

Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
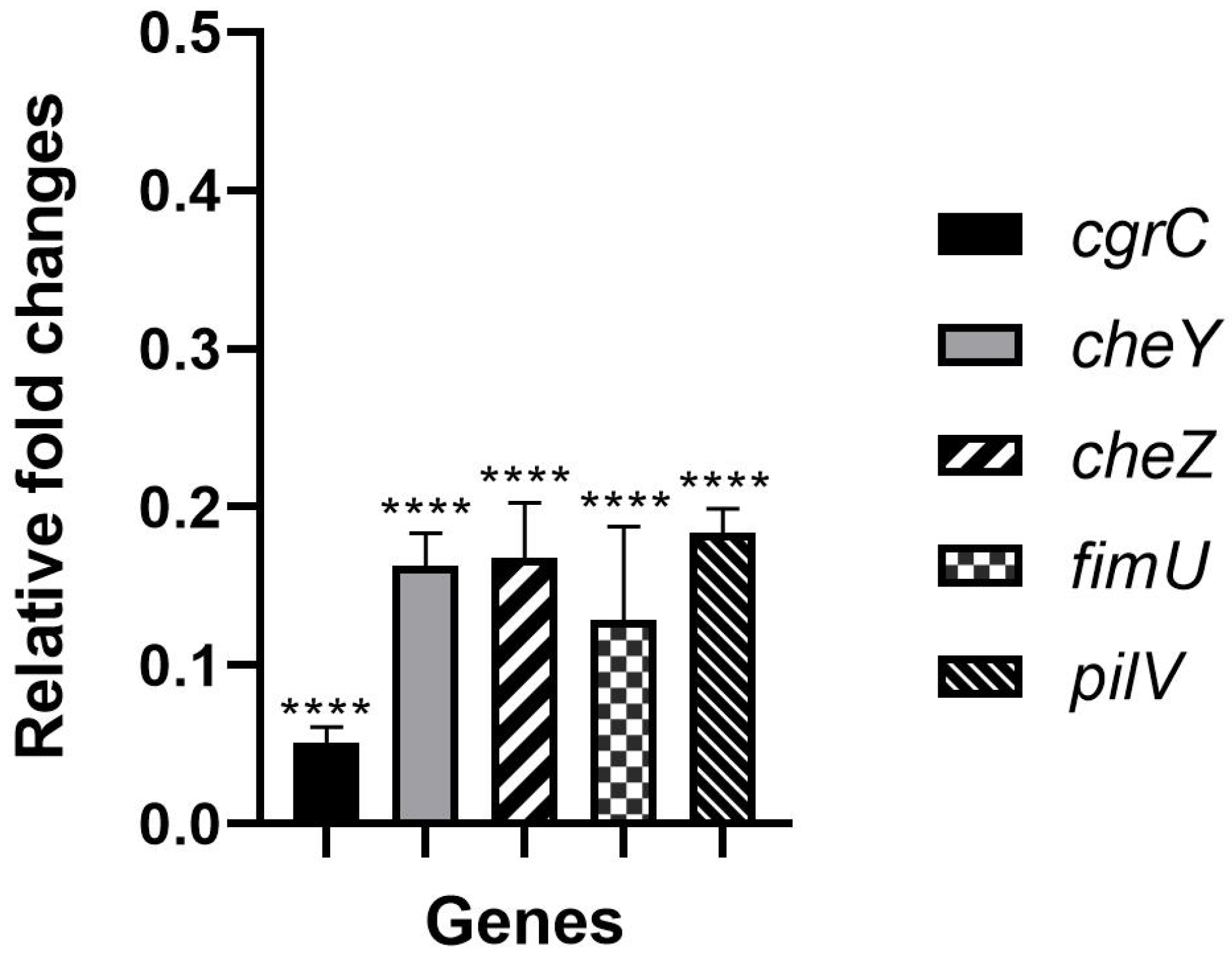
2.1. Hydroquinine Inhibits P. aeruginosa Growth through Decreased Expression Levels of Adhesion-Related Genes
2.2. MPSs Inhibit P. aeruginosa Growth Using the ISO 14729 Criteria
2.3. MPSs at Half Its Original Concentration Also Reduce P. aeruginosa Adhesion
2.4. Hydroquinine Shows Comparable Inhibitory Activity to MPSs, and When Used in Combination, Damages Cell Structure
2.5. Hydroquinine with Commercial MPS Demonstrates the Synergistic Effect to Reduce P. aeriginosa Adhesion on Contact Lens Surface
2.6. MPS Formulations Containing Hydroquinine Reduce Biofilm Mass on Contact Lens Surfaces
3. Discussion
4. Materials and Methods
4.1. P. aeruginosa Strains, Cultivation, and Inoculum Preparation
4.2. Contact Lenses and Lens Cases
4.3. Commercial Multipurpose Solutions
4.4. Solution Preparation
4.5. Studying Gene Expression Levels
4.5.1. RNA Extraction
4.5.2. Complementary DNA (cDNA) Synthesis
4.5.3. Quantitative Reverse Transcription PCR (RT-qPCR)
4.6. Stand-Alone Testing with Microorganisms
4.7. Anti-Adhesion Efficacy of Tested Solutions
4.8. Anti-Adhesion Efficacy on Contact Lens
4.9. Destruction of Biofilm on Contact Lens
4.10. Morphological Observations Using the FE-SEM
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreddu, R.; Vigolo, D.; Yetisen, A.K. Contact lens technology: From fundamentals to applications. Adv. Healthc. Mater. 2019, 8, e1900368. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.H.L.; Stapleton, F.; Mehta, J.S. Review of Contact Lens-Related Complications. Eye Contact Lens 2018, 44 (Suppl. S2), S1–S10. [Google Scholar] [CrossRef] [PubMed]
- Forister, J.F.; Forister, E.F.; Yeung, K.K.; Ye, P.; Chung, M.Y.; Tsui, A.; Weissman, B.A. Prevalence of contact lens-related complications: UCLA contact lens study. Eye Contact Lens 2009, 35, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Eltis, M. Contact-lens-related microbial keratitis: Case report and review. J. Optom. 2011, 4, 122–127. [Google Scholar] [CrossRef]
- Dart, J.K.G.; Stapleton, F.; Minassian, D.; Dart, J.K.G. Contact lenses and other risk factors in microbial keratitis. Lancet 1991, 338, 650–653. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Carnt, N. Contact lens-related microbial keratitis: How have epidemiology and genetics helped us with pathogenesis and prophylaxis. Eye 2012, 26, 185–193. [Google Scholar] [CrossRef]
- Cohen, E.J.; Laibson, P.R.; Arentsen, J.J.; Clemons, C.S. Corneal ulcers associated with cosmetic extended wear soft contact lenses. Ophthalmology 1987, 94, 109–114. [Google Scholar] [CrossRef]
- Preechawat, P.; Ratananikom, U.; Lerdvitayasakul, R.; Kunavisarut, S. Contact lens-related microbial keratitis. J. Med. Assoc. Thai. 2007, 90, 737–743. [Google Scholar]
- Green, M.; Apel, A.; Stapleton, F. Risk factors and causative organisms in microbial keratitis. Cornea 2008, 27, 22–27. [Google Scholar] [CrossRef]
- Al-Mujaini, A.; Al-Kharusi, N.; Thakral, A.; Wali, U.K. Bacterial keratitis: Perspective on epidemiology, clinico-pathogenesis, diagnosis and treatment. Sultan Qaboos Univ. Med. J. 2009, 9, 184–195. [Google Scholar]
- Robaei, D.; Watson, S. Corneal blindness: A global problem. Clin. Exp. Ophthalmol. 2014, 42, 213–214. [Google Scholar] [CrossRef] [PubMed]
- Resnikoff, S.; Pascolini, D.; Etya’ale, D.; Kocur, I.; Pararajasegaram, R.; Pokharel, G.P.; Mariotti, S.P. Global data on visual impairment in the year 2002. Bull. World Health Organ. 2004, 82, 844–851. [Google Scholar] [PubMed]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014.
- Choy, M.H.; Stapleton, F.; Willcox, M.D.P.; Zhu, H. Comparison of virulence factors in Pseudomonas aeruginosa strains isolated from contact lens- and non-contact lens-related keratitis. J. Med. Microbiol. 2008, 57, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Ozer, E.; Yaniv, K.; Chetrit, E.; Boyarski, A.; Meijler, M.M.; Berkovich, R.; Kushmaro, A.; Alfonta, L. An inside look at a biofilm: Pseudomonas aeruginosa flagella biotracking. Sci. Adv. 2021, 7, eabg8581. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Borlace, L.; Stapleton, F.; Matheson, M.; Dart, J.K. Bacterial biofilm on contact lenses and lens storage cases in wearers with microbial keratitis. J. Appl. Microbiol. 1998, 84, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13. [Google Scholar] [CrossRef] [PubMed]
- ISO 14729:2001; Ophthalmic Optics—Contact Lens Care Products—Microbiological Requirements and Test Methods for Products and Regimens for Hygienic Management of Contact Lenses. ISO: Geneva, Switzerland, 2001.
- McAnally, C.; Walters, R.; Campolo, A.; Harris, V.; King, J.; Thomas, M.; Gabriel, M.M.; Shannon, P.; Crary, M. Antimicrobial efficacy of contact lens solutions assessed by ISO standards. Microorganisms 2021, 9, 2173. [Google Scholar] [CrossRef]
- Szczotka-Flynn, L.B.; Imamura, Y.; Chandra, J.; Yu, C.; Mukherjee, P.K.; Pearlman, E.; Ghannoum, M.A. Increased resistance of contact lens-related bacterial biofilms to antimicrobial activity of soft contact lens care solutions. Cornea 2009, 28, 918–926. [Google Scholar] [CrossRef]
- Kilvington, S.; Huang, L.; Kao, E.; Powell, C.H. Development of a new contact lens multipurpose solution: Comparative analysis of microbiological, biological and clinical performance. J. Optom. 2010, 3, 134–142. [Google Scholar] [CrossRef]
- Bispo, P.J.; Haas, W.; Gilmore, M.S. Biofilms in infections of the eye. Pathogens 2015, 4, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Biswas, N.R.; Gupta, S.K.; Das, G.K.; Kumar, N.; Mongre, P.K.; Haldar, D.; Beri, S. Evaluation of Ophthacare eye drops-a herbal formulation in the management of various ophthalmic disorders. Phytother. Res. 2001, 15, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Kraikongjit, S.; Jongjitvimol, T.; Mianjinda, N.; Sirithep, N.; Kaewbor, T.; Jumroon, N.; Jongjitwimol, J. Antibacterial effect of plant resin collected from Tetrigona apicalis (Smith, 1857) in Thung Salaeng Luang National Park, Phitsanulok. Walailak J. Sci. Technol. 2017, 15, 599–607. [Google Scholar] [CrossRef]
- Jongjitvimol, T.; Kraikongjit, S.; Paensuwan, P.; Jongjitwimol, J. In vitro biological profiles and chemical contents of ethanolic nest entrance extracts of thai stingless bees Tetrigona apicalis. J. Biol. Sci. 2020, 20, 157–165. [Google Scholar] [CrossRef]
- Nationnal Center for Advancing Translation Science; NIH. Inxignt: Drugs, Hydroquinine 31J30Q51T6L. Available online: https://drugs.ncats.io/substance/31J3Q51T6L (accessed on 1 September 2023).
- National Health Care Institute. Hydroquinine (Inhibin®) for Patients with Nocturnal Muscle Cramps. Available online: https://english.zorginstituutnederland.nl/publications/reports/2020/03/04/hydroquinine-inhibin-for-patients-with-nocturnal-muscle-cramps (accessed on 1 September 2023).
- Rattanachak, N.; Weawsiangsang, S.; Jongjitvimol, T.; Baldock, R.A.; Jongjitwimol, J. Hydroquinine possesses antibacterial activity, and at half the MIC, induces the overexpression of RND-type efflux pumps using multiplex digital PCR in Pseudomonas aeruginosa. Trop. Med. Infect. Dis. 2022, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Jongjitwimol, J.; Baldock, R.A. Hydroquinine: A potential new avenue in drug discovery for drug-resistant bacteria? Expert Opin. Drug Discov. 2023, 18, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Weawsiangsang, S.; Rattanachak, N.; Jongjitvimol, T.; Jaifoo, T.; Charoensit, P.; Viyoch, J.; Ross, S.; Ross, G.M.; Baldock, R.A.; Jongjitwimol, J. Hydroquinine inhibits the growth of multidrug-resistant Pseudomonas aeruginosa via the suppression of the arginine deiminase pathway genes. Int. J. Mol. Sci. 2023, 24, 13914. [Google Scholar] [CrossRef] [PubMed]
- Rattanachak, N.; Weawsiangsang, S.; Daowtak, K.; Thongsri, Y.; Ross, S.; Ross, G.; Nilsri, N.; Baldock, R.A.; Pongcharoen, S.; Jongjitvimol, T.; et al. High-throughput transcriptomic profiling reveals the inhibitory effect of hydroquinine on virulence factors in Pseudomonas aeruginosa. Antibiotics 2022, 11, 1436. [Google Scholar] [CrossRef]
- Conrad, J.C.; Gibiansky, M.L.; Jin, F.; Gordon, V.D.; Motto, D.A.; Mathewson, M.A.; Stopka, W.G.; Zelasko, D.C.; Shrout, J.D.; Wong, G.C. Flagella and pili-mediated near-surface single-cell motility mechanisms in P. aeruginosa. Biophys. J. 2011, 100, 1608–1616. [Google Scholar] [CrossRef]
- Berg, H.C. The rotary motor of bacterial flagella. Annu. Rev. Biochem. 2003, 72, 19–54. [Google Scholar] [CrossRef]
- Craig, L.; Li, J. Type IV pili: Paradoxes in form and function. Curr. Opin. Struct. Biol. 2008, 18, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Vallet-Gely, I.; Sharp, J.S.; Dove, S.L. Local and global regulators linking anaerobiosis to cupA fimbrial gene expression in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 8667–8676. [Google Scholar] [CrossRef] [PubMed]
- Giraud, C.; de Bentzmann, S. Inside the complex regulation of Pseudomonas aeruginosa chaperone usher systems. Environ. Microbiol. 2012, 14, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Vallet, I.; Olson, J.W.; Lory, S.; Lazdunski, A.; Filloux, A. The chaperone/usher pathways of Pseudomonas aeruginosa: Identification of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc. Natl. Acad. Sci. USA 2001, 98, 6911–6916. [Google Scholar] [CrossRef] [PubMed]
- McManus Heather, R.; Dove Simon, L. The CgrA and CgrC proteins form a complex that positively regulates cupA fimbrial gene expression in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 6152–6161. [Google Scholar] [CrossRef]
- Kuo, S.C.; Koshland, D.E., Jr. Roles of cheY and cheZ gene products in controlling flagellar rotation in bacterial chemotaxis of Escherichia coli. J. Bacteriol. 1987, 169, 1307–1314. [Google Scholar] [CrossRef]
- Sampedro, I.; Parales, R.E.; Krell, T.; Hill, J.E. Pseudomonas chemotaxis. FEMS Microbiol. Rev. 2015, 39, 17–46. [Google Scholar] [CrossRef]
- Han, X.; Kennan, R.M.; Parker, D.; Davies, J.K.; Rood, J.I. Type IV fimbrial biogenesis is required for protease secretion and natural transformation in Dichelobacter nodosus. J. Bacteriol. 2007, 189, 5022–5033. [Google Scholar] [CrossRef]
- Giltner, C.L.; Nguyen, Y.; Burrows, L.L. Type IV pilin proteins: Versatile molecular modules. Microbiol. Mol. Biol. Rev. 2012, 76, 740–772. [Google Scholar] [CrossRef]
- Alm, R.A.; Mattick, J.S. Identification of a gene, pilV, required for type 4 fimbrial biogenesis in Pseudomonas aeruginosa, whose product possesses a pre-pilin-like leader sequence. Mol. Microbiol. 1995, 16, 485–496. [Google Scholar] [CrossRef]
- Strom, M.S.; Nunn, D.N.; Lory, S. A single bifunctional enzyme, PilD, catalyzes cleavage and N-methylation of proteins belonging to the type IV pilin family. Proc. Natl. Acad. Sci. USA 1993, 90, 2404–2408. [Google Scholar] [CrossRef] [PubMed]
- Alm, R.A.; Mattick, J.S. Identification of two genes with prepilin-like leader sequences involved in type 4 fimbrial biogenesis in Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 3809–3817. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef]
- Chiang, P.; Habash, M.; Burrows, L.L. Disparate subcellular localization patterns of Pseudomonas aeruginosa type IV pilus ATPases involved in twitching motility. J. Bacteriol. 2005, 187, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Codling, C.E.; Maillard, J.Y.; Russell, A.D. Aspects of the antimicrobial mechanisms of action of a polyquaternium and an amidoamine. J. Antimicrob. Chemother. 2003, 51, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.A.; Joshi, L.T. Biocide use in the antimicrobial era: A review. Molecules 2021, 26, 2276. [Google Scholar] [CrossRef]
- Salton, M.R. Lytic agents, cell permeability, and monolayer penetrability. J. Gen. Physiol. 1968, 52, 227–252. [Google Scholar] [CrossRef]
- de Azevedo Magalhães, O.; Ribeiro Dos Santos, D.; Coch Broetto, B.G.; Corção, G. Polyhexamethylene biguanide multipurpose solutions on bacterial disinfection: A comparison study of effectiveness in a developing country. Eye Contact Lens 2023, 49, 139–142. [Google Scholar] [CrossRef]
- Bradley, C.S.; Sicks, L.A.; Pucker, A.D. Common ophthalmic preservatives in soft contact lens care products: Benefits, complications, and a comparison to non-preserved solutions. Clin. Optom. 2021, 13, 271–285. [Google Scholar] [CrossRef]
- ISO 18259: 2001/2014; Ophthalmic Optics—Contact Lens Care Products—Method to Assess Contact Lens Care Products with Contact Lenses in a Lens Case, Challenged with Bacterial and Fungal Organisms. ISO: Geneva, Switzerland, 2014.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Solutions | P. aeruginosa ATCC 27853 | Clinical P. aeruginosa Strain |
|---|---|---|
| PBS | 00.00 ± 0.54 | 00.00 ± 0.63 |
| 50% MPS A | 89.87 ± 0.03 *,a | 89.76 ± 0.07 *,a |
| 100% MPS A | 91.89 ± 0.19 *,b | 91.37 ± 0.08 *,d |
| 50% MPS B | 75.30 ± 1.36 *,a,c | 75.05 ± 1.58 *,a,c |
| 100% MPS B | 83.54 ± 1.10 *,b,c | 86.83 ± 0.90 *,c,d |
| Tested Solutions | P. aeruginosa ATCC 27853 | Clinical P. aeruginosa Strain |
|---|---|---|
| PBS | 00.00 ± 0.54 | 00.00 ± 0.63 |
| HQ [1.25] in PBS | 57.80 ± 0.41 *,A | 54.84 ± 3.76 *,C |
| HQ [1.25] with 50% MPS A | 95.59 ± 0.13 *,a | 92.39 ± 0.55 *,c |
| HQ [1.25] with 100% MPS A | 97.91 ± 0.21 *,a | 93.64 ± 0.17 *,c |
| HQ [2.50] in PBS | 59.56 ± 0.34 *,B | 56.37 ± 1.27 *,D |
| HQ [2.50] with 50% MPS A | 96.49 ± 0.22 *,b | 92.23 ± 0.16 *,d |
| HQ [2.50] with 100% MPS A | 97.16 ± 0.03 *,b | 93.31 ± 0.20 *,d |
| Code | MPS A | MPS B |
|---|---|---|
| Product | Opti-free® Replenish® solution | Q-eye multipurpose solution |
| Manufacturer | Alcon Laboratories, Inc., Fort Worth, TX, USA | Stericon Pharma Pvt. Ltd., Karnataka, India |
| Disinfectants and preservatives | polyquaternium-1—POLYQUAD® 0.001%, myristamidopropyl dimethylamine—ALDOX® 0.0005% | polyhexamethylene biguanide (PHMB) 0.0001% |
| Wettings agents | poloxamine—TETRONIC® 1304 | poloxamer, dexpanthenol |
| Lubricants | propylene glycol | sorbitol |
| Buffer and saline | sodium citrate, sodium borate, sodium chloride | disodium edetate, trometamol, sodium dihydrogen phosphate dihydrate |
| Genes | Primer Direction | Oligonucleotide Sequences (5′ to 3′) | Annealing Temperature (°C) | References |
|---|---|---|---|---|
| cgrC | Forward | CGAGCGGATTGAAGCCATC | 57 | This study |
| Reverse | ACGATGGGCTGGGTGAATC | This study | ||
| cheZ | Forward | ACTGGTGGACTGTCTCGAAC | 57 | This study |
| Reverse | CGATCTGCGACATTTCCTGC | This study | ||
| cheY | Forward | CCACGATGAGACGCATCATC | 56 | This study |
| Reverse | ATGTTCCAGTCGGTGACGAG | This study | ||
| fimU | Forward | GGAACTCAATGCGATGCTGC | 56 | This study |
| Reverse | GAAGGTCAGATGTTCCACGG | This study | ||
| pilV | Forward | ACGACGTCAAGGACCAGATG | 57 | This study |
| Reverse | GGCAGTTCGTTCTTCACCTG | This study | ||
| 16s rRNA | Forward | CATGGCTCAGATTGAACGCTG | 58 | [29] |
| Reverse | GCTAATCCGACCTAGGCTCATC | [29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weawsiangsang, S.; Rattanachak, N.; Ross, S.; Ross, G.M.; Baldock, R.A.; Jongjitvimol, T.; Jongjitwimol, J. Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation. Antibiotics 2024, 13, 56. https://doi.org/10.3390/antibiotics13010056
Weawsiangsang S, Rattanachak N, Ross S, Ross GM, Baldock RA, Jongjitvimol T, Jongjitwimol J. Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation. Antibiotics. 2024; 13(1):56. https://doi.org/10.3390/antibiotics13010056
Chicago/Turabian StyleWeawsiangsang, Sattaporn, Nontaporn Rattanachak, Sukunya Ross, Gareth M. Ross, Robert A. Baldock, Touchkanin Jongjitvimol, and Jirapas Jongjitwimol. 2024. "Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation" Antibiotics 13, no. 1: 56. https://doi.org/10.3390/antibiotics13010056
APA StyleWeawsiangsang, S., Rattanachak, N., Ross, S., Ross, G. M., Baldock, R. A., Jongjitvimol, T., & Jongjitwimol, J. (2024). Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation. Antibiotics, 13(1), 56. https://doi.org/10.3390/antibiotics13010056