
Pseudouridine Synthase RsuA Confers a Survival Advantage to Bacteria under Streptomycin Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
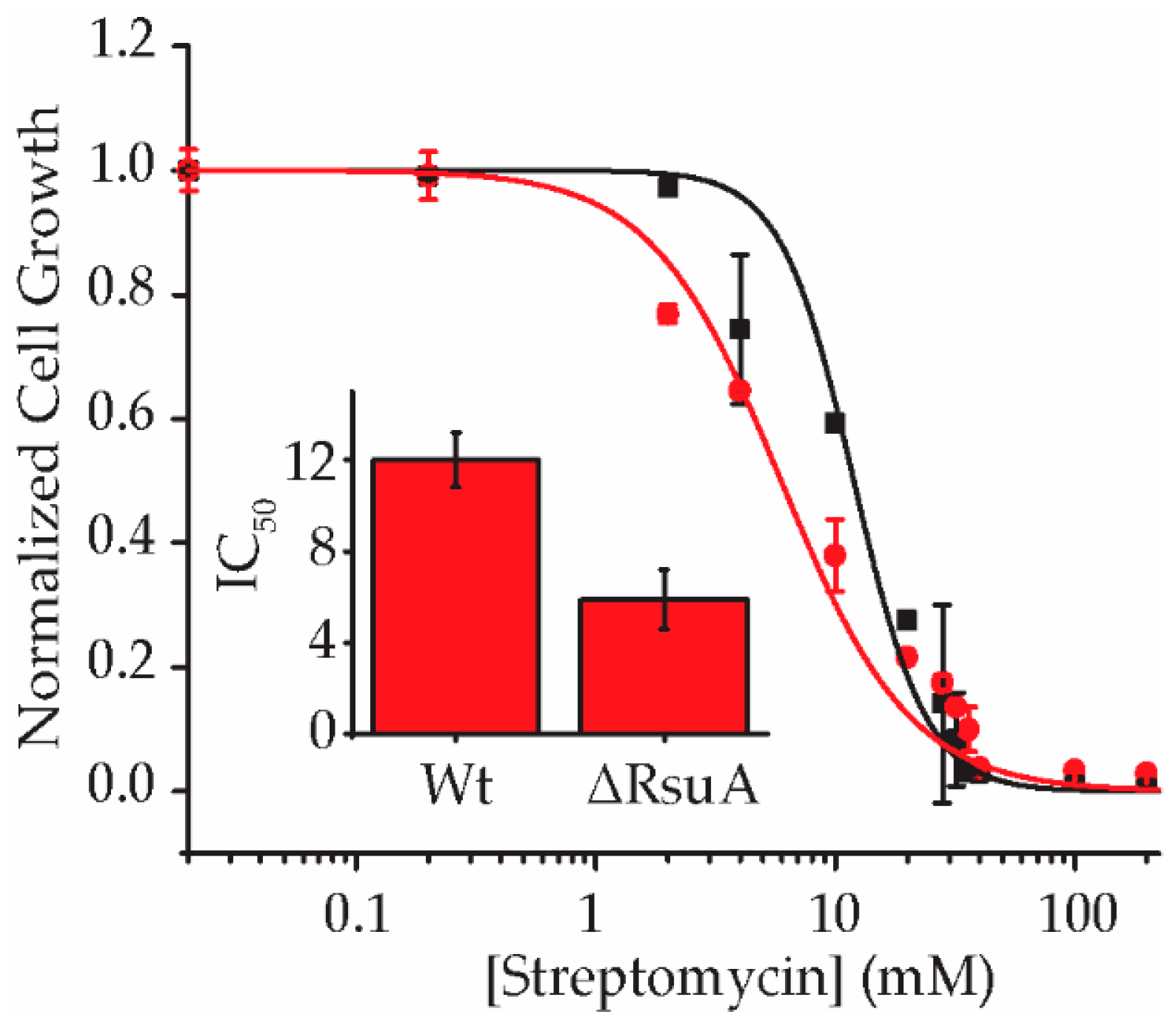
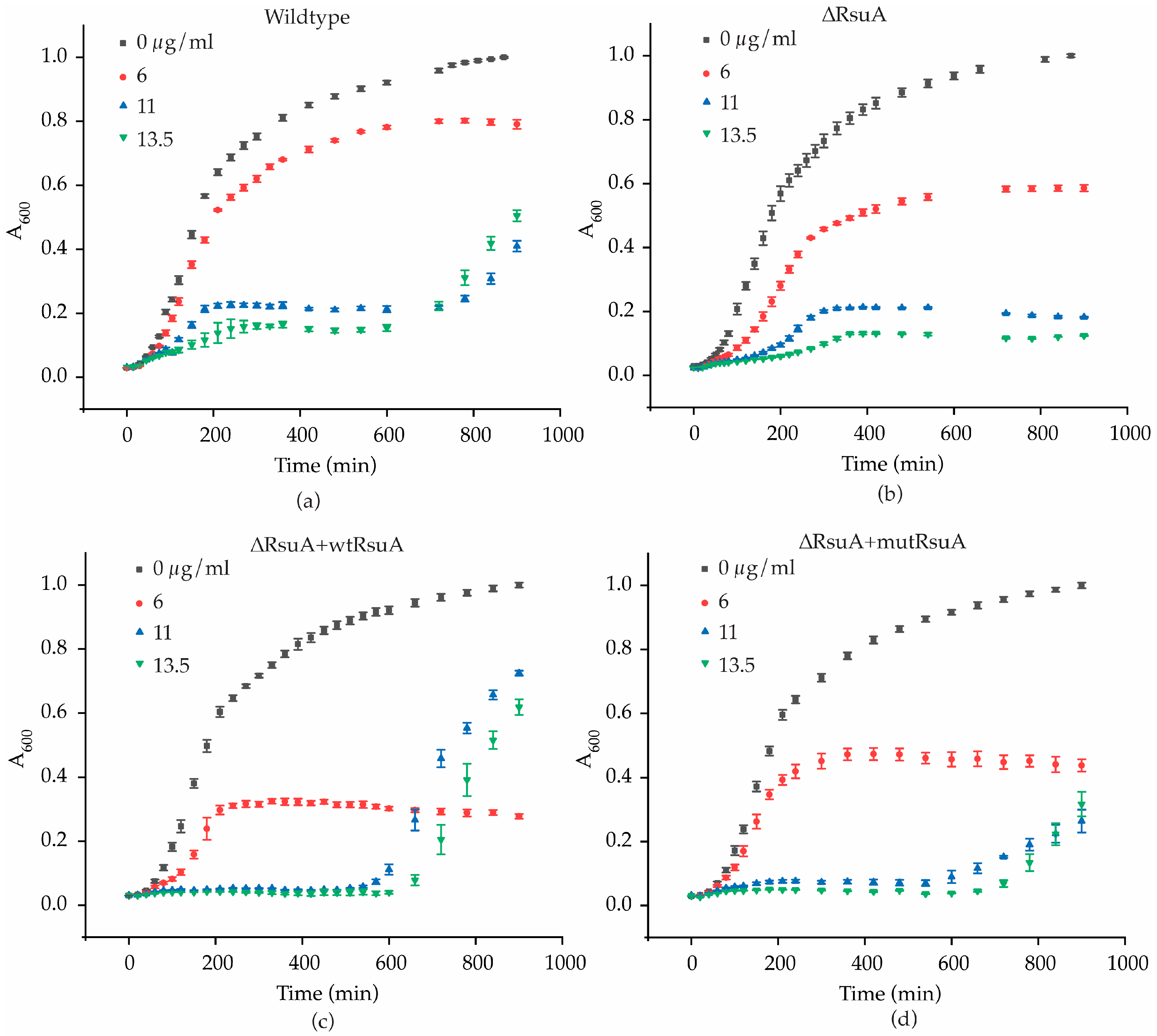
2.1. RsuA Provides a Survival Advantage to Bacteria under Streptomycin Stress
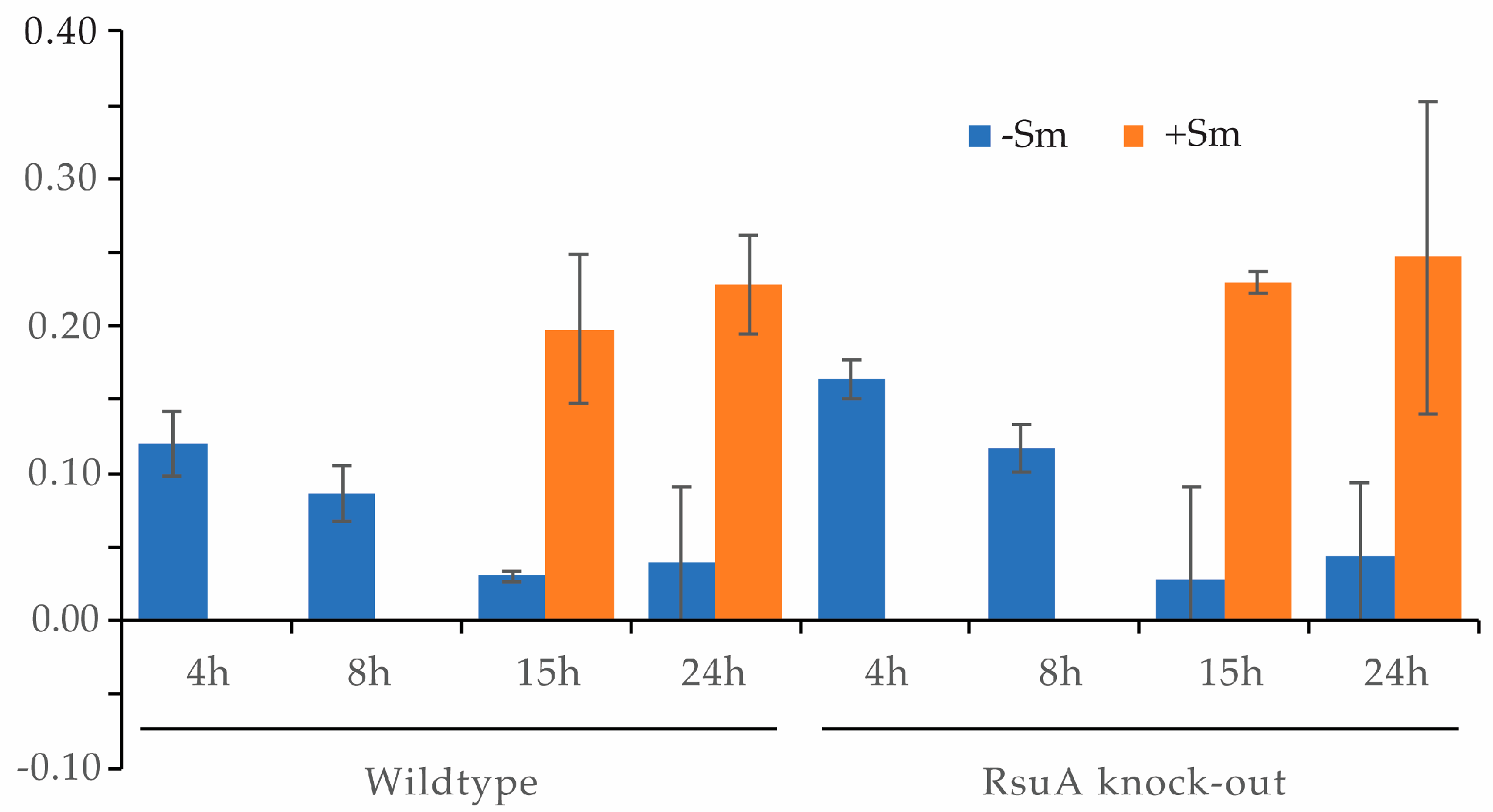
2.2. RsuA Influences rRNA Maturation during 30S Ribosome Biogenesis
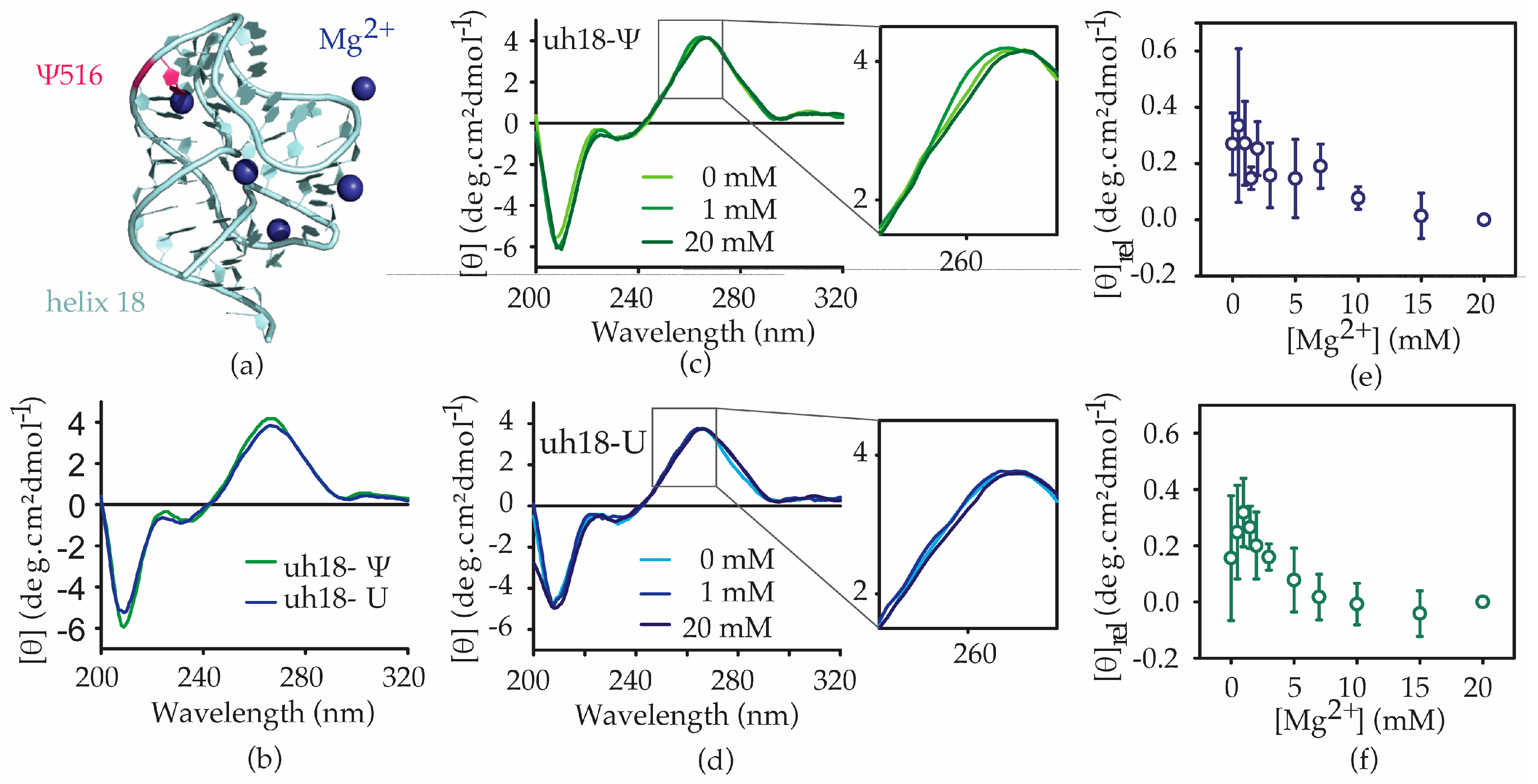
2.3. The Ψ516 Modification Influences the h18 Structure
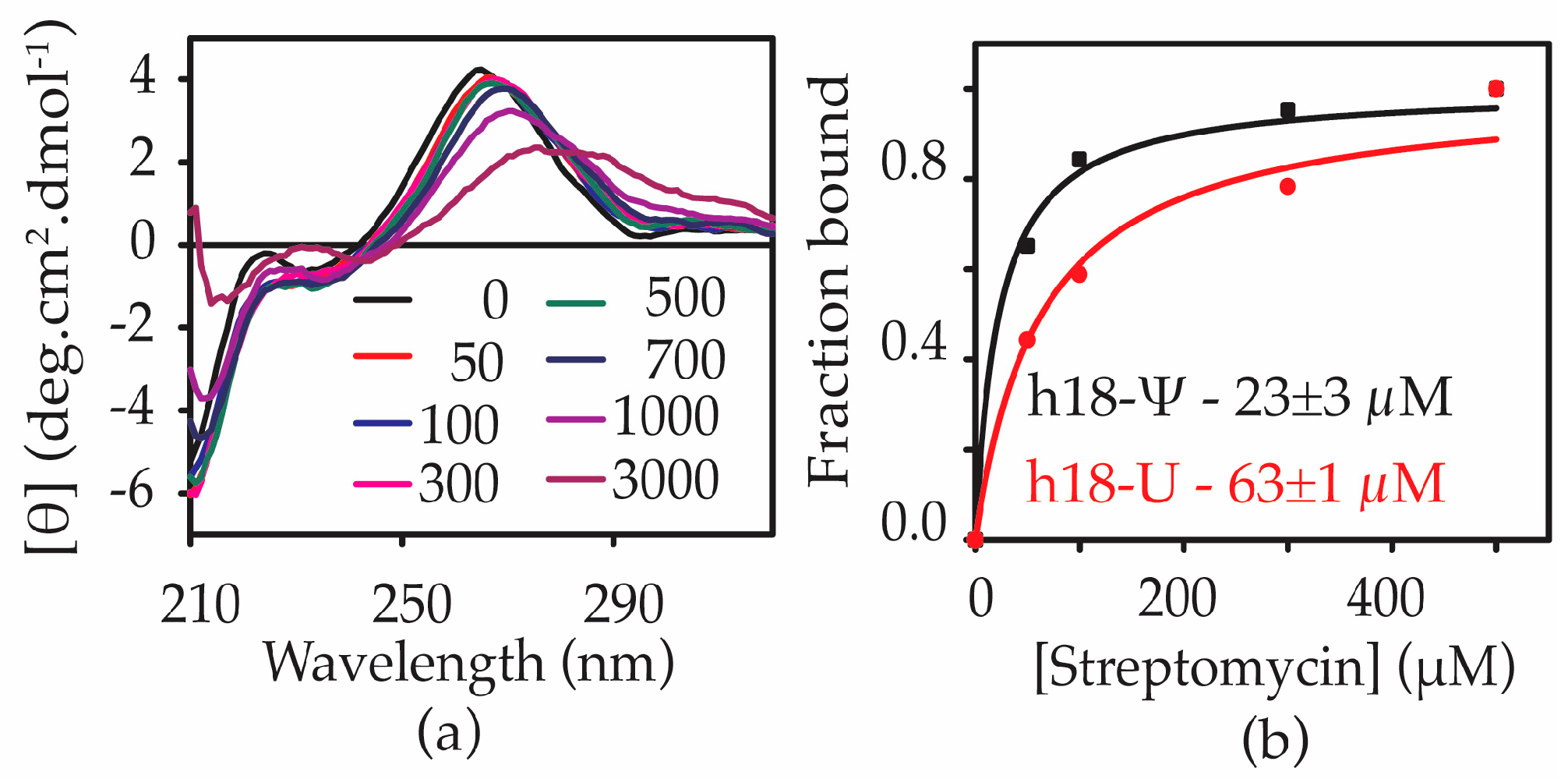
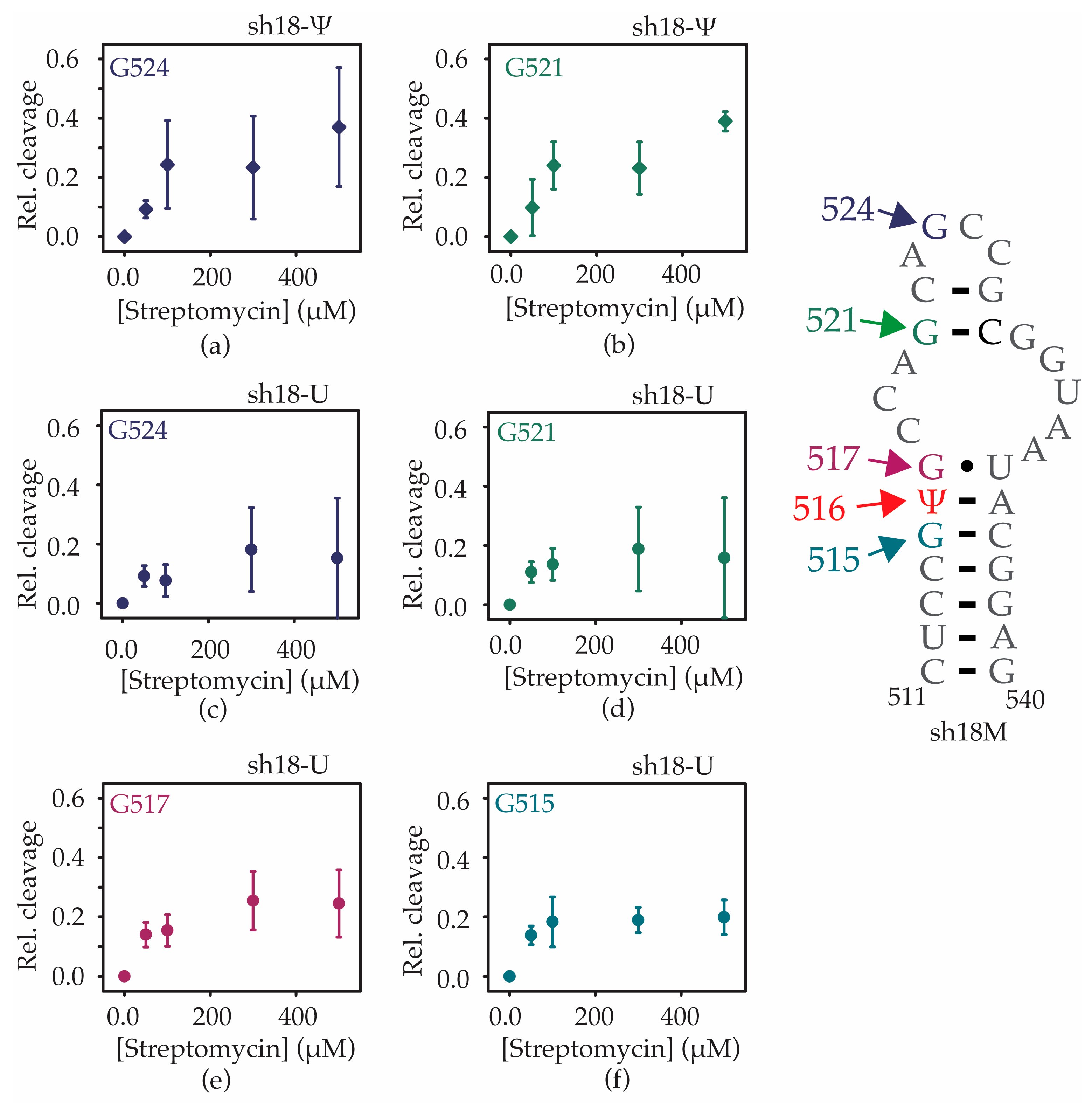
2.4. Ψ516 Modification Increases Streptomycin Binding to Helix 18
3. Discussion
4. Materials and Methods
4.1. Experimental Details Preparation of Helix 18 RNA
4.2. Circular Dichroism (CD) Spectroscopy
4.3. RNase Footprinting Experiments
4.4. Growth Inhibition Assays
4.5. Generation of Recombinant pCA24N Vector Encoding Catalytically Inactive RsuA
4.6. Generation of ΔRsuA + wtRsuA and ΔRsuA + mutRsuA E. coli Strains
4.7. Growth Curve Analysis
4.8. Non-Denaturing Gel Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chow, C.S.; Lamichhane, T.N.; Mahto, S.K. Expanding the Nucleotide Repertoire of the Ribosome with Post-Transcriptional Modifications. ACS Chem. Biol. 2007, 2, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Helm, M. Post-transcriptional nucleotide modification and alternative folding of RNA. Nucleic Acids Res. 2006, 34, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Douthwaite, S.; Fourmy, D.; Yoshizawa, S. Fine-Tuning of RNA Functions by Modification and Editing, 2nd ed.; Grosjean, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 12, pp. 285–307. [Google Scholar]
- Lázaro, E.; Rodriguez-Fonseca, C.; Porse, B.; Ureña, D.; Garrett, R.A.; Ballesta, J.P.G. A Sparsomycin-resistant Mutant of Halobacterium salinarium Lacks a Modification at Nucleotide U2603 in the Peptidyl Transferase Centre of 23 S rRNA. J. Mol. Biol. 1996, 261, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.M.; Mankin, A.S. An indigenous post-transcriptional modification in the ribosomal peptidyl transferase center confers resistance to an array of protein synthesis inhibitors. J. Mol. Biol. 2008, 380, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Boehringer, D.; O’Farrell, H.C.; Rife, J.P.; Ban, N. Structural insights into methyltransferase KsgA function in 30S ribosomal subunit biogenesis. J. Biol. Chem. 2012, 287, 10453–10459. [Google Scholar] [CrossRef]
- Okamoto, S.; Tamaru, A.; Nakajima, C.; Nishimura, K.; Tanaka, Y.; Tokuyama, S.; Suzuki, Y.; Ochi, K. Loss of a conserved 7-methylguanosine modification in 16S rRNA confers low-level streptomycin resistance in bacteria. Mol. Microbiol. 2007, 63, 1096–1106. [Google Scholar] [CrossRef]
- Helser, T.L.; Davies, J.E.; Dahlberg, J.E. Mechanism of Kasugamycin Resistance in Escherichia coli. Nat. New Biol. 1972, 235, 6–9. [Google Scholar] [CrossRef]
- Wrzesinski, J.; Bakin, A.; Nurse, K.; Lane, B.G.; Ofengand, J. Purification, cloning, and properties of the 16S RNA pseudouridine 516 synthase from Escherichia coli. Biochemistry 1995, 34, 8904–8913. [Google Scholar] [CrossRef]
- Conrad, J.; Niu, L.; Rudd, K.; Lane, B.G.; Ofengand, J. 16S ribosomal RNA pseudouridine synthase RsuA of Escherichia coli: Deletion, mutation of the conserved Asp102 residue, and sequence comparison among all other pseudouridine synthases. RNA 1999, 5, 751–763. [Google Scholar] [CrossRef]
- Bakin, A.; Kowalak, J.A.; McCloskey, J.A.; Ofengand, J. The single pseudouridine residue in Escherichia coli 16S RNA is located at position 516. Nucleic Acids Res. 1994, 22, 3681–3684. [Google Scholar] [CrossRef]
- Sivaraman, J.; Sauvé, V.; Larocque, R.; Stura, E.A.; Schrag, J.D.; Cygler, M.; Matte, A. Structure of the 16S rRNA pseudouridine synthase RsuA bound to uracil and UMP. Nat. Struct. Mol. Biol. 2002, 9, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Jayalath, K.; Frisbie, S.; To, M.; Abeysirigunawardena, S. Pseudouridine Synthase RsuA Captures an Assembly Intermediate that Is Stabilized by Ribosomal Protein S17. Biomolecules 2020, 10, 841. [Google Scholar] [CrossRef] [PubMed]
- Amitai, S.; Kolodkin-Gal, I.; Hananya-Meltabashi, M.; Sacher, A.; Engelberg-Kulka, H. Escherichia coli MazF Leads to the Simultaneous Selective Synthesis of Both “Death Proteins” and “Survival Proteins”. PLoS Genet. 2009, 5, e1000390. [Google Scholar] [CrossRef] [PubMed]
- Funatsu, G.; Wittmann, H.G. Ribosomal proteins. 33. Location of amino-acid replacements in protein S12 isolated from Escherichia coli mutants resistant to streptomycin. J. Mol. Biol. 1972, 68, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Melancon, P.; Lemieux, C.; Brakier-Gingras, L. A mutation in the 530 loop of Escherichia coli 16S ribosomal RNA causes resistance to streptomycin. Nucleic Acids Res. 1988, 16, 9631–9639. [Google Scholar] [CrossRef]
- Frattali, A.L.; Flynn, M.K.; de Stasio, E.A.; Dahĺberg, A.E. Effects of mutagenesis of C912 in the streptomycin binding region of Escherichia coli 16S ribosomal RNA. Biochim. Biophys. Acta 1990, 1050, 27–33. [Google Scholar] [CrossRef]
- Pinard, R.; Payant, C.; Melançon, P.; Brakier-Gingras, L. The 5’ proximal helix of 16S rRNA is involved in the binding of streptomycin to the ribosome. FASEB J. 1993, 7, 173–176. [Google Scholar] [CrossRef]
- Nishimura, K.; Johansen, S.K.; Inaoka, T.; Hosaka, T.; Tokuyama, S.; Tahara, Y.; Okamoto, S.; Kawamura, F.; Douthwaite, S.; Ochi, K. Identification of the RsmG Methyltransferase Target as 16S rRNA Nucleotide G527 and Characterization of Bacillus subtilis rsmG Mutants. J. Bacteriol. 2007, 189, 6068–6073. [Google Scholar] [CrossRef]
- Powers, T.; Noller, H.F. Selective perturbation of G530 of 16 S rRNA by translational miscoding agents and a streptomycin-dependence mutation in protein S12. J. Mol. Biol. 1994, 235, 156–172. [Google Scholar] [CrossRef]
- Hall, K.B.; McLaughlin, L.W. Properties of pseudouridine N1 imino protons located in the major groove of an A-form RNA duplex. Nucleic Acids Res. 1992, 20, 1883–1889. [Google Scholar] [CrossRef]
- Arnez, J.G.; Steitz, T.A. Crystal Structure of Unmodified tRNAGln Complexed with Glutaminyl-tRNA Synthetase and ATP Suggests a Possible Role for Pseudo-Uridines in Stabilization of RNA Structure. Biochemistry 1994, 33, 7560–7567. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.R. Stabilization of RNA stacking by pseudouridine. Nucleic Acids Res. 1995, 23, 5020–5026. [Google Scholar] [CrossRef]
- Abeysirigunawardena, S.C.; Chow, C.S. pH-dependent structural changes of helix 69 from Escherichia coli 23S ribosomal RNA. RNA 2008, 14, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, Y.; Abeysirigunawardena, S.C.; Duc, A.-C.E.; Dremann, D.N.; Chow, C.S. Ligand- and pH-Induced Conformational Changes of RNA Domain Helix 69 Revealed by 2-Aminopurine Fluorescence. Angew. Chem. Int. Ed. Engl. 2012, 51, 12095–12098. [Google Scholar] [CrossRef] [PubMed]
- Meroueh, M.; Grohar, P.J.; Qiu, J.; SantaLucia, J., Jr.; Scaringe, S.A.; Chow, C.S. Unique structural and stabilizing roles for the individual pseudouridine residues in the 1920 region of Escherichia coli 23S rRNA. Nucleic Acids Res. 2000, 28, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Wimberly, B.; Brodersen, D.; Clemons, W.; Morgan-Warren, R.; Carter, A.; Vonrhein, C.; Hartsch, T.; Ramakrishnan, V. Structure of the 30S Ribosomal Subunit. Nature 2000, 407, 327–339. [Google Scholar] [CrossRef]
- Adilakshmi, T.; Ramaswamy, P.; Woodson, S.A. Protein-independent Folding Pathway of the 16S rRNA 5′ Domain. J. Mol. Biol. 2005, 351, 508–519. [Google Scholar] [CrossRef]
- Woese, C.R.; Magrum, L.J.; Gupta, R.; Siegel, R.B.; Stahl, D.A.; Kop, J.; Crawford, N.; Brosius, J.; Gutell, R.; Hogan, J.J.; et al. Secondary structure model for bacterial 16S ribosomal RNA: Phylogenetic, enzymatic, and chemical evidence. Nucleic Acids Res. 1980, 8, 2275–2293. [Google Scholar] [CrossRef]
- Carter, A.P.; Clemons, W.M.; Brodersen, D.E.; Morgan-Warren, R.J.; Wimberly, B.T.; Ramakrishnan, V. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 2000, 407, 340–348. [Google Scholar] [CrossRef]
- Demirci, H.; Murphy, F.; Murphy, E.; Gregory, S.T.; Dahlberg, A.E.; Jogl, G. A structural basis for streptomycin-induced misreading of the genetic code. Nat. Commun. 2013, 4, 1355. [Google Scholar] [CrossRef]
- O’Connor, M.; Leppik, M.; Remme, J. Pseudouridine-Free Escherichia coli Ribosomes. J. Bacteriol. 2021, 200, e00540-17. [Google Scholar] [CrossRef]
- Green, R.; Noller, H.F. In vitro complementation analysis localizes 23S rRNA post-transcriptional modifications that are required for Escherichia coli 50S ribosomal subunit assembly and function. RNA 1996, 2, 1011–1021. [Google Scholar] [PubMed]
- Tollervey, D.; Lehtonen, H.; Jansen, R.; Kern, H.; Hurt, E.C. Temperature-sensitive mutations demonstrate roles for yeast fibrillarin in pre-rRNA processing, pre-rRNA methylation, and ribosome assembly. Cell 1993, 72, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Krzyzosiak, W.; Denman, R.; Nurse, K.; Hellmann, W.; Boublik, M.; Gehrke, C.W.; Agris, P.F.; Ofengand, J. In vitro synthesis of 16S ribosomal RNA containing single base changes and assembly into a functional 30S ribosome. Biochemistry 1987, 26, 2353–2364. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knock-out mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef]
- Li, B.; Qiu, Y.; Shi, H.; Yin, H. The importance of lag time extension in determining bacterial resistance to antibiotics. Analyst 2016, 141, 3059–3067. [Google Scholar] [CrossRef] [PubMed]
- Garvin, R.T.; Rosset, R.; Gorini, L. Ribosomal assembly influenced by growth in the presence of streptomycin. Proc. Natl. Acad. Sci. USA 1973, 70, 2762–2766. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhu, M.; Warren, M.; Balakrishnan, R.; Patsalo, V.; Okano, H.; Williamson, J.R.; Fredrick, K.; Wang, Y.-P.; Hwa, T. Reduction of translating ribosomes enables Escherichia coli to maintain elongation rates during slow growth. Nat. Microbiol. 2016, 2, 16231. [Google Scholar] [CrossRef]
- Powers, T.; Noller, H.F. A functional pseudoknot in 16S ribosomal RNA. EMBO J. 1991, 10, 2203–2214. [Google Scholar] [CrossRef]
- Ogle, J.M.; Ramakrishnan, V. Structural insights into translational fidelity. Annu. Rev. Biochem. 2005, 74, 129–177. [Google Scholar] [CrossRef]
- GC, K.; Gyawali, P.; Balci, H.; Abeysirigunawardena, S. Ribosomal RNA Methyltransferase RsmC Moonlights as an RNA Chaperone. ChemBioChem 2020, 21, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Meier, A.; Kirschner, P.; Bange, F.C.; Vogel, U.; Böttger, E.C. Genetic alterations in streptomycin-resistant Mycobacterium tuberculosis: Mapping of mutations conferring resistance. Antimicrob. Agents Chemother. 1994, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.T.; Cate, J.H.D.; Dahlberg, A.E. Streptomycin-resistant and streptomycin-dependent mutants of the extreme thermophile Thermus thermophilus. J. Mol. Biol. 2001, 309, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.T.; Carr, J.F.; Dahlberg, A.E. A Mutation in the Decoding Center of Thermus thermophilus 16S rRNA Suggests a Novel Mechanism of Streptomycin Resistance. J. Bacteriol. 2005, 187, 2200–2202. [Google Scholar] [CrossRef]
- Abedeera, S.M.; Hawkins, C.M.; Abeysirigunawardena, S.C. RsmG forms stable complexes with premature small subunit rRNA during bacterial ribosome biogenesis. RSC Adv. 2020, 10, 22361–22369. [Google Scholar] [CrossRef]
- Byrgazov, K.; Vesper, O.; Moll, I. Ribosome heterogeneity: Another level of complexity in bacterial translation regulation. Curr. Opin. Microbiol. 2013, 16, 133–139. [Google Scholar] [CrossRef]
- Vesper, O.; Amitai, S.; Belitsky, M.; Byrgazov, K.; Kaberdina, A.C.; Engelberg-Kulka, H.; Moll, I. Selective translation of leaderless mRNAs by specialized ribosomes generated by MazF in Escherichia coli. Cell 2011, 147, 147–157. [Google Scholar] [CrossRef]
- Kaberdina, A.; Szaflarski, W.; Nierhaus, K.; Moll, I. An Unexpected Type of Ribosomes Induced by Kasugamycin: A Look into Ancestral Times of Protein Synthesis? Mol. Cell 2009, 33, 227–236. [Google Scholar] [CrossRef]
- Kitagawa, M.; Ara, T.; Arifuzzaman, M.; Ioka-Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Mori, H. Complete set of ORF clones of Escherichia coli ASKA library (A Complete Set of E. coli K-12 ORF Archive): Unique Resources for Biological Research. DNA Res. 2005, 12, 291–299. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abedeera, S.M.; Jayalath, K.S.; Xie, J.; Rauff, R.M.; Abeysirigunawardena, S.C. Pseudouridine Synthase RsuA Confers a Survival Advantage to Bacteria under Streptomycin Stress. Antibiotics 2023, 12, 1447. https://doi.org/10.3390/antibiotics12091447
Abedeera SM, Jayalath KS, Xie J, Rauff RM, Abeysirigunawardena SC. Pseudouridine Synthase RsuA Confers a Survival Advantage to Bacteria under Streptomycin Stress. Antibiotics. 2023; 12(9):1447. https://doi.org/10.3390/antibiotics12091447
Chicago/Turabian StyleAbedeera, Sudeshi M., Kumudie S. Jayalath, Jiale Xie, Rushdhi M. Rauff, and Sanjaya C. Abeysirigunawardena. 2023. "Pseudouridine Synthase RsuA Confers a Survival Advantage to Bacteria under Streptomycin Stress" Antibiotics 12, no. 9: 1447. https://doi.org/10.3390/antibiotics12091447
APA StyleAbedeera, S. M., Jayalath, K. S., Xie, J., Rauff, R. M., & Abeysirigunawardena, S. C. (2023). Pseudouridine Synthase RsuA Confers a Survival Advantage to Bacteria under Streptomycin Stress. Antibiotics, 12(9), 1447. https://doi.org/10.3390/antibiotics12091447