Prevalence and Epidemiology of Multidrug-Resistant Pathogens in the Food Chain and the Urban Environment in Northwestern Germany


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of AMR Genes
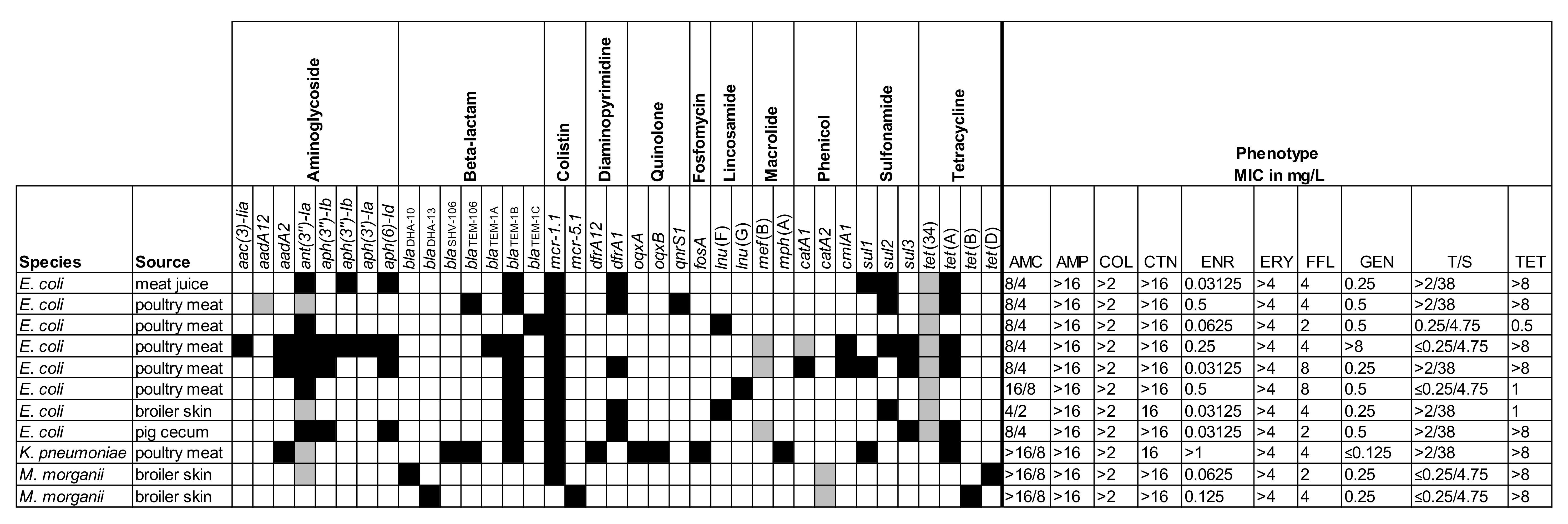
2.2. Genotyping of Selected Isolates
3. Discussion
4. Materials and Methods
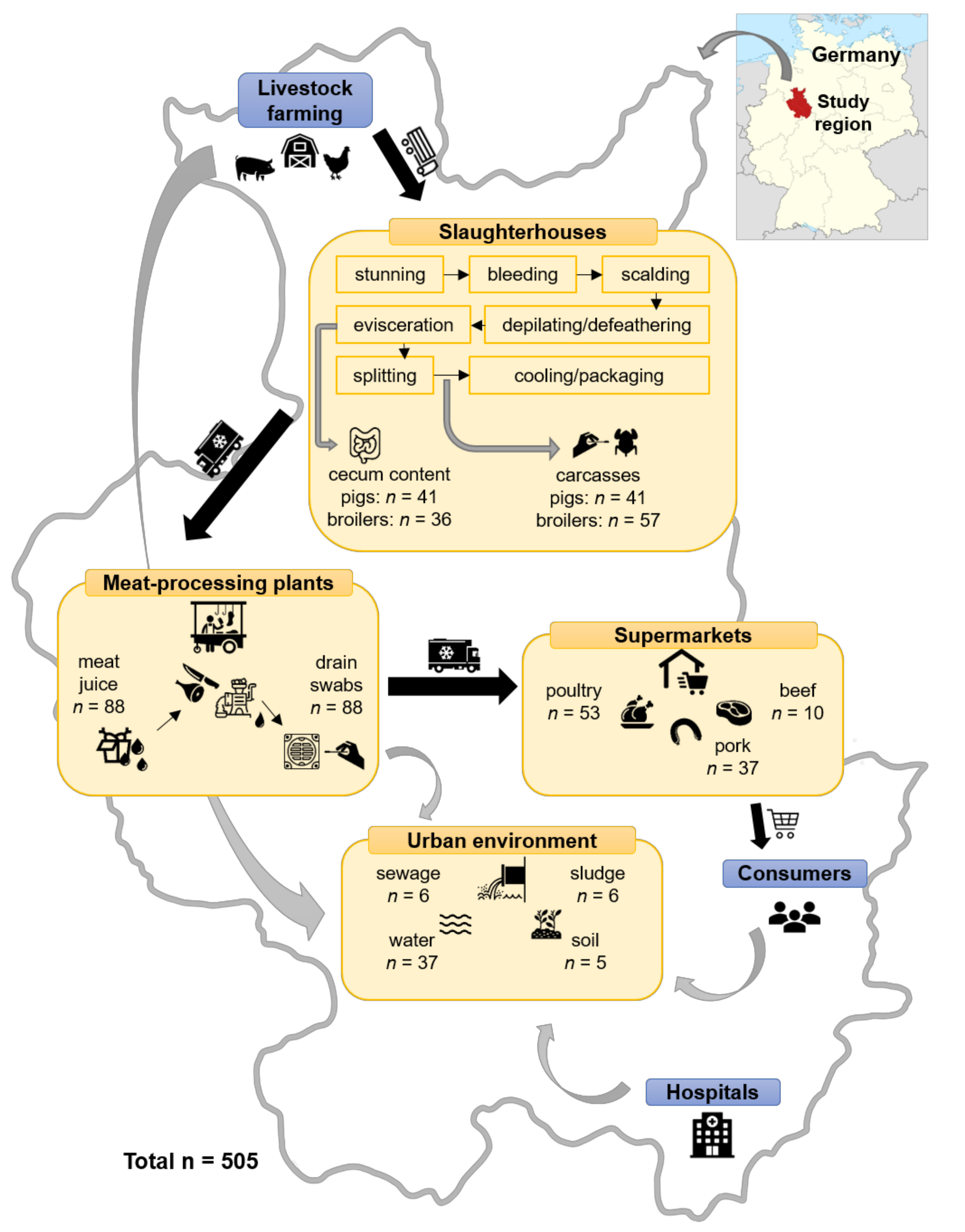
4.1. Sample Collection
4.2. Molecular and Culture-Based Screening for Col-E, CPE, VRE, and ESBL-Producers
4.3. Molecular Typing of Selected Isolates
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Assessing the antimicrobial susceptibility of bacteria obtained from animals. Vet. Microbiol. 2010, 141, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, P.M.; Loureiro, L.; Matos, A.J.F. Transfer of multidrug-resistant bacteria between intermingled ecological niches: The interface between humans, animals and the environment. Int. J. Environ. Res. Public Health 2013, 10, 278–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.; Hille, K.; Ruddat, I.; Mellmann, A.; Köck, R.; Kreienbrock, L. Simultaneous occurrence of MRSA and ESBL-producing Enterobacteriaceae on pig farms and in nasal and stool samples from farmers. Vet. Microbiol. 2017, 200, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Friese, A.; Schulz, J.; Laube, H.; von Salviati, C.; Hartung, J.; Roesler, U. Faecal occurrence and emissions of livestock-associated methicillin-resistant Staphylococcus aureus (laMRSA) and ESBL/AmpC-producing E. coli from animal farms in Germany. Berl. Münch. Tierärztl. Wochenschr. 2013, 126, 175–180. [Google Scholar]
- Beneke, B.; Klees, S.; Stührenberg, B.; Fetsch, A.; Kraushaar, B.; Tenhagen, B.A. Prevalence of methicillin-resistant Staphylococcus aureus in a fresh meat pork production chain. J. Food Prot. 2011, 74, 126–129. [Google Scholar] [CrossRef]
- Schulz, J.; Friese, A.; Klees, S.; Tenhagen, B.A.; Fetsch, A.; Rösler, U.; Hartung, J. Longitudinal study of the contamination of air and of soil surfaces in the vicinity of pig barns by livestock-associated methicillin-resistant Staphylococcus aureus. Appl. Environ. Microbiol. 2012, 78, 5666–5671. [Google Scholar] [CrossRef] [Green Version]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: A systematic review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschanski, N.; Falgenhauer, L.; Grobbel, M.; Guenther, S.; Kreienbrock, L.; Imirzalioglu, C.; Roesler, U. Retrospective survey of mcr-1 and mcr-2 in German pig-fattening farms, 2011–2012. Int. J. Antimicrob. Agents 2017, 50, 266–271. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Van Essen-Zandbergen, A.; Veldman, K.T.; Tafro, N.; Haenen, O.; Mevius, D.J. Chromosome-based blaOXA-48-like variants in Shewanella species isolates from food-producing animals, fish, and the aquatic environment. Antimicrob. Agents Chemother. 2017, 61, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; Johnson, A.P. Transferable resistance to colistin: A new but old threat. J. Antimicrob. Chemother. 2016, 71, 2066–2070. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, R.; Schwarz, S.; Wu, C.; Shen, J.; Walsh, T.R.; Wang, Y. Farm animals and aquaculture: Significant reservoirs of mobile colistin resistance genes. Environ. Microbiol. 2020, 22, 2469–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybysz, S.M.; Correa-Martinez, C.; Köck, R.; Becker, K.; Schaumburg, F. SuperPolymyxinTM medium for the screening of colistin-resistant gram-negative bacteria in stool samples. Front. Microbiol. 2018, 9, 2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remschmidt, C.; Schröder, C.; Behnke, M.; Gastmeier, P.; Geffers, C.; Kramer, T.S. Continuous increase of vancomycin resistance in enterococci causing nosocomial infections in Germany - 10 years of surveillance. Antimicrob. Resist. Infect. Control 2018, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Pauly, N.; Hammerl, J.A.; Grobbel, M.; Tenhagen, B.A.; Käsbohrer, A.; Bisenius, S.; Fuchs, J.; Horlacher, S.; Lingstädt, H.; Mauermann, U.; et al. ChromID® CARBA agar fails to detect carbapenem-resistant Enterobacteriaceae with slightly reduced susceptibility to carbapenems. Front. Microbiol. 2020, 11, 1678. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Sib, E.; Gajdiss, M.; Klanke, U.; Lenz-Plet, F.; Barabasch, V.; Albert, C.; Schallenberg, A.; Timm, C.; Zacharias, N.; et al. Dissemination of multi-resistant Gram-negative bacteria into German wastewater and surface waters. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a novel mcr-6 to mcr-9 multiplex PCR and assessment of mcr-1 to mcr-9 occurrence in colistin-resistant salmonella enterica isolates from environment, feed, animals and food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Irrgang, A.; Roschanski, N.; Tenhagen, B.A.; Grobbel, M.; Skladnikiewicz-Ziemer, T.; Thomas, K.; Roesler, U.; Käsbohrer, A. Prevalence of mcr-1 in E. coli from livestock and food in Germany, 2010-2015. PLoS ONE 2016, 11, e0159863. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, J.; Hu, Q.; Rao, X. Morganella morganii, a non-negligent opportunistic pathogen. Int. J. Infect. Dis. 2016, 50, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Bundesministerium für Ernährung und Landwirtschaft (BMEL); Bundesamt für Verbraucherschutz und Lebensmittelsicherheit (BVL); Bundesinstitut für Risikobewertung (BfR). Lagebild zur Antibiotikaresistenz im Bereich Tierhaltung und Lebensmittelkette. 2018. Available online: https://www.bmel.de/SharedDocs/Downloads/DE/_Tiere/Tiergesundheit/Tierarzneimittel/Lagebild-Antibiotikaeinsatz-bei-Tieren-Juli-2018.pdf?__blob=publicationFile&v=2 (accessed on 17 September 2020).
- Moennighoff, C.; Thomas, N.; Nienhaus, F.; Hartmann, M.; Menrath, A.; Merkel, J.; Detlefsen, H.; Kreienbrock, L.; Hennig-Pauka, I. Phenotypic antimicrobial resistance in Escherichia coli strains isolated from swine husbandries in North Western Germany - Temporal patterns in samples from laboratory practice from 2006 to 2017. BMC Vet. Res. 2020, 16, 37. [Google Scholar] [CrossRef]
- Schaumburg, F.; Sertic, S.M.; Correa-Martinez, C.; Mellmann, A.; Köck, R.; Becker, K. Acquisition and colonization dynamics of antimicrobial-resistant bacteria during international travel: A prospective cohort study. Clin. Microbiol. Infect. 2019, 25, 1287.e1–1287.e7. [Google Scholar] [CrossRef]
- Terveer, E.M.; Nijhuis, R.H.T.; Crobach, M.J.T.; Knetsch, C.W.; Veldkamp, K.E.; Gooskens, J.; Kuijper, E.J.; Claas, E.C.J. Prevalence of colistin resistance gene (mcr-1) containing Enterobacteriaceae in feces of patients attending a tertiary care hospital and detection of a mcr-1 containing, colistin susceptible E. coli. PLoS ONE 2017, 12, e0178598. [Google Scholar] [CrossRef] [Green Version]
- Saly, M.; Jayol, A.; Poirel, L.; Megraud, F.; Nordmann, P.; Dubois, V. Prevalence of faecal carriage of colistin-resistant gram-negative rods in a university hospital in Western France, 2016. J. Med. Microbiol. 2017, 66, 842–843. [Google Scholar] [CrossRef] [Green Version]
- Zurfluh, K.; Stephan, R.; Widmer, A.; Poirel, L.; Nordmann, P.; Nüesch, H.J.; Hächler, H.; Nüesch-Inderbinen, M. Screening for fecal carriage of MCR-producing Enterobacteriaceae in healthy humans and primary care patients. Antimicrob. Resist. Infect. Control 2017, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bich, V.T.N.; Thanh, L.V.; Thai, P.D.; Van Phuong, T.T.; Oomen, M.; Driessen, C.; Beuken, E.; Hoang, T.H.; Van Doorn, H.R.; Penders, J.; et al. An exploration of the gut and environmental resistome in a community in northern Vietnam in relation to antibiotic use. Antimicrob. Resist. Infect. Control 2019, 8, 194. [Google Scholar] [CrossRef]
- Giani, T.; Sennati, S.; Antonelli, A.; Di Pilato, V.; Di Maggio, T.; Mantella, A.; Niccolai, C.; Spinicci, M.; Monasterio, J.; Castellanos, P.; et al. High prevalence of carriage of mcr-1-positive enteric bacteria among healthy children from rural communities in the Chaco region, Bolivia, september to october 2016. Eurosurveillance 2018, 23, 1800115. [Google Scholar] [CrossRef]
- Trung, N.V.; Matamoros, S.; Carrique-Mas, J.J.; Nghia, N.H.; Nhung, N.T.; Chieu, T.T.B.; Mai, H.H.; van Rooijen, W.; Campbell, J.; Wagenaar, J.A.; et al. Zoonotic transmission of mcr-1 colistin resistance gene from small-scale poultry farms, Vietnam. Emerg. Infect. Dis. 2017, 23, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Dorado-García, A.; van Duijkeren, E.; van den Bunt, G.; Dierikx, C.M.; Bonten, M.J.M.; Bootsma, M.C.J.; Schmitt, H.; Hald, T.; Evers, E.G.; et al. Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: A population-based modelling study. Lancet Planet. Health 2019, 3, e357–e369. [Google Scholar] [CrossRef] [Green Version]
- Bager, F.; Madsen, M.; Christensen, J.; Aarestrup, F.M. Avoparcin used as a growth promoter is associated with the occurrence of vancomycin-resistant Enterococcus faecium on Danish poultry and pig farms. Prev. Vet. Med. 1997, 31, 95–112. [Google Scholar] [CrossRef]
- Klare, I.; Badstübner, D.; Konstabel, C.; Böhme, G.; Claus, H.; Witte, W. Decreased incidence of VanA-type vancomycin-resistant enterococci isolated from poultry meat and from fecal samples of humans in the community after discontinuation of avoparcin usage in animal husbandry. Microb. Drug Resist. 1999, 5, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, G.S.; Haaheim, H.; Dahl, K.H.; Kruse, H.; Løvseth, A.; Olsvik, Ø.; Sundsfjord, A. Transmission of vanA-type vancomycin-resistant enterococci and vanA resistance elements between chicken and humans at avoparcin-exposed farms. Microb. Drug Resist. 1998, 4, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Borgen, K.; Sorum, M.; Wasteson, Y.; Kruse, H. VanA-type vancomycin-resistant enterococci (VRE) remain prevalent in poultry carcasses 3 years after avoparcin was banned. Int. J. Food Microbiol. 2001, 64, 89–94. [Google Scholar] [CrossRef]
- Del Grosso, M.; Caprioli, A.; Chinzari, P.; Fontana, M.C.; Pezzotti, G.; Manfrin, A.; Di Giannatale, E.; Goffredo, E.; Pantosti, A. Detection and characterization of vancomycin-resistant enterococci in farm animals and raw meat products in Italy. Microb. Drug Resist. 2000, 6, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.K.; Kim, T.S.; Lee, H.S.; Nam, H.M.; Joo, Y.S.; Koh, H.B. Persistence of vanA-type Enterococcus faecium in Korean livestock after ban on avoparcin. Microb. Drug Resist. 2006, 12, 136–139. [Google Scholar] [CrossRef]
- Heuer, O.E.; Pedersen, K.; Jensen, L.B.; Madsen, M.; Olsen, J.E. Persistence of vancomycin-resistant enterococci (VRE) in broiler houses after the avoparcin ban. Microb. Drug Resist. 2002, 8, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leinweber, H.; Alotaibi, S.M.I.; Overballe-Petersen, S.; Hansen, F.; Hasman, H.; Bortolaia, V.; Hammerum, A.M.; Ingmer, H. Vancomycin resistance in Enterococcus faecium isolated from Danish chicken meat is located on a pVEF4-like plasmid persisting in poultry for 18 years. Int. J. Antimicrob. Agents 2018, 52, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Sting, R.; Richter, A.; Popp, C.; Hafez, H.M. Occurrence of vancomycin-resistant enterococci in Turkey flocks. Poult. Sci. 2013, 92, 346–351. [Google Scholar] [CrossRef]
- Top, J.; Willems, R.; Bonten, M. Emergence of CC17 Enterococcus faecium: From commensal to hospital-adapted pathogen. FEMS Immunol. Med. Microbiol. 2008, 52, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Savin, M.; Bierbaum, G.; Hammerl, J.A.; Heinemann, C.; Parcina, M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-resistant bacteria and antimicrobial residues in wastewater and process water from German pig slaughterhouses and their receiving municipal wastewater treatment plants. Sci. Total Environ. 2020, 727, 138788. [Google Scholar] [CrossRef]
- Correa-Martinez, C.L.; Tönnies, H.; Froböse, N.J.; Mellmann, A.; Kampmeier, S. Transmission of vancomycin-resistant enterococci in the hospital setting: Uncovering the patient– environment interplay. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 1–11. [Google Scholar] [CrossRef]
- Hasman, H.; Agersø, Y.; Hendriksen, R.; Cavaco, L.M.; Guerra-Roman, B. Isolation of ESBL-AmpC-and Carbapenemase-Producing E. coli from Fresh Meat; Laboratory Protocol. Version 4; DTU Foof: Lyngby, Denmark, 2018. [Google Scholar]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | No. of Samples | No. of Positive Samples in Direct PCR | No. of Isolates Cultured from Samples | ||||
|---|---|---|---|---|---|---|---|
| Carbapene-Mases (n) | mcr (n) | CPE | Col-E | VRE | ESBL | ||
| Carcasses broilers | 57 | blaOXA-48 (57) | mcr-1 (20), mcr-5 (22) ** | 0 | mcr-1 (6), mcr-5 (1) | 1 | 8 * |
| Carcasses pigs | 41 | blaOXA-48 (25) | mcr-2 (1), mcr-4 (2) | 0 | 0 | 0 | n.a. |
| Cecum broilers | 36 | – | mcr-1 (6), mcr-5 (1) | 0 | mcr-1 (5) | 0 | 20 |
| Cecum pigs | 41 | – | mcr-1 (3) | 0 | mcr-1 (1) | 0 | 17 |
| Drain swab | 88 | blaOXA-48 (60) | mcr-1 (1), mcr-4 (14) | 0 | mcr-1 (1) | 0 | 8 |
| Meat juice | 88 | blaOXA-48 (38) | mcr-1 (4), mcr-4 (5) | 0 | mcr-1(1) | 0 | 13 |
| Beef | 10 | blaOXA-48 (1) | - | 0 | 0 | 0 | 1 |
| Pork | 37 | blaOXA-48 (4) | mcr-1 (1), mcr-4 (1), mcr-5 (1) | 0 | 0 | 0 | 3 |
| Poultry | 53 | blaOXA-48 (4) | mcr-1 (13) | 0 | mcr-1 (6) | 0 | 15 |
| Water | 37 | blaOXA-48 (6) | mcr-4 (1) | 0 | 0 | 0 | 0 |
| Sewage/Sludge/Soil | 17 | blaOXA-48 (7), blaKPC (2), blaVIM/NDM (3) | mcr-1 (4) | blaOXA-48 (6), blaKPC (2) | 0 | 4 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klees, S.; Effelsberg, N.; Stührenberg, B.; Mellmann, A.; Schwarz, S.; Köck, R. Prevalence and Epidemiology of Multidrug-Resistant Pathogens in the Food Chain and the Urban Environment in Northwestern Germany. Antibiotics 2020, 9, 708. https://doi.org/10.3390/antibiotics9100708
Klees S, Effelsberg N, Stührenberg B, Mellmann A, Schwarz S, Köck R. Prevalence and Epidemiology of Multidrug-Resistant Pathogens in the Food Chain and the Urban Environment in Northwestern Germany. Antibiotics. 2020; 9(10):708. https://doi.org/10.3390/antibiotics9100708
Chicago/Turabian StyleKlees, Sylvia, Natalie Effelsberg, Birgit Stührenberg, Alexander Mellmann, Stefan Schwarz, and Robin Köck. 2020. "Prevalence and Epidemiology of Multidrug-Resistant Pathogens in the Food Chain and the Urban Environment in Northwestern Germany" Antibiotics 9, no. 10: 708. https://doi.org/10.3390/antibiotics9100708
APA StyleKlees, S., Effelsberg, N., Stührenberg, B., Mellmann, A., Schwarz, S., & Köck, R. (2020). Prevalence and Epidemiology of Multidrug-Resistant Pathogens in the Food Chain and the Urban Environment in Northwestern Germany. Antibiotics, 9(10), 708. https://doi.org/10.3390/antibiotics9100708