

Treatment of Mouse Infants with Amoxicillin, but Not the Human Milk-Derived Antimicrobial HAMLET, Impairs Lung Th17 Responses

 , and
, and 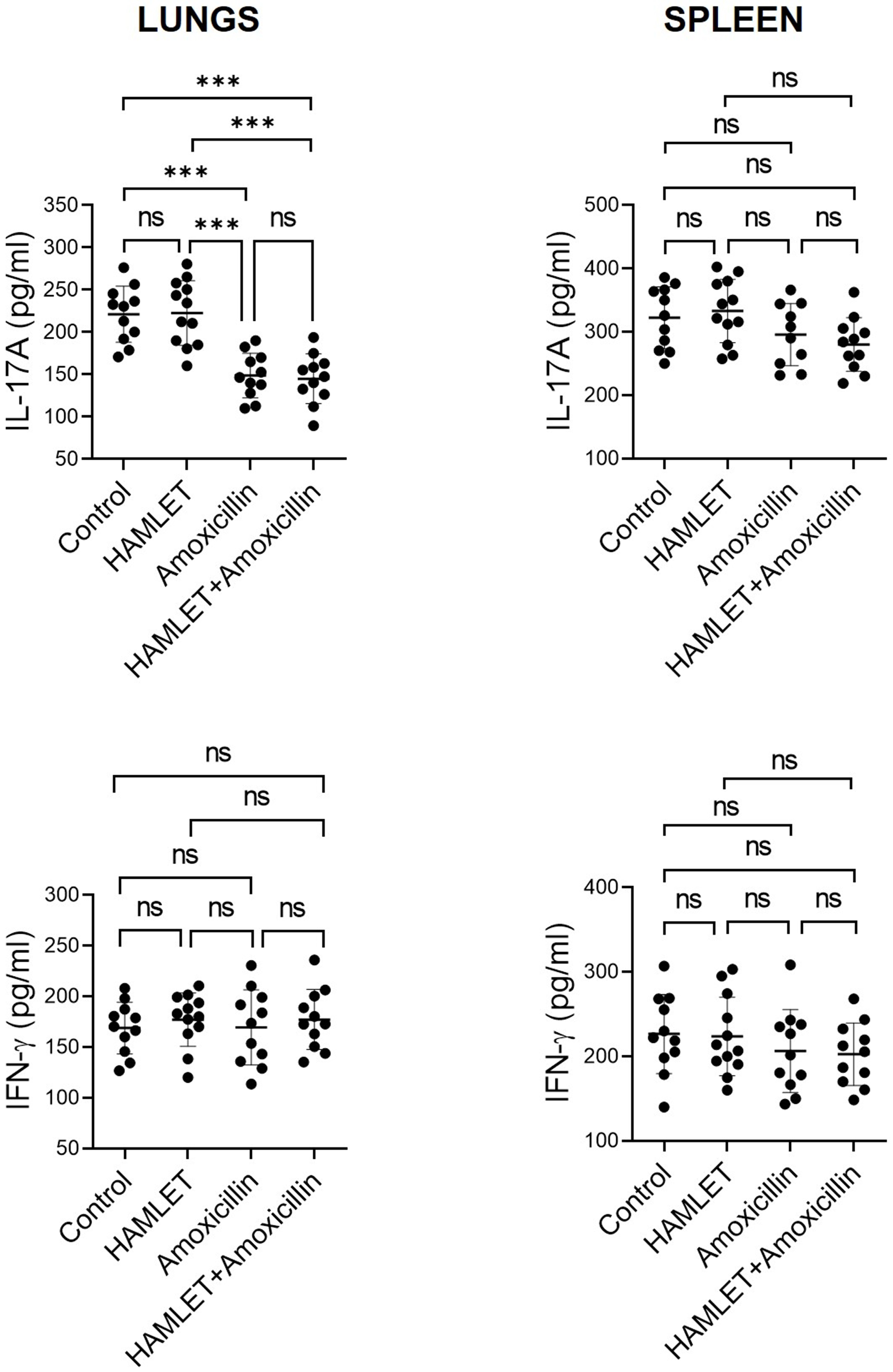{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Treatment of Infant Mice with Amoxicillin, but Not HAMLET, Suppresses Lung IL-17A Responses to S. pneumoniae
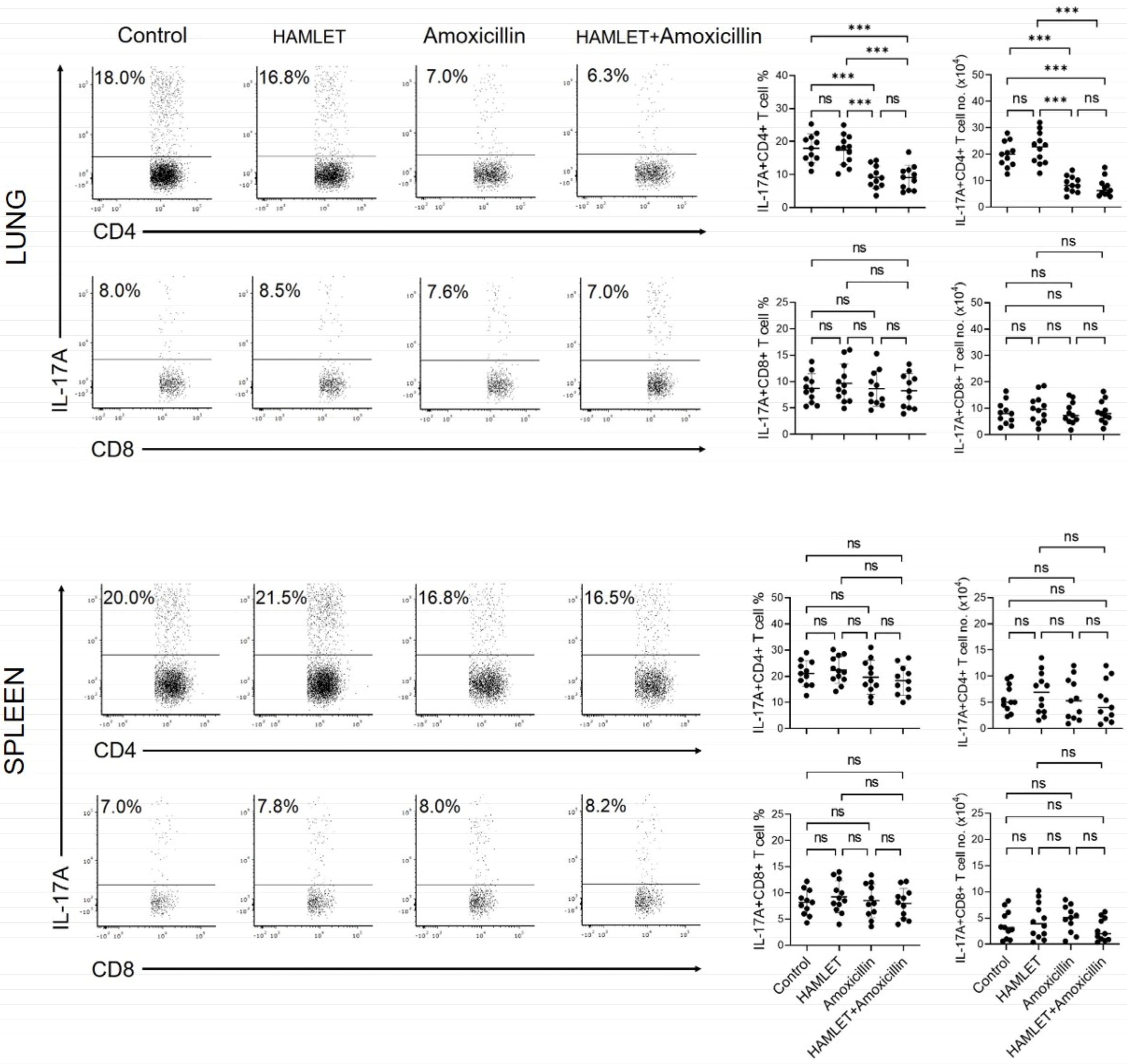
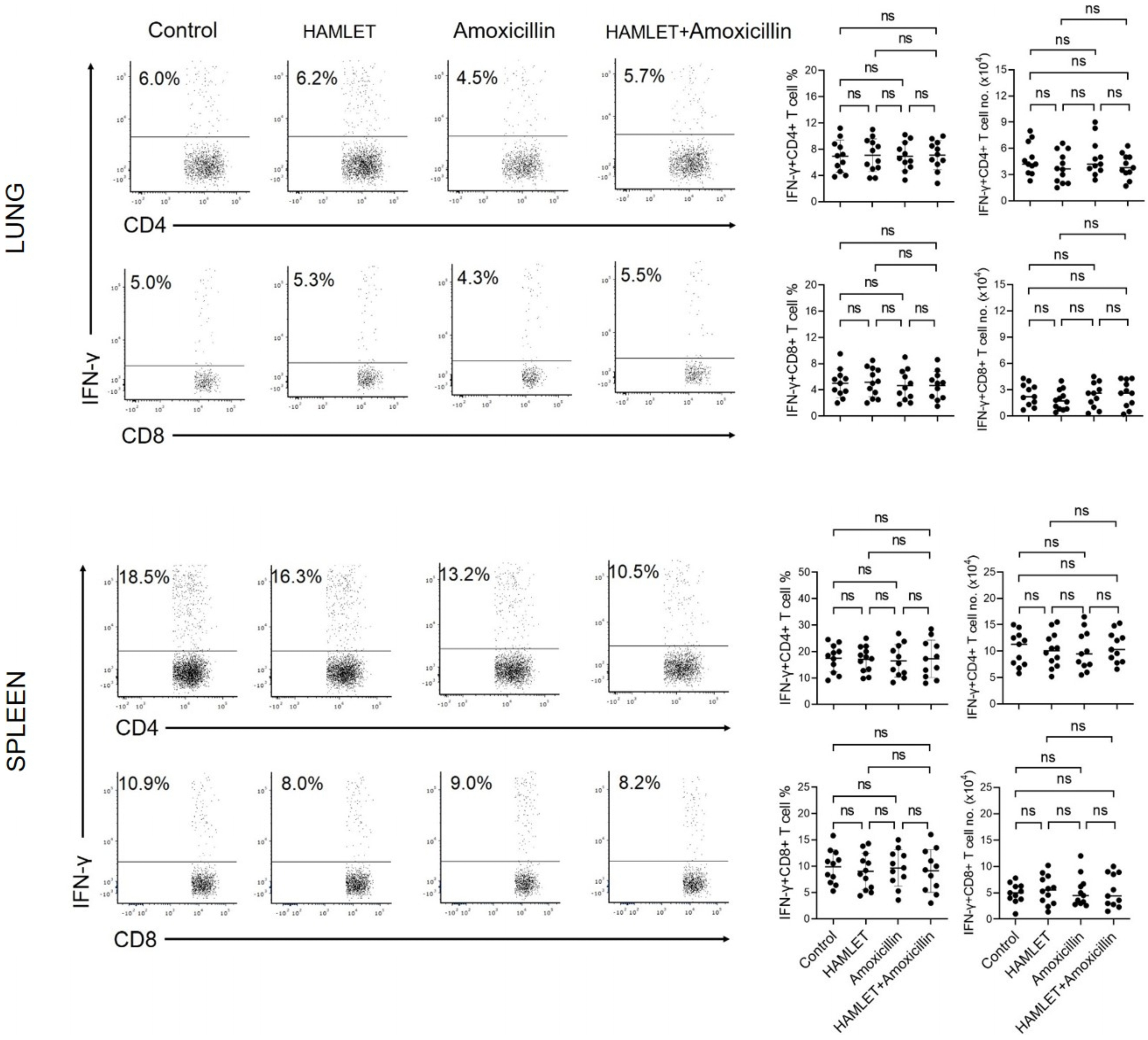
2.2. Exposure of Infants to Amoxicillin Diminishes Th17, but Not Th1, Responses
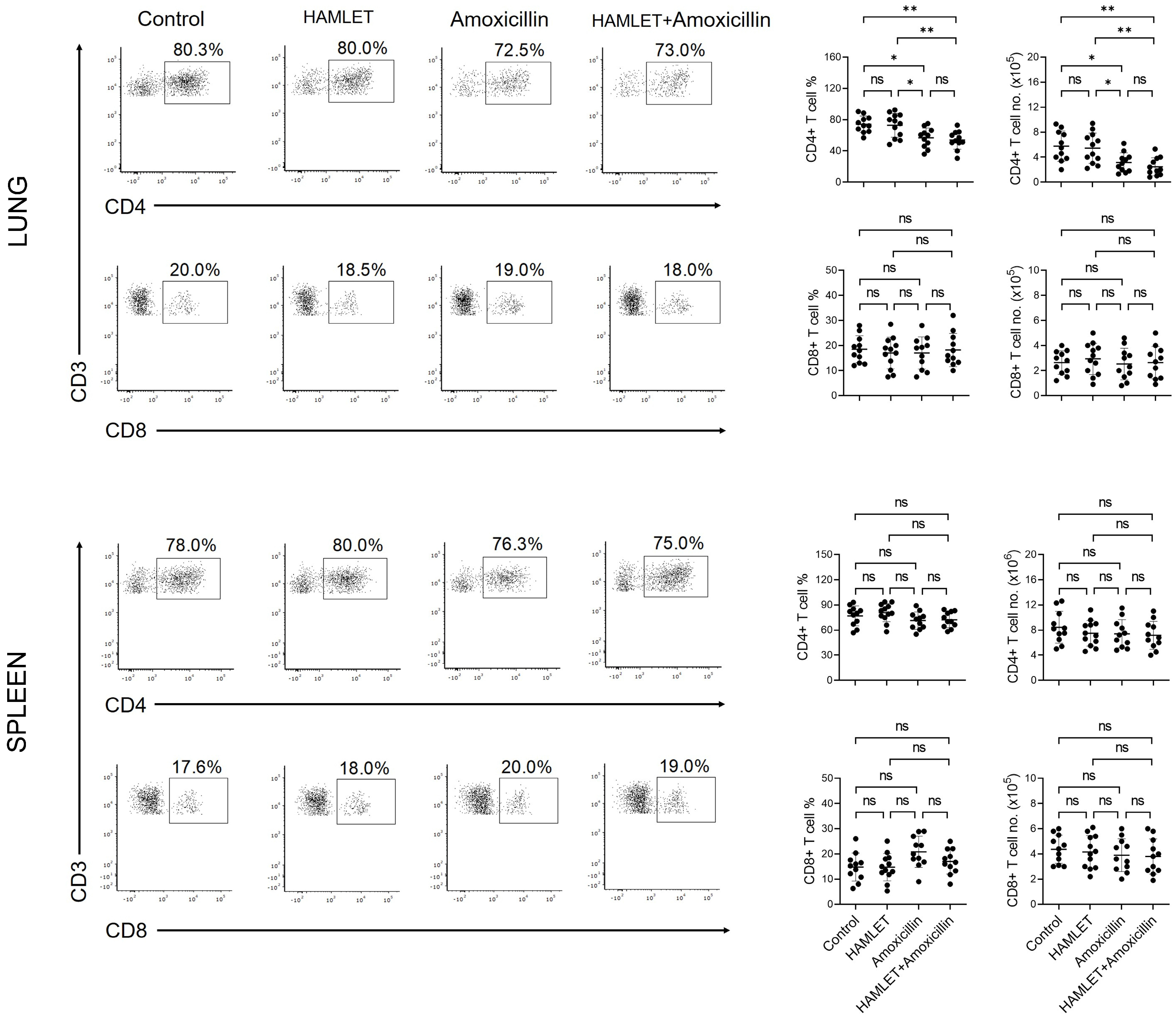
2.3. Amoxicillin Alone or with HAMLET Reduces the CD4+ T Cell Number
3. Discussion
4. Materials and Methods
4.1. Streptococcus pneumoniae
4.2. HAMLET Production
4.3. Mice
4.4. Cell Isolation and Antigenic Stimulation
4.5. Flow Cytometric Analysis
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhutta, Z.A.; Black, R.E. Global Maternal, Newborn, and Child Health—So Near and Yet So Far. N. Engl. J. Med. 2013, 369, 2226–2235. [Google Scholar] [CrossRef] [Green Version]
- Wagstaff, J.S.; Durrant, R.J.; Newman, M.G.; Eason, R.; Ward, R.M.; Sherwin, C.M.T.; Enioutina, E.Y. Antibiotic Treatment of Suspected and Confirmed Neonatal Sepsis within 28 Days of Birth: A Retrospective Analysis. Front. Pharmacol. 2019, 10, 1191. [Google Scholar] [CrossRef] [Green Version]
- Shane, A.L.; Sanchez, P.J.; Stoll, B.J. Neonatal sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Jackson, C.; Hsia, Y.; Basmaci, R.; Bielicki, J.; Heath, P.T.; Versporten, A.; Goossens, H.; Sharland, M. Global Divergence From World Health Organization Treatment Guidelines for Neonatal and Pediatric Sepsis. Pediatr. Infect. Dis. J. 2019, 38, 1104–1106. [Google Scholar] [CrossRef]
- Sutherland, R.; Croydon, E.A.; Rolinson, G.N. Amoxycillin: A new semi-synthetic penicillin. Br. Med. J. 1972, 3, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Abgueguen, P.; Azoulay-Dupuis, E.; Noel, V.; Moine, P.; Rieux, V.; Fantin, B.; Bedos, J.P. Amoxicillin is effective against penicillin-resistant Streptococcus pneumoniae strains in a mouse pneumonia model simulating human pharmacokinetics. Antimicrob. Agents Chemother. 2007, 51, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Petersen, F.C. The Dark Side of Antibiotics: Adverse Effects on the Infant Immune Defense Against Infection. Front. Pediatr. 2020, 8, 544460. [Google Scholar] [CrossRef]
- Zeissig, S.; Blumberg, R.S. Life at the beginning: Perturbation of the microbiota by antibiotics in early life and its role in health and disease. Nat. Immunol. 2014, 15, 307–310. [Google Scholar] [CrossRef]
- Neuman, H.; Forsythe, P.; Uzan, A.; Avni, O.; Koren, O. Antibiotics in early life: Dysbiosis and the damage done. FEMS Microbiol. Rev. 2018, 42, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Perez, G.; Hicks, A.L.; Tekieli, T.M.; Radens, C.M.; Williams, B.L.; Lamouse-Smith, E.S.N. Maternal Antibiotic Treatment Impacts Development of the Neonatal Intestinal Microbiome and Antiviral Immunity. J. Immunol. 2016, 196, 3768–3779. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Perez, G.; Lamouse-Smith, E.S.N. Gastrointestinal Microbiome Dysbiosis in Infant Mice Alters Peripheral CD8(+) T Cell Receptor Signaling. Front. Immunol. 2017, 8, 265. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Brar, N.K.; Petersen, F.C. Suppressive effect of therapeutic antibiotic regimen on antipneumococcal Th1/Th17 responses in neonatal mice. Pediatr. Res. 2022. [Google Scholar] [CrossRef]
- Jernberg, C.; Lofmark, S.; Edlund, C.; Jansson, J.K. Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology 2010, 156, 3216–3223. [Google Scholar] [CrossRef] [Green Version]
- Fouhse, J.M.; Yang, K.Y.; More-Bayona, J.; Gao, Y.H.; Goruk, S.; Plastow, G.; Field, C.J.; Barreda, D.R.; Willing, B.P. Neonatal Exposure to Amoxicillin Alters Long-Term Immune Response Despite Transient Effects on Gut-Microbiota in Piglets. Front. Immunol. 2019, 10, 2059. [Google Scholar] [CrossRef] [Green Version]
- Lynn, M.A.; Tumes, D.J.; Choo, J.M.; Sribnaia, A.; Blake, S.J.; Leong, L.E.X.; Young, G.P.; Marshall, H.S.; Wesselingh, S.L.; Rogers, G.B.; et al. Early-Life Antibiotic-Driven Dysbiosis Leads to Dysregulated Vaccine Immune Responses in Mice. Cell Host Microbe 2018, 23, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Hakansson, A.; Svensson, M.; Mossberg, A.K.; Sabharwal, H.; Linse, S.; Lazou, I.; Lonnerdal, B.; Svanborg, C. A folding variant of alpha-lactalbumin with bactericidal activity against Streptococcus pneumoniae. Mol. Microbiol. 2000, 35, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Clementi, E.A.; Marks, L.R.; Duffey, M.E.; Hakansson, A.P. A novel initiation mechanism of death in Streptococcus pneumoniae induced by the human milk protein-lipid complex HAMLET and activated during physiological death. J. Biol. Chem. 2012, 287, 27168–27182. [Google Scholar] [CrossRef] [Green Version]
- Meikle, V.; Mossberg, A.K.; Mitra, A.; Hakansson, A.P.; Niederweis, M. A Protein Complex from Human Milk Enhances the Activity of Antibiotics and Drugs against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2019, 63, e01846-18. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.R.; Clementi, E.A.; Hakansson, A.P. The human milk protein-lipid complex HAMLET sensitizes bacterial pathogens to traditional antimicrobial agents. PLoS ONE 2012, 7, e43514. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.R.; Clementi, E.A.; Hakansson, A.P. Sensitization of Staphylococcus aureus to methicillin and other antibiotics in vitro and in vivo in the presence of HAMLET. PLoS ONE 2013, 8, e63158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamiri, F.; Riesbeck, K.; Hakansson, A.P. HAMLET, a Protein Complex from Human Milk, Has Bactericidal Activity and Enhances the Activity of Antibiotics against Pathogenic Streptococci. Antimicrob. Agents Chemother. 2019, 63, e01193-19. [Google Scholar] [CrossRef] [PubMed]
- Vansarla, G.; Hakansson, A.P.; Bergenfelz, C. HAMLET a human milk protein-lipid complex induces a pro-inflammatory phenotype of myeloid cells. Eur. J. Immunol. 2021, 51, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Levine, O.S.; O’Brien, K.L.; Knoll, M.; Adegbola, R.A.; Black, S.; Cherian, T.; Dagan, R.; Goldblatt, D.; Grange, A.; Greenwood, B.; et al. Pneumococcal vaccination in developing countries. Lancet 2006, 367, 1880–1882. [Google Scholar] [CrossRef]
- Lundgren, A.; Bhuiyan, T.R.; Novak, D.; Kaim, J.; Reske, A.; Lu, Y.J.; Qadri, F.; Malley, R. Characterization of Th17 responses to Streptococcus pneumoniae in humans: Comparisons between adults and children in a developed and a developing country. Vaccine 2012, 30, 3897–3907. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Cohen, J.M.; Jose, R.J.; de Vogel, C.; Baxendale, H.; Brown, J.S. Protection against Streptococcus pneumoniae lung infection after nasopharyngeal colonization requires both humoral and cellular immune responses. Mucosal Immunol. 2015, 8, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Malley, R.; Trzcinski, K.; Srivastava, A.; Thompson, C.M.; Anderson, P.W.; Lipsitch, M. CD4+ T cells mediate antibody-independent acquired immunity to pneumococcal colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 4848–4853. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Yao, J.; Deng, L.; Yang, X.; Luo, W.; Zhou, W. Pretreatment with Antibiotics Impairs Th17-Mediated Antifungal Immunity in Newborn Rats. Inflammation 2020, 43, 2202–2208. [Google Scholar] [CrossRef]
- Zambrano-Zaragoza, J.F.; Romo-Martinez, E.J.; Duran-Avelar Mde, J.; Garcia-Magallanes, N.; Vibanco-Perez, N. Th17 cells in autoimmune and infectious diseases. Int. J. Inflam. 2014, 2014, 651503. [Google Scholar] [CrossRef]
- Mardikasari, S.A.; Sipos, B.; Csoka, I.; Katona, G. Nasal route for antibiotics delivery: Advances, challenges and future opportunities applying the quality by design concepts. J. Drug Deliv. Sci. Technol. 2022, 77, 103887. [Google Scholar] [CrossRef]
- Olliver, M.; Hiew, J.; Mellroth, P.; Henriques-Normark, B.; Bergman, P. Human monocytes promote Th1 and Th17 responses to Streptococcus pneumoniae. Infect. Immun. 2011, 79, 4210–4217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Clarke, T.B.; Weiser, J.N. Cellular effectors mediating Th17-dependent clearance of pneumococcal colonization in mice. J. Clin. Invest. 2009, 119, 1899–1909. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.K.; Bangert, M.; Gritzfeld, J.F.; Ferreira, D.M.; Jambo, K.C.; Wright, A.D.; Collins, A.M.; Gordon, S.B. Experimental human pneumococcal carriage augments IL-17A-dependent T-cell defence of the lung. PLoS Pathog 2013, 9, e1003274. [Google Scholar] [CrossRef]
- Lee, A.; Wong, S.Y.; Chai, L.Y.A.; Lee, S.C.; Lee, M.X.; Muthiah, M.D.; Tay, S.H.; Teo, C.B.; Tan, B.K.J.; Chan, Y.H.; et al. Efficacy of covid-19 vaccines in immunocompromised patients: Systematic review and meta-analysis. BMJ 2022, 376, e068632. [Google Scholar] [CrossRef]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; DeBoy, R.T.; Haft, D.H.; Dodson, R.J.; et al. Complete genome sequence of a virulent isolate of Streptococcus pneumoniae. Science 2001, 293, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Svensson, M.; Hakansson, A.; Mossberg, A.K.; Linse, S.; Svanborg, C. Conversion of alpha-lactalbumin to a protein inducing apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 4221–4226. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Khan, R.; Schenck, K.; Petersen, F.C. Intranasal immunization with the commensal Streptococcus mitis confers protective immunity against pneumococcal lung infection. Appl. Environ. Microbiol. 2019. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shekhar, S.; Brar, N.K.; Håkansson, A.P.; Petersen, F.C. Treatment of Mouse Infants with Amoxicillin, but Not the Human Milk-Derived Antimicrobial HAMLET, Impairs Lung Th17 Responses. Antibiotics 2023, 12, 423. https://doi.org/10.3390/antibiotics12020423
Shekhar S, Brar NK, Håkansson AP, Petersen FC. Treatment of Mouse Infants with Amoxicillin, but Not the Human Milk-Derived Antimicrobial HAMLET, Impairs Lung Th17 Responses. Antibiotics. 2023; 12(2):423. https://doi.org/10.3390/antibiotics12020423
Chicago/Turabian StyleShekhar, Sudhanshu, Navdeep Kaur Brar, Anders P. Håkansson, and Fernanda Cristina Petersen. 2023. "Treatment of Mouse Infants with Amoxicillin, but Not the Human Milk-Derived Antimicrobial HAMLET, Impairs Lung Th17 Responses" Antibiotics 12, no. 2: 423. https://doi.org/10.3390/antibiotics12020423
APA StyleShekhar, S., Brar, N. K., Håkansson, A. P., & Petersen, F. C. (2023). Treatment of Mouse Infants with Amoxicillin, but Not the Human Milk-Derived Antimicrobial HAMLET, Impairs Lung Th17 Responses. Antibiotics, 12(2), 423. https://doi.org/10.3390/antibiotics12020423