

Artificial Neural Network Prediction of Antiadhesion and Antibiofilm-Forming Effects of Antimicrobial Active Mushroom Extracts on Food-Borne Pathogens
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Activity
2.2. Antiadhesion and Antibiofilm-Forming Activity
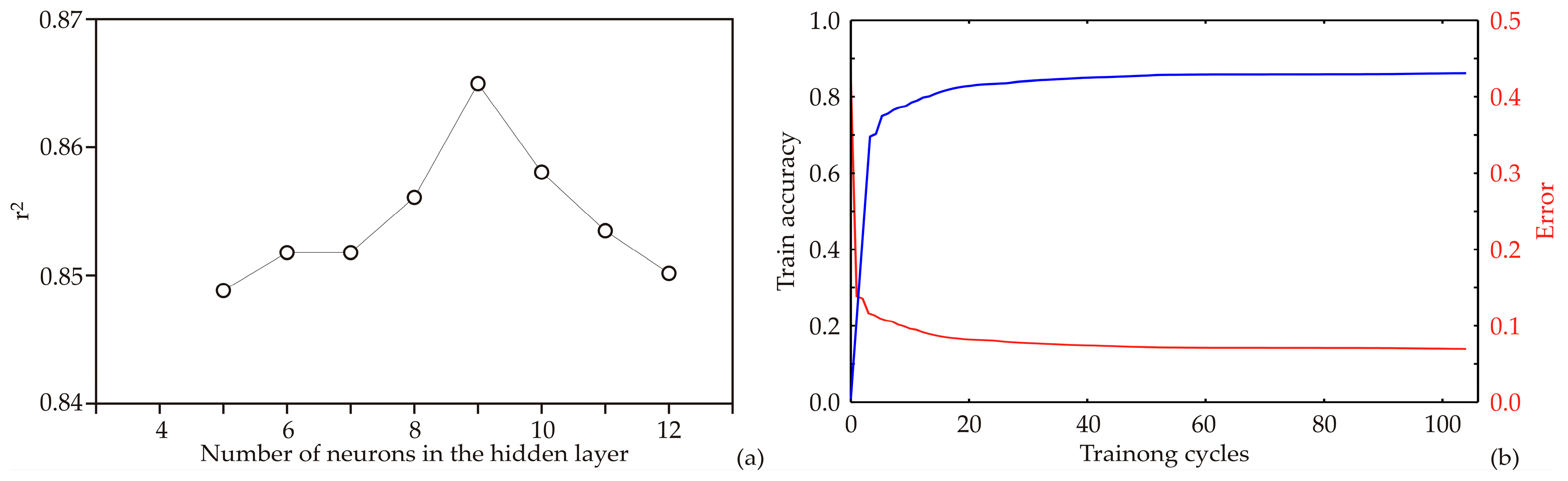
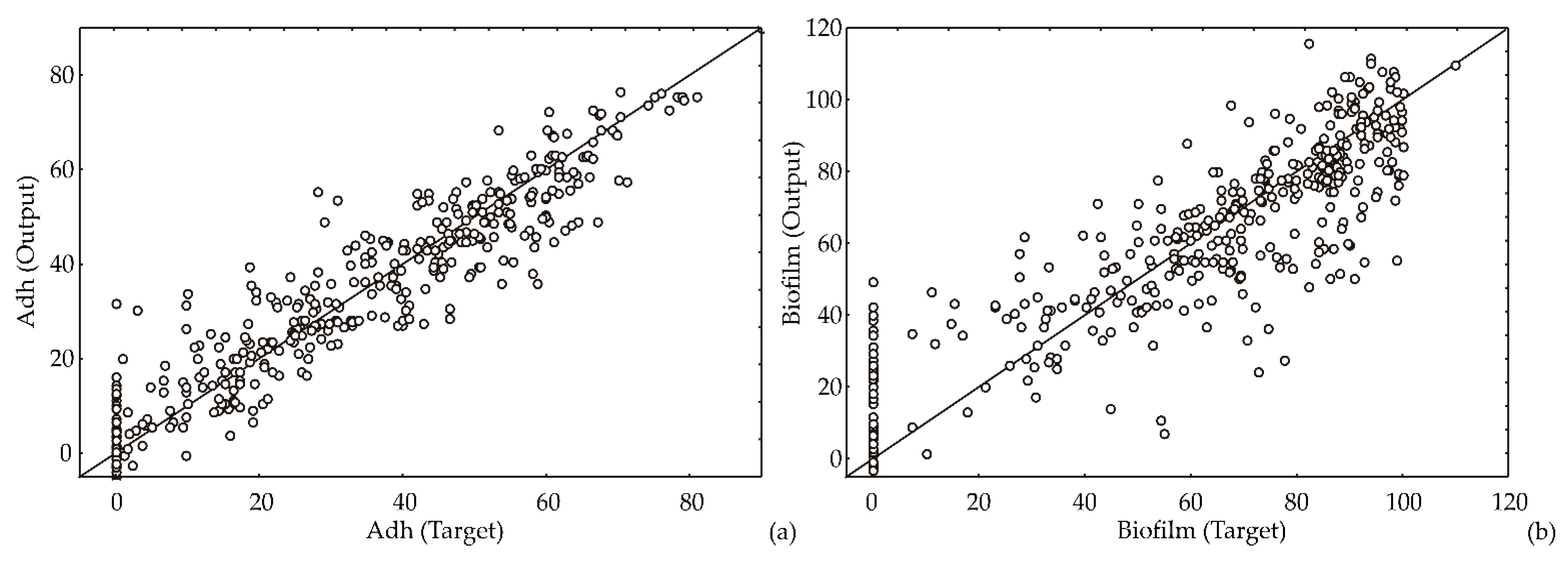
2.3. Artificial Neural Network Model
3. Discussion
3.1. Antimicrobial and Antibiofilm Activity
3.2. ANN Model
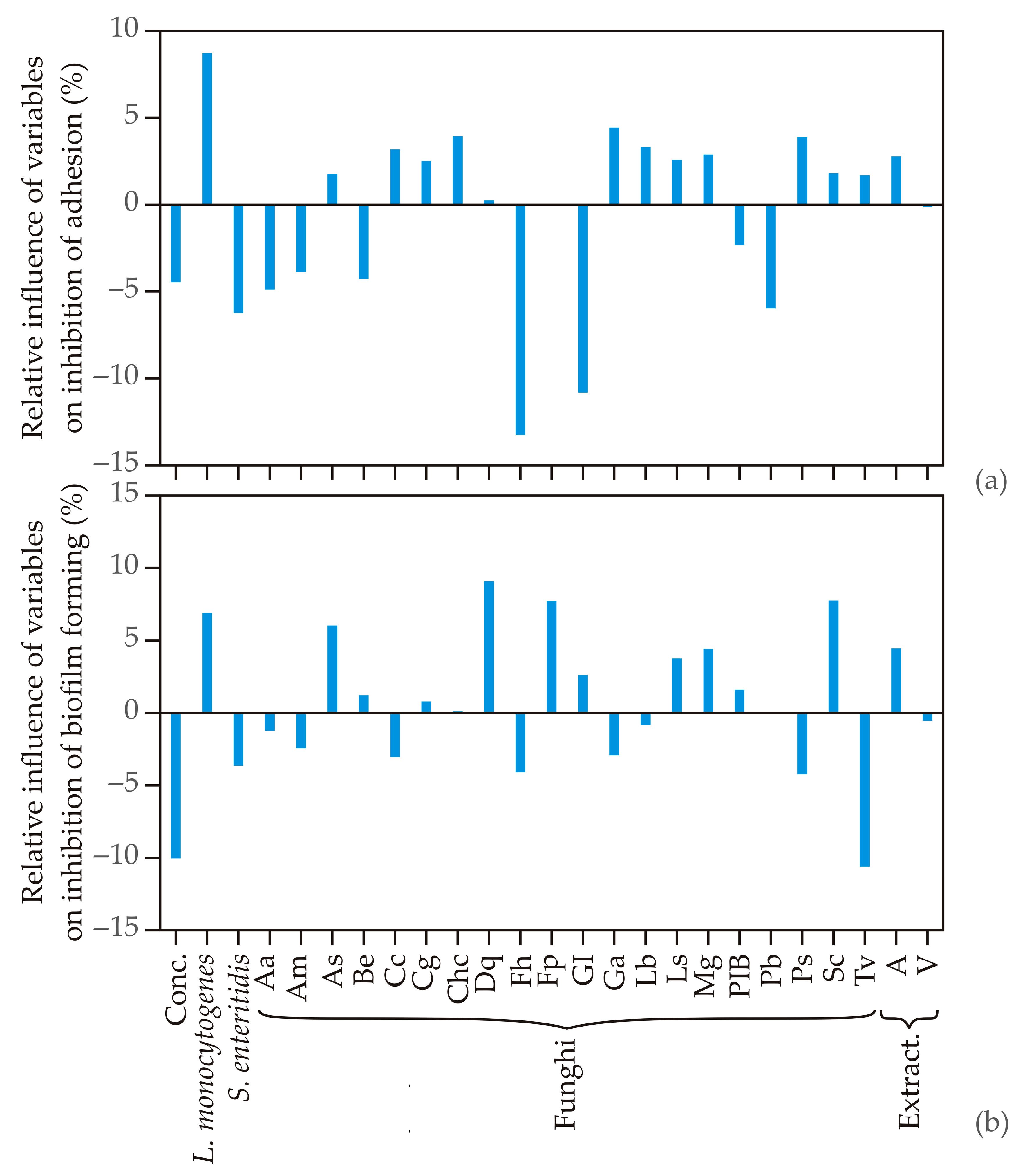
3.3. Global Sensitivity Analysis-Yoon’s Interpretation Method
4. Materials and Methods
4.1. Reagents
4.2. Mushroom Samples Collection and Identification
4.3. Preparation of Mushroom Extracts
4.4. Bacterial Strains and Culture Conditions
4.5. Determination of Mínimum Inhibitory Concentrations (MIC)
4.6. Antiadhesion and Antibiofilm-Forming Assay
4.7. Artificial Neural Network (ANN)
Global Sensitivity Analysis
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Drug Administration, Outbreak Investigation of Listeria monocytogenes: Dole Packaged Salad (December 2021). Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/outbreak-investigation-listeria-monocytogenes-dole-packaged-salad-december-2021 (accessed on 26 December 2022).
- Food Safety News. Biofilms with Listeria Pose Persistent Threat in Food Production. Available online: https://www.foodsafetynews.com/2017/06/biofilms-with-listeria-pose-persistent-threat-in-food-production/#more-141170 (accessed on 26 December 2022).
- Wagner, E.M.; Pracser, N.; Thalguter, S.; Fischel, K.; Rammer, N.; Pospíšilová, L.; Alispahic, M.; Wagner, M.; Rychli, K. Identification of biofilm hotspots in a meat processing environment: Detection of spoilage bacteria in multi-species biofilms. Int. J. Food Microbiol. 2020, 328, 108668. [Google Scholar] [CrossRef] [PubMed]
- Highmore, C.J.; Melaugh, G.; Morris, R.J.; Parker, J.; Direito, S.O.L.; Romero, M.; Soukarieh, F.; Robertson, S.N.; Bamford, N.C. Translational challenges and opportunities in biofilm science: A BRIEF for the future. NPJ Biofilms Microbiomes 2022, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial Biofilms and Their Implications in Pathogenesis and Food Safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef] [PubMed]
- Cámara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Economic significance of biofilms: A multidisciplinary and cross-sectoral challenge. NPJ Biofilms Microbiomes 2022, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Toushik, S.H.; Mizan, F.R.; Hossain, I.; Ha, S.D. Fighting with old foes: The pledge of microbe-derived biological agents to defeat mono- and mixed-bacterial biofilms concerning food industries. Trends Food Sci. Technol. 2020, 99, 413–425. [Google Scholar] [CrossRef]
- Kjelleberg, S.; McDougald, D.; Rasmussen, T.B.; Givskov, M. Quorum-sensing inhibition. In Chemical Communication among Bacteria; Winans, S.C., Bassler, B.L., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 393–416. [Google Scholar]
- Al-Dhabi, N.A.; Arasu, M.V.; Rejiniemon, T.S. In vitro antibacterial, antifungal, antibiofilm, antioxidant and anticancer properties of isosteviol isolated from endangered medicinal plant Pittosporum tetrasoermum. Evid. Based Complement. Alternat. Med. 2015, 2015, 164261. [Google Scholar]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural Anti-biofilm Agents: Strategies to Control Biofilm-Forming Pathogens. Front Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Golberg, K.; Markus, V.; Kagan, B.E.; Barzanizan, S.; Yaniv, K.; Teralı, K.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A. Anti-Virulence Activity of 3,3′-Diindolylmethane (DIM): A Bioactive Cruciferous Phytochemical with Accelerated Wound Healing Benefits. Pharmaceutics 2022, 14, 967. [Google Scholar] [CrossRef]
- Balaban, M.; Koc, C.; Sar, T.; Akbas, M.Y. Antibiofilm effects of pomegranate peel extracts against B. cereus, B. subtilis, and E. faecalis. Int. J. Food Sci. Technol. 2021, 56, 4915–4924. [Google Scholar] [CrossRef]
- Elisashvili, V.; Asatiani, M.D.; Khardziani, T.; Rai, M. Natural antimicrobials from Basidiomycota Mushrooms. In Promising Antimicrobials from Natural Products; Rai, M., Kosalec, I., Eds.; Springer: Cham, Switzerland, 2022; pp. 323–353. [Google Scholar]
- Alves, M.J.; Ferreira, I.C.; Lourenço, I.; Costa, E.; Martins, A.; Pintado, M. Wild mushroom extracts as inhibitors of bacterial biofilm formation. Pathogens 2014, 3, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Klančnik, A.; Megušar, P.; Sterniša, M.; Jeršek, B.; Bucar, F.; SmoleMožina, S.; Kos, J.; Sabotič, J. Aqueous extracts of wild mushrooms show antimicrobial and antiadhesion activities against bacteria and fungi. Phytother. Res. 2017, 31, 1971–1976. [Google Scholar] [CrossRef]
- Signoretto, C.; Marchi, A.; Bertoncelli, A.; Burlacchini, G.; Papetti, A.; Pruzzo, C.; Zaura, E.; Lingström, P.; Ofek, I.; Pratten, J.; et al. The anti-adhesive mode of action of purified mushroom (Lentinus edodes) extract with anticaries and antigingivitis properties in two oral bacterial pathogens. Complement. Altern. Med. 2014, 14, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papetti, A.; Signoretto, C.; Spratt, D.A.; Pratten, J.; Lingström, P.; Zaura, E.; Ofek, I.; Wilson, M.; Pruzzo, C.; Gazzani, G. Components in Lentinus edodes mushroom with anti-biofilm activity directed against bacteria involved in caries and gingivitis. Food Funct. 2018, 9, 3489–3499. [Google Scholar] [CrossRef]
- Vunduk, J.; Wan-Mohtar, W.A.A.Q.I.; Mohamad, S.A.; Halim, N.H.A.; Dzomir, A.Z.M.; Žižak, Ž.; Klaus, A. Polysaccharides of Pleurotus flabellatus strain Mynuk produced by submerged fermentation as a promising novel tool against adhesion and biofilm formation of foodborne pathogens. LWT 2019, 112, 108221. [Google Scholar] [CrossRef]
- Song, X.; Gaascht, F.; Schmidt-Dannert, C.; Salomon, C.E. Discovery of antifungal and biofilm preventive compounds from mycelial cultures of a unique North American Hericium sp. fungus. Molecules 2020, 25, 963. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Oh, S.; Kim, S.H. Released exopolysaccharide (r-ESP) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157:H7. Biochem. Biophys. Res. Commun. 2009, 379, 324–329. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Library of Medicine. Available online: https://clinicaltrials.gov/ (accessed on 29 November 2022).
- U.S. National Library of Medicine. Antibacterial and Anti-biofilm Activity of Frankincense Extract Against Porphyromonas Gingivalis. Available online: https://clinicaltrials.gov/ct2/show/NCT04705714?cond=antibiofilm&draw=2&rank=3 (accessed on 29 November 2022).
- Shaban, T.F.; Alkawareek, M.Y. Prediction of qualitative activity of antibiotics using supervised machine learning techniques. Comput. Biol. Med. 2021, 140, 105065. [Google Scholar] [CrossRef]
- Wawrzyniak, J. Application of Artificial Neural Networks to Assess the Mycological State of Bulk Stored Rapeseeds. Agriculture 2020, 10, 567. [Google Scholar] [CrossRef]
- Tomić, A.; Šovljanski, O.; Nikolić, V.; Pezo, L.; Aćimović, M.; Cvetković, M.; Stanojev, J.; Kuzmanović, N.; Markov, S. Screening of Antifungal Activity of Essential Oils in Controlling Biocontamination of Historical Papers in Archives. Antibiotics 2023, 12, 103. [Google Scholar] [CrossRef]
- Šovljanski, O.; Saveljić, A.; Aćimović, M.; Šeregelj, V.; Pezo, L.; Tomić, A.; Ćetković, G.; Tešević, V. Biological Profiling of Essential Oils and Hydrolates of Ocimum basilicum var. Genovese and var. Minimum Originated from Serbia. Processes 2022, 10, 1893. [Google Scholar] [CrossRef]
- Aćimović, M.; Šovljanski, O.; Pezo, L.; Travičić, V.; Tomić, A.; Zheljazkov, V.D.; Ćetković, G.; Švarc-Gajić, J.; Brezo-Borjan, T.; Sofrenić, I. Variability in Biological Activities of Satureja montana Subsp. montana and Subsp. variegata Based on Different Extraction Methods. Antibiotics 2022, 11, 1235. [Google Scholar] [CrossRef]
- Marks, B.P. Status of microbial modeling in food process models. Compr. Rev. Food Sci. Food Saf. 2008, 7, 137–143. [Google Scholar] [CrossRef]
- Rajković, D.; Marjanović Jeromela, D.; Pezo, L.; Lončar, B.; Zanetti, F.; Monti, A.; Kondić Špika, A. Yield and Quality Prediction of Winter Rapeseed—Artificial Neural Network and Random Forest Models. Agronomy 2022, 12, 58. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 25 January 2023).
- Center for Disease Control. Antibiotic Resistance: A Global Threat. Available online: https://www.cdc.gov/drugresistance/solutions-initiative/stories/ar-global-threat.html (accessed on 25 January 2023).
- Alves, M.J.; Ferreira, I.C.F.R.; Martins, A.; Pintado, M. Antimicrobial activity of wild mushroom extracts against clinical isolates resistant to different antibiotics. J. Appl. Microbiol. 2012, 113, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Suay, I.; Arenal, A.; Asensio, F.J.; Basilio, A.; Cabello, M.A.; Diez, M.T.; Garcia, J.B.; González del Val, A.; Gorrochategui, J.; Hernández, P.; et al. Screening of basidiomycetes for antimicrobial activities. Antonie Van Leeuwenhoek 2000, 78, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Venturini, M.E.; Rivera, C.S.; Gonzalez, C.; Blanco, D. Antimicrobial activity of extracts of edible wild and cultivated mushrooms against foodborne bacterial strains. J. Food Prot. 2008, 71, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Huguet, C.; Bourjot, M.; Bellanger, J.M.; Prévost, G.; Urbain, A. Screening for antibacterial activity of French mushrooms against pathogenic and multidrug resistant bacteria. Appl. Sci. 2022, 12, 5229. [Google Scholar] [CrossRef]
- Barros, L.; Calhelha, R.; Vaz, J.; Ferreira, I.C.F.R.; Baptista, P.; Estevinho, L.M. Antimicrobial activity and bioactive compounds of Portuguese wild edible mushrooms methanolic extracts. Eur. Food Res. Technol. 2007, 225, 151–156. [Google Scholar] [CrossRef]
- Klaus, A.; Kozarski, M.; Niksic, M.; Jakovljevic, D.; Todorovic, N.; Van Griensven, L.J.L.D. Antioxidative activities and chemical characterization of polysaccharides extracted from the basidiomycete Schizophyllum commune. LWT-Food Sci. Technol. 2011, 44, 2005–2011. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; van Griensven, L.; Jakovljevic, D.; Todorovic, N.; Wan-Mohtar, W.A.A.Q.I. Mushroom β-glucan and polyphenol formulations as natural immunity boosters and balancers: Nature of the application. Food Sci. Hum. Wellness 2023, 12, 378–396. [Google Scholar] [CrossRef]
- Bernal, P.; Liamas, M.A. Promising biotechnological applications of antibiofilm exopolysaccharides. Microb. Biotechnol. 2012, 5, 670–673. [Google Scholar] [CrossRef] [Green Version]
- Moussa, A.Y.; Fayez, S.; Xiao, H.; Xu, B. New insights into antimicrobial and antibiofilm effects of edible mushrooms. Food Res. Int. 2022, 162, 111982. [Google Scholar] [CrossRef]
- Shomali, N.; Onar, O.; Cihan, A.C.; Akata, I.; Yildirim, O. Antioxidant, anticancer, antimicrobial, and antibiofilm properties of the culinary-medicinal fairy ring mushroom, Marasmius oreades (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Pu, Y.; Sun, L.; Wang, Y.; Liu, Y.; Wang, R.; Liao, J.; Xu, D.; Liu, Y.; Yr, R.; et al. Antimicrobial peptide AMPNT-6 from bacillus subtilis inhibits biofilm formation by Shewanella putrefaciens and disrupts its preformed biofilms on both abiotic and shrimp shell surfaces. Food Res. Int. 2017, 102, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.R.; Zhong, K.; Wu, Y.P.; Elena, G.; Gao, H. Antibiofilm activity of shikimic acid against Staphylococcus aureus. Food Control 2019, 95, 327–333. [Google Scholar] [CrossRef]
- Ye, J.; Shao, C.; Zhang, X.; Guo, X.; Gao, P.; Cen, Y.; Ma, S.; Liu, Y. Effects of DNase I coating of titanium on bacterial adhesion and biofilm formation. Mater. Sci. Eng. C 2017, 78, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Younus, M.; Wilkins, M.J.; Arshad, M.M.; Rahbar, M.H.; Saeed, A.M. Demographic risk factors and incidence of Salmonella enteritidis infection in Michigan. Foodborne Pathog. Dis. 2006, 3, 266–273. [Google Scholar] [CrossRef]
- Shamloo, E.; Hosseini, H.; Abdi Moghadam, Z.; Halberg, L.M.; Haslberger, A.; Alebouyeh, M. Importance of Listeria monocytogenes in food safety: A review of its prevalence, detection, and antibiotic resistance. Iran. J. Vet. Res. 2019, 20, 241–254. [Google Scholar]
- Al-Bakri, A.G.; Othman, G.; Afifi, F.U. Determination of the antibiofilm, antiadhesive, and anti-MRSA activities of seven Salvia species. Pharmacogn. Mag. 2010, 6, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Kavita, K.; Singh, V.K.; Mishra, A.; Jha, B. Characterization and anti-biofilm activity of extracellular polymeric substances from Oceanobacillus iheyensis. Carbohydr. Polym. 2014, 101, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Rodrigues, F.; Castro, F.; Aires, A.; Marques, G.; Saavedra, M.J. Antimicrobial, antibiofilm, and antioxidant properties of Boletus edulis and Neoboletus luridiformis against multidrug-resistant ESKAPE pathogens. Front. Nutr. 2022, 8, 1093. [Google Scholar] [CrossRef]
- Soković, M.; Ćirić, A.; Glamočlija, J.; Nikolić, M.; van Griensven, L.J.L.D. Agaricus blazei hot water extract shows anti quorum sensing activity in the nosocomial human pathogen Pseudomonas aeruginosa. Molecules 2014, 19, 4189–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, M.P.; Gulotta, G.; do Amaral, M.W.; Lünsdorf, H.; Sasse, F.; Abraham, W.R.A. Coprinuslactone protects the edible mushroom Coprinus comatus against biofilm infections by blocking both quorum-sensing and MurA. Environ. Microbiol. 2016, 18, 4254–4264. [Google Scholar] [CrossRef] [PubMed]
- Šeregelj, V.; Šovljanski, O.; TumbasŠaponjac, V.; Vulić, J.; Ćetković, G.; Markov, S.; Čanadanović-Brunet, J. Horned Melon (Cucumis metuliferus E. Meyer Ex. Naudin)—Current Knowledge on Its Phytochemicals, Biological Benefits, and Potential Applications. Processes 2022, 10, 94. [Google Scholar] [CrossRef]
- Aćimović, M.; Šovljanski, O.; Šeregelj, V.; Pezo, L.; Zheljazkov, V.D.; Ljujić, J.; Tomić, A.; Ćetković, G.; Čanadanović-Brunet, J.; Miljković, A.; et al. Chemical Composition, Antioxidant, and Antimicrobial Activity of Dracocephalum moldavica L. Essential Oil and Hydrolate. Plants 2022, 11, 941. [Google Scholar] [CrossRef]
- Pezo, L.; Curcic, B.; Filipovic, V.; Nicetin, M.; Koprivica, G.; Misljenovic, N.; Levic, L. Artificial neural network model of pork meat cubes osmotic dehydration. Hem. Ind. 2013, 67, 465–475. [Google Scholar] [CrossRef]
- Basheer, I.A.; Hajmeer, M. Artificial neural networks: Fundamentals, computing, design, and application. J. Microbiol. Methods 2000, 43, 3–31. [Google Scholar] [CrossRef]
- Montaño, J.J.; Palmer, A. Numeric sensitivity analysis applied to feedforward neural networks. Neural Comput. Appl. 2003, 12, 119–125. [Google Scholar] [CrossRef]
- Chattopadhyay, P.B.; Rangarajan, R. Application of ANN in sketching spatial nonlinearity of unconfined aquifer in agricultural basin. Agric. Water Manag. 2014, 113, 81–91. [Google Scholar] [CrossRef]
- Arsenovic, M.; Pezo, L.; Stankovic, S.; Radojevic, Z. Factor space differentiation of brick clays according to mineral content: Prediction of final brick product quality. Appl. Clay Sci. 2015, 115, 108–114. [Google Scholar] [CrossRef]
- Montgomery, D.C. Design and Analysis of Experiments; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 1984. [Google Scholar]
- Madamba, P.S. The Response Surface Methodology: An Application to Optimize Dehydration Operations of Selected Agricultural Crops. LWT 2002, 35, 584–592. [Google Scholar] [CrossRef]
- Klaus, A.; Kozarski, M.; Vunduk, J.; Todorovic, N.; Jakovljevic, D.; Zizak, Z.; Pavlovic, V.; Levic, S.; Niksic, M.; Van Griensven, L.J.L.D. Biological potential of extracts of the wild edible Basidiomycete mushroom Grifola frondosa. Food Res. Int. 2015, 67, 272–283. [Google Scholar] [CrossRef]
- Momenzadeh, L.; Zomorodian, A.; Mowla, D. Experimental and theoretical investigation of shelled corn drying in a microwave assisted fluidized bed dryer using Artificial Neural Network. Food Bioprod. Process. 2011, 89, 15–21. [Google Scholar] [CrossRef]
- Trelea, I.C.; Raoult-Wack, A.L.; Trystram, G. Note: Application of neural network modelling for the control of dewatering and impregnation soaking process (osmotic dehydration). Food Sci. Technol. Int. 1997, 3, 459–465. [Google Scholar] [CrossRef]
- Yoon, Y.; Swales, G.; Margavio, T.M. A comparison of discriminant analysis versus artificial neural networks. J. Oper. Res. Soc. 2017, 44, 51–60. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| L. monocytogenes | S. enteritidis | A Sample | L. monocytogenes | S. enteritidis | |||||
|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | ||
| MgV | 20 | / | 20 | / | MgA | 10 | 20 | 5 | / |
| AsV | / | / | 20 | / | AsA | 10 | / | 10 | / |
| CcV | 10 | / | 10 | / | CcA | 2.5 | / | 10 | / |
| GaV | 20 | / | 20 | / | GaA | 10 | / | 5 | / |
| PbV | / | / | / | / | PbA | 20 | / | 10 | / |
| LsV | 20 | / | / | / | LsA | 20 | / | 20 | / |
| ScV | / | / | / | / | ScA | / | / | / | / |
| FpV | / | / | 20 | / | FpA | 10 | / | 10 | / |
| BeV | 20 | / | 20 | / | BeA | 20 | / | 20 | / |
| CgV | / | / | / | / | CgA | / | / | 20 | / |
| LbV | 10 | / | 20 | / | LbA | 10 | / | 10 | / |
| PsV | 20 | / | / | / | PsA | 10 | / | 5 | / |
| ChcV | / | / | 10 | / | ChcA | 20 | / | 5 | / |
| AaV | 20 | / | 20 | / | AaA | 20 | / | 10 | / |
| TvV | / | / | / | / | TvA | 20 | / | 2.5 | / |
| DqV | 20 | / | 5 | / | DqA | 10 | / | 5 | / |
| AmV | 20 | / | 20 | / | AmA | 10 | / | 5 | / |
| PibV | 10 | / | / | / | PibA | 5 | / | 2.5 | / |
| GlV | 10 | / | 20 | / | GlA | 20 | 20 | 5 | / |
| FhV | 20 | / | 20 | / | FhA | 10 | / | 10 | / |
| Gentamicin | L. monocytogenes MIC < 0.0024 and MBC < 0.0024 S. enteritidis MIC < 0.0024 and MBC < 0.0024 | ||||||||
| Network Name | Performance | Error | Training Algorithm | Error Function | Hidden Activation | Output Activation | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Train. | Test. | Valid. | Train. | Test. | Valid. | |||||
| MLP 25-9-2 | 0.865 | 0.761 | 0.747 | 112.980 | 178.491 | 220.136 | BFGS 54 | SOS | Logistic | Identity |
| Output Variable | χ2 | RMSE | MBE | MPE | Skewness | Residual Analysis | |||
|---|---|---|---|---|---|---|---|---|---|
| Kurtoisis | Average | SD | Variance | ||||||
| Antiadhesion | 59.777 | 7.722 | −0.242 | 29.740 | −0.454 | 1.373 | −0.242 | 7.728 | 59.718 |
| Antibiofilm forming | 235.989 | 15.343 | 0.480 | 17.529 | 0.096 | 1.054 | 0.480 | 15.354 | 235.758 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vunduk, J.; Klaus, A.; Lazić, V.; Kozarski, M.; Radić, D.; Šovljanski, O.; Pezo, L. Artificial Neural Network Prediction of Antiadhesion and Antibiofilm-Forming Effects of Antimicrobial Active Mushroom Extracts on Food-Borne Pathogens. Antibiotics 2023, 12, 627. https://doi.org/10.3390/antibiotics12030627
Vunduk J, Klaus A, Lazić V, Kozarski M, Radić D, Šovljanski O, Pezo L. Artificial Neural Network Prediction of Antiadhesion and Antibiofilm-Forming Effects of Antimicrobial Active Mushroom Extracts on Food-Borne Pathogens. Antibiotics. 2023; 12(3):627. https://doi.org/10.3390/antibiotics12030627
Chicago/Turabian StyleVunduk, Jovana, Anita Klaus, Vesna Lazić, Maja Kozarski, Danka Radić, Olja Šovljanski, and Lato Pezo. 2023. "Artificial Neural Network Prediction of Antiadhesion and Antibiofilm-Forming Effects of Antimicrobial Active Mushroom Extracts on Food-Borne Pathogens" Antibiotics 12, no. 3: 627. https://doi.org/10.3390/antibiotics12030627
APA StyleVunduk, J., Klaus, A., Lazić, V., Kozarski, M., Radić, D., Šovljanski, O., & Pezo, L. (2023). Artificial Neural Network Prediction of Antiadhesion and Antibiofilm-Forming Effects of Antimicrobial Active Mushroom Extracts on Food-Borne Pathogens. Antibiotics, 12(3), 627. https://doi.org/10.3390/antibiotics12030627