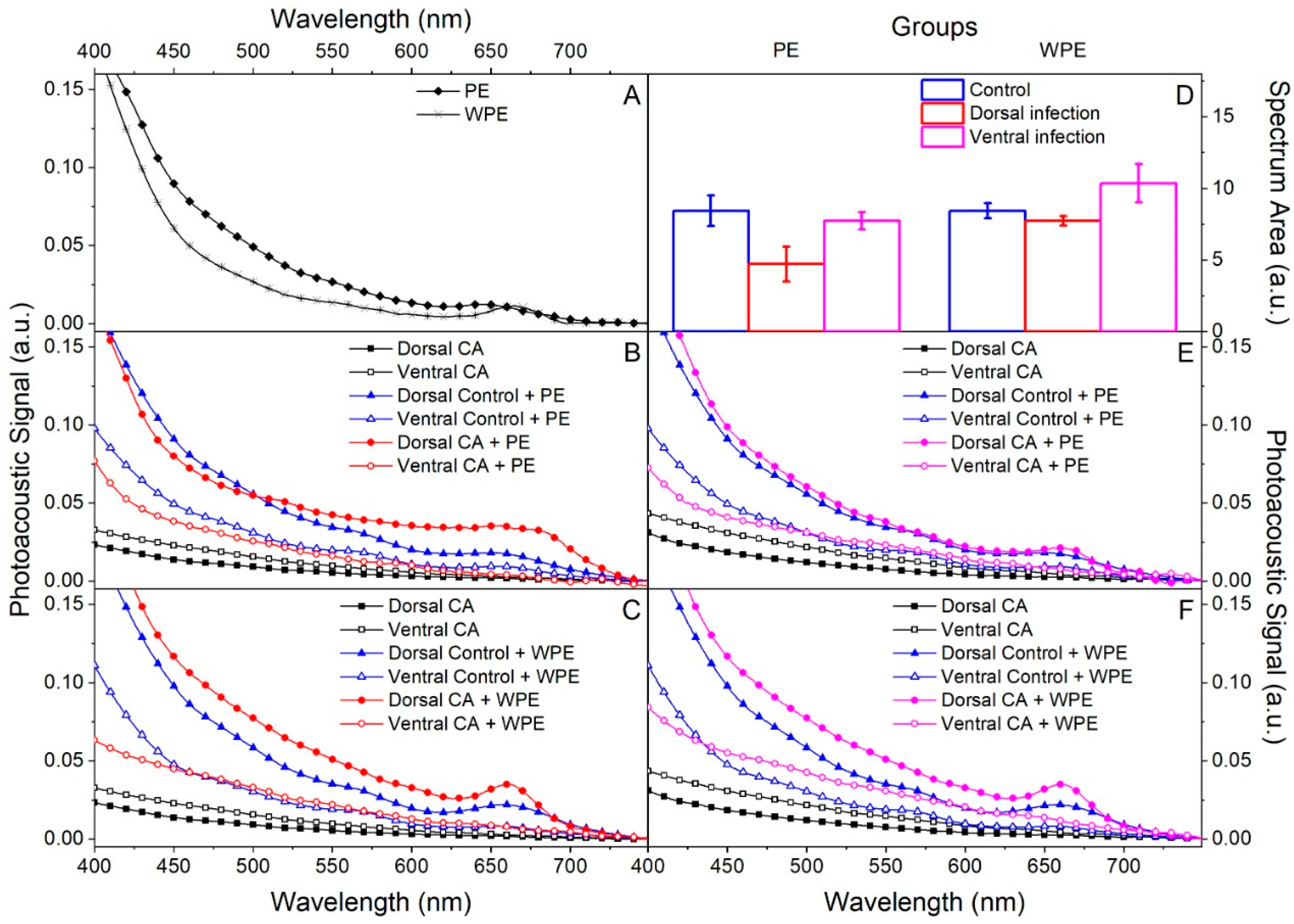
2.5. Permeation Capacity of Both Extracts in an Ex Vivo Model of Onychomycosis Caused by C. albicans
3. Discussion
4. Materials and Methods
4.1. Fungal Strain
4.2. Biofilm Formation in Culture Microplates
4.3. Determination of the Number of Viable Cells in Biofilm
4.4. Biofilm Biomass Quantification by Crystal Violet Staining
4.5. Evaluation of Metabolic Activity by XTT Assay
4.6. Characterization of Biofilm
4.7. Propolis and By-Product Extracts
4.8. Antibiofilm Effect of Propolis and By-Product Extracts
4.9. Effect of Propolis and By-Product Extracts on Mammalian Cells
4.10. Biofilm Formation and Permeance Analysis of Propolis and Propolis By-Product Extracts in Human Nails
4.10.1. Human Nails
4.10.2. Fourier Transform Infrared-Attenuated Total Reflectance (FTIR-ATR) Spectroscopy
4.10.3. Photoacoustic Spectroscopy (PAS)
4.11. Scanning Electron Microscopy (SEM) Analyses
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, A.K.; Stec, N.; Summerbell, R.C.; Shear, N.H.; Piguet, V.; Tosti, A.; Piraccini, B.M. Onychomycosis: A review. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1972–1990. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.; Lam, J.M.; Leong, K.F.; Hon, K.L.; Barankin, B.; Leung, A.A.; Wong, A.H. Onychomycosis: An Updated Review. Recent Patents Inflamm. Allergy Drug Discov. 2020, 14, 32–45. [Google Scholar] [CrossRef]
- Gupta, A.K.; Versteeg, S.G.; Shear, N.H. Onychomycosis in the 21st Century: An Update on Diagnosis, Epidemiology, and Treatment. J. Cutan. Med. Surg. 2017, 21, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Widaty, S.; Miranda, E.; Bramono, K.; Menaldi, S.L.; Marissa, M.; Oktarina, C.; Surya, D.; Kusumawardhani, H.T. Prognostic factors influencing the treatment outcome of onychomycosis Candida. Mycoses 2020, 63, 71–77. [Google Scholar] [CrossRef]
- Castillo, C.; Masi, M.F.; Mishko, A.; Sheltzer, A.; Speer, A.; Tran, H.; Vlahovic, T.C. Biofilms and the Nail Unit. Clin. Podiatr. Med. Surg. 2021, 38, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Daigle, D.; Carviel, J.L. The role of biofilms in onychomycosis. J. Am. Acad. Dermatol. 2016, 74, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; A Foley, K. Evidence for biofilms in onychomycosis. G. Ital. di Dermatol. e Venereol. 2019, 154, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Veiga, F.F.; de Castro-Hoshino, L.V.; Rezende, P.S.T.; Baesso, M.L.; Svidzinski, T.I.E. Insights on the etiopathogenesis of onychomycosis by dermatophyte, yeast and non-dermatophyte mould in ex vivo model. Exp. Dermatol. 2022, 31, 1810–1814. [Google Scholar] [CrossRef]
- Dota, K.F.D.; Consolaro, M.E.L.; Svidzinski, T.I.E.; Bruschi, M.L. Antifungal Activity of Brazilian Propolis Microparticles against Yeasts Isolated from Vulvovaginal Candidiasis. Evid.-Based Complement. Altern. Med. 2011, 2011, 201953. [Google Scholar] [CrossRef] [Green Version]
- Pereira, R.R.D.A.; Godoy, J.S.R.; Svidzinski, T.I.S.; Bruschi, M.L. Preparation and Characterization of Mucoadhesive Thermoresponsive Systems Containing Propolis for the Treatment of Vulvovaginal Candidiasis. J. Pharm. Sci. 2013, 102, 1222–1234. [Google Scholar] [CrossRef]
- Capoci, I.R.G.; Bonfim-Mendonça, P.D.S.; Arita, G.S.; Pereira, R.R.D.A.; Consolaro, M.E.L.; Bruschi, M.L.; Negri, M.; Svidzinski, T.I.E. Propolis Is an Efficient Fungicide and Inhibitor of Biofilm Production by Vaginal Candida albicans. Evid.-Based Complement. Altern. Med. 2015, 2015, 287693. [Google Scholar] [CrossRef] [Green Version]
- Sayyadi, F.; Mahdavi, S.; Moghadamnia, A.A.; Moslemi, D.; Shirzad, A.; Motallebnejad, M. The effect of aqueous and ethanolic extract of Iranian propolis on Candida albicans isolated from the mouth of patients with colorectal malignancy undergone chemotherapy: An in-vitro study. Casp. J. Intern. Med. 2020, 11, 62–66. [Google Scholar] [CrossRef]
- Siqueira, A.B.S.; Rodriguez, L.R.N.D.A.; Santos, R.K.B.; Marinho, R.R.B.; Abreu, S.; Peixoto, R.F.; Gurgel, B.C.D.V. Antifungal activity of propolis against Candida species isolated from cases of chronic periodontitis. Braz. Oral Res. 2015, 29, 1–6. [Google Scholar] [CrossRef]
- Bonfim, A.P.; Sakita, K.M.; Faria, D.R.; Arita, G.S.; Vendramini, F.A.V.R.; Capoci, I.R.G.; Braga, A.G.; dos Santos, R.S.; Bruschi, M.L.; Becker, T.C.A.; et al. Preclinical approaches in vulvovaginal candidiasis treatment with mucoadhesive thermoresponsive systems containing propolis. PLoS ONE 2020, 15, e0243197. [Google Scholar] [CrossRef]
- Moghim, H.; Taghipour, S.; Kheiri, S.; Khabbazi, H.; Baradaran, A. Antifungal Effects of Iranian Propolis Extract and Royal jelly Against Candida albicans In-Vitro. Int. J. Prev. Med. 2021, 12, 163. [Google Scholar] [CrossRef]
- Oliveira, A.C.P.; Shinobu, C.S.; Longhini, R.; Franco, S.L.; Svidzinski, T.I.E. Antifungal activity of propolis extract against yeasts isolated from onychomycosis lesions. Mem. Inst. Oswaldo Cruz. 2006, 101, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, A.R.; Shokri, H.; Nikaein, D.; Mansouri, P.; Erfanmanesh, A.; Chalangari, R.; Katalin, M. Yeasts as Important Agents of Onychomycosis: In Vitro Activity of Propolis Against Yeasts Isolated from Patients with Nail Infection. J. Altern. Complement. Med. 2013, 19, 57–62. [Google Scholar] [CrossRef]
- Barros, I.L.; Veiga, F.F.; Jarros, I.C.; Corrêa, J.L.; DOS Santos, R.S.; Bruschi, M.L.; Negri, M.; Svidzinski, T.I. Promising effect of propolis and a by-product on planktonic cells and biofilm formation by the main agents of human fungal infections. Acad. Bras. Cienc. 2022, 94, e20210189. [Google Scholar] [CrossRef]
- Pierce, C.G.; Vila, T.; Romo, J.A.; Montelongo-Jauregui, D.; Wall, G.; Ramasubramanian, A.; Lopez-Ribot, J.L. The Candida albicans Biofilm Matrix: Composition, Structure and Function. J. Fungi 2017, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Zarnowski, R.; Westler, W.M.; Lacmbouh, G.A.; Marita, J.M.; Bothe, J.R.; Bernhardt, J.; Sahraoui, A.L.-H.; Fontaine, J.; Sanchez, H.; Hatfield, R.D.; et al. Novel Entries in a Fungal Biofilm Matrix Encyclopedia. mBio 2014, 5, e01333-14. [Google Scholar] [CrossRef]
- Pereira, R.; Fontenelle, R.S.; Brito, E.; Morais, S.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef]
- Ramage, G.; Vandewalle, K.; Wickes, B.L.; López-Ribot, J.L. Characteristics of biofilm formation by Candida albicans. Rev. Iberoam. Micol 2001, 18, 163–170. [Google Scholar]
- Jayatilake, J.A.M.S.; Tilakaratne, W.M.; Panagoda, G.J. Candidal onychomycosis: A Mini-Review. Mycopathologia 2009, 168, 165–173. [Google Scholar] [CrossRef]
- Mohammadi, F.; Ghasemi, Z.; Familsatarian, B.; Salehi, E.; Sharifynia, S.; Barikani, A.; Mirzadeh, M.; Hosseini, M.A. Relationship between antifungal susceptibility profile and virulence factors in Candida albicans isolated from nail specimens. Rev. Soc. Bras. Med. Trop. 2020, 53, e20190214. [Google Scholar] [CrossRef]
- Baswan, S.; Kasting, G.B.; Li, S.K.; Wickett, R.; Adams, B.; Eurich, S.; Schamper, R. Understanding the formidable nail barrier: A review of the nail microstructure, composition and diseases. Mycoses 2017, 60, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Adt, I.; Toubas, D.; Pinon, J.-M.; Manfait, M.; Sockalingum, G.D. FTIR spectroscopy as a potential tool to analyse structural modifications during morphogenesis of Candida albicans. Arch. Microbiol. 2006, 185, 277–285. [Google Scholar] [CrossRef]
- Shi, Q.-Y.; Câmara, C.R.S.; Schlegel, V. Biochemical alterations of Candida albicans during the phenotypic transition from yeast to hyphae captured by Fourier transform mid-infrared-attenuated reflectance spectroscopy. Anal. 2018, 143, 5404–5416. [Google Scholar] [CrossRef]
- Veiga, F.F.; de Castro-Hoshino, L.V.; Sato, F.; Bombassaro, A.; A Vicente, V.; Mendes, V.; Baesso, M.L.; Negri, M.; Svidzinski, T.I. Fusarium oxysporum is an onychomycosis etiopathogenic agent. Futur. Microbiol. 2018, 13, 1745–1756. [Google Scholar] [CrossRef]
- Veiga, F.F.; de Castro-Hoshino, L.V.; Sato, F.; Baesso, M.L.; Silva, S.; Negri, M.; Svidzinski, T.I.E. Characterization of a biofilm formed by Fusarium oxysporum on the human nails. Int. J. Dermatol. 2021, 61, 191–198. [Google Scholar] [CrossRef]
- Freires, I.; Queiroz, V.; Furletti, V.; Ikegaki, M.; de Alencar, S.; Duarte, M.; Rosalen, P. Chemical composition and antifungal potential of Brazilian propolis against Candida spp. J. Med Mycol. 2016, 26, 122–132. [Google Scholar] [CrossRef]
- Tobaldini-Valerio, F.K.; Bonfim-Mendonça, P.S.; Rosseto, H.C.; Bruschi, M.L.; Henriques, M.; Negri, M.; Silva, S.C.; Svidzinski, T.I.E. Propolis: A potential natural product to fight Candida species infections. Future Microbiol. 2016, 11, 1035–1046. [Google Scholar] [CrossRef]
- Martorano-Fernandes, L.; Cavalcanti, Y.W.; Almeida, L.D.F.D.D. “Inhibitory effect of Brazilian red propolis on Candida biofilms developed on titanium surfaces”. BMC Complement. Med. Ther. 2020, 20, 104. [Google Scholar] [CrossRef] [Green Version]
- Stähli, A.; Schröter, H.; Bullitta, S.; Serralutzu, F.; Dore, A.; Nietzsche, S.; Milia, E.; Sculean, A.; Eick, S. In Vitro Activity of Propolis on Oral Microorganisms and Biofilms. Antibiotics 2021, 10, 1045. [Google Scholar] [CrossRef]
- Kirchhoff, L.; Dittmer, S.; Weisner, A.-K.; Buer, J.; Rath, P.-M.; Steinmann, J. Antibiofilm activity of antifungal drugs, including the novel drug olorofim, against Lomentospora prolificans. J. Antimicrob. Chemother. 2020, 75, 2133–2140. [Google Scholar] [CrossRef]
- Veiga, F.F.; Gadelha, M.C.; Da Silva, M.R.T.; Costa, M.I.; Kischkel, B.; De Castro-Hoshino, L.V.; Sato, F.; Baesso, M.L.; Voidaleski, M.F.; Vasconcellos-Pontello, V.; et al. Propolis Extract for Onychomycosis Topical Treatment: From Bench to Clinic. Front. Microbiol. 2018, 9, 779. [Google Scholar] [CrossRef] [Green Version]
- Rufatto, L.C.; Luchtenberg, P.; Garcia, C.; Thomassigny, C.; Bouttier, S.; Henriques, J.A.P.; Roesch-Ely, M.; Dumas, F.; Moura, S. Brazilian red propolis: Chemical composition and antibacterial activity determined using bioguided fractionation. Microbiol. Res. 2018, 214, 74–82. [Google Scholar] [CrossRef]
- de Francisco, L.; Pinto, D.; Rosseto, H.; Toledo, L.; Santos, R.; Tobaldini-Valério, F.; Svidzinski, T.; Bruschi, M.; Sarmento, B.; Oliveira, M.B.P.; et al. Evaluation of radical scavenging activity, intestinal cell viability and antifungal activity of Brazilian propolis by-product. Food Res. Int. 2018, 105, 537–547. [Google Scholar] [CrossRef]
- Galletti, J.; Tobaldini-Valerio, F.K.; Silva, S.; Kioshima, S.; Trierveiler-Pereira, L.; Bruschi, M.; Negri, M.; Svidzinski, T.I.E. Antibiofilm activity of propolis extract on Fusarium species from onychomycosis. Futur. Microbiol. 2017, 12, 1311–1321. [Google Scholar] [CrossRef] [Green Version]
- ISO DOCUMENT 10993-5; Biological Evaluation of Medical Devices—Part 5, Tests for Cytotoxicity: In Vitro Methods. The British Standards Institution: London, UK, 2009. [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Suffness, M.; Pezzuto, J.M. Methods in Plant Biochemistry: Assays for Bioactivity. In Methods in Plant Biochemistry, 6th ed.; Hostettmann, K., Ed.; Academic Press: London, UK, 1991; pp. 33–71. Available online: https://books.google.com/books/about/Methods_in_Plant_Biochemistry.html?hl=&id=M567oAEACAAJ (accessed on 3 August 2022).





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, I.L.E.; Veiga, F.F.; de Castro-Hoshino, L.V.; Souza, M.; Malacrida, A.M.; Diniz, B.V.; dos Santos, R.S.; Bruschi, M.L.; Baesso, M.L.; Negri, M.; et al. Performance of Two Extracts Derived from Propolis on Mature Biofilm Produced by Candida albicans. Antibiotics 2023, 12, 72. https://doi.org/10.3390/antibiotics12010072
Barros ILE, Veiga FF, de Castro-Hoshino LV, Souza M, Malacrida AM, Diniz BV, dos Santos RS, Bruschi ML, Baesso ML, Negri M, et al. Performance of Two Extracts Derived from Propolis on Mature Biofilm Produced by Candida albicans. Antibiotics. 2023; 12(1):72. https://doi.org/10.3390/antibiotics12010072
Chicago/Turabian StyleBarros, Isabella Letícia Esteves, Flávia Franco Veiga, Lidiane Vizioli de Castro-Hoshino, Monique Souza, Amanda Milene Malacrida, Beatriz Vesco Diniz, Rafaela Said dos Santos, Marcos Luciano Bruschi, Mauro Luciano Baesso, Melyssa Negri, and et al. 2023. "Performance of Two Extracts Derived from Propolis on Mature Biofilm Produced by Candida albicans" Antibiotics 12, no. 1: 72. https://doi.org/10.3390/antibiotics12010072
APA StyleBarros, I. L. E., Veiga, F. F., de Castro-Hoshino, L. V., Souza, M., Malacrida, A. M., Diniz, B. V., dos Santos, R. S., Bruschi, M. L., Baesso, M. L., Negri, M., & Svidzinski, T. I. E. (2023). Performance of Two Extracts Derived from Propolis on Mature Biofilm Produced by Candida albicans. Antibiotics, 12(1), 72. https://doi.org/10.3390/antibiotics12010072