Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3


Abstract
:1. Introduction
2. Results
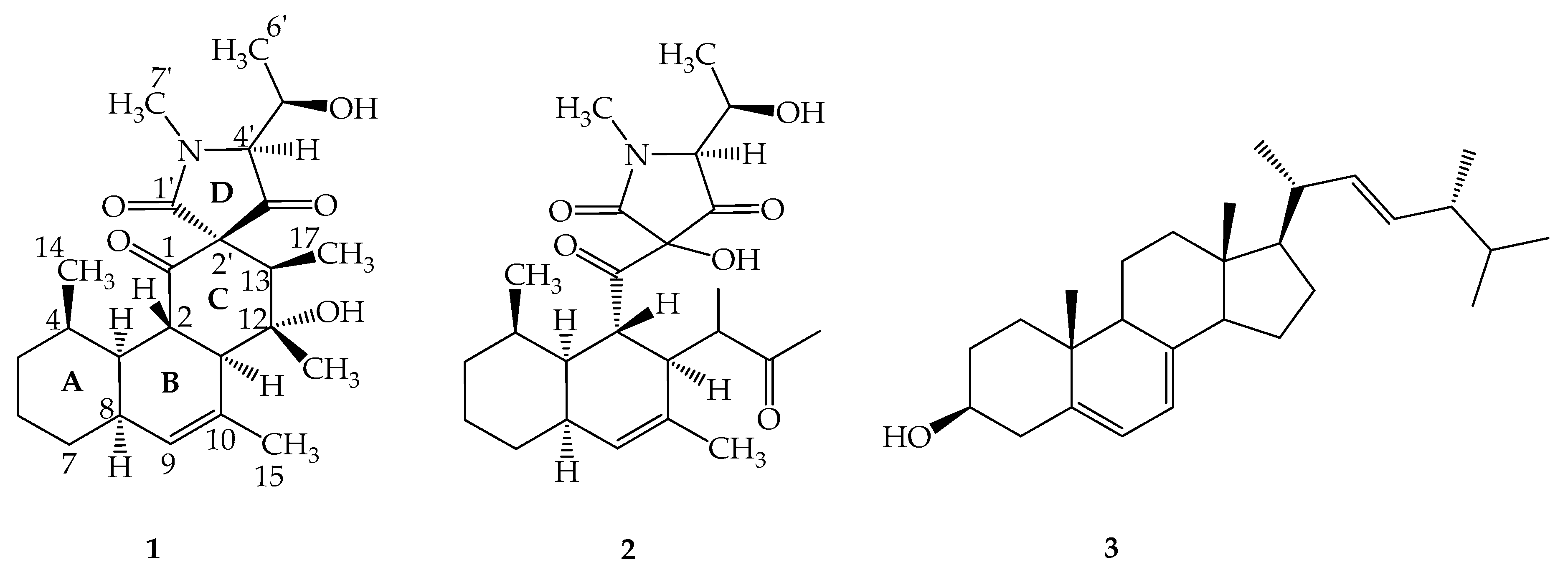
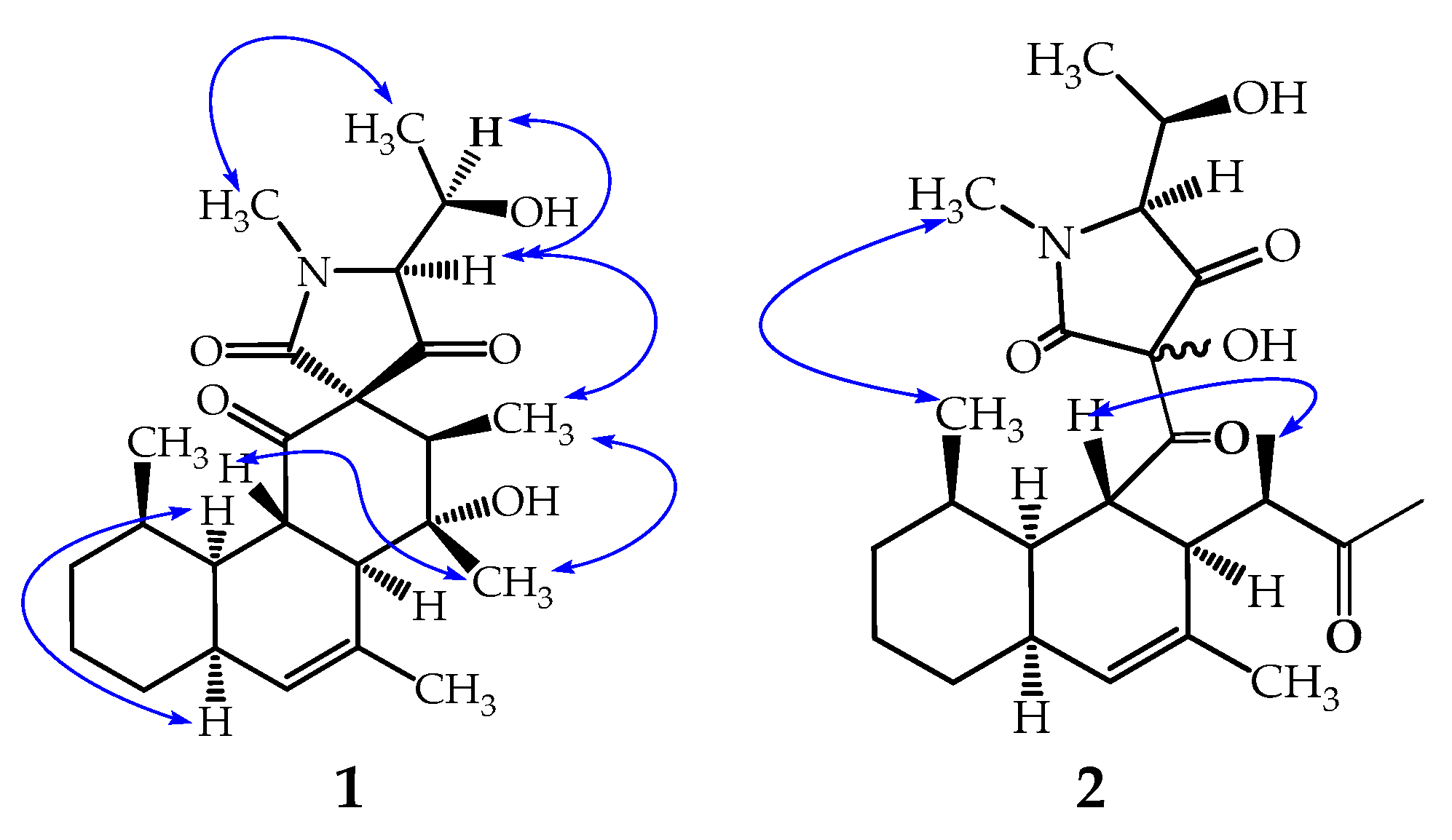
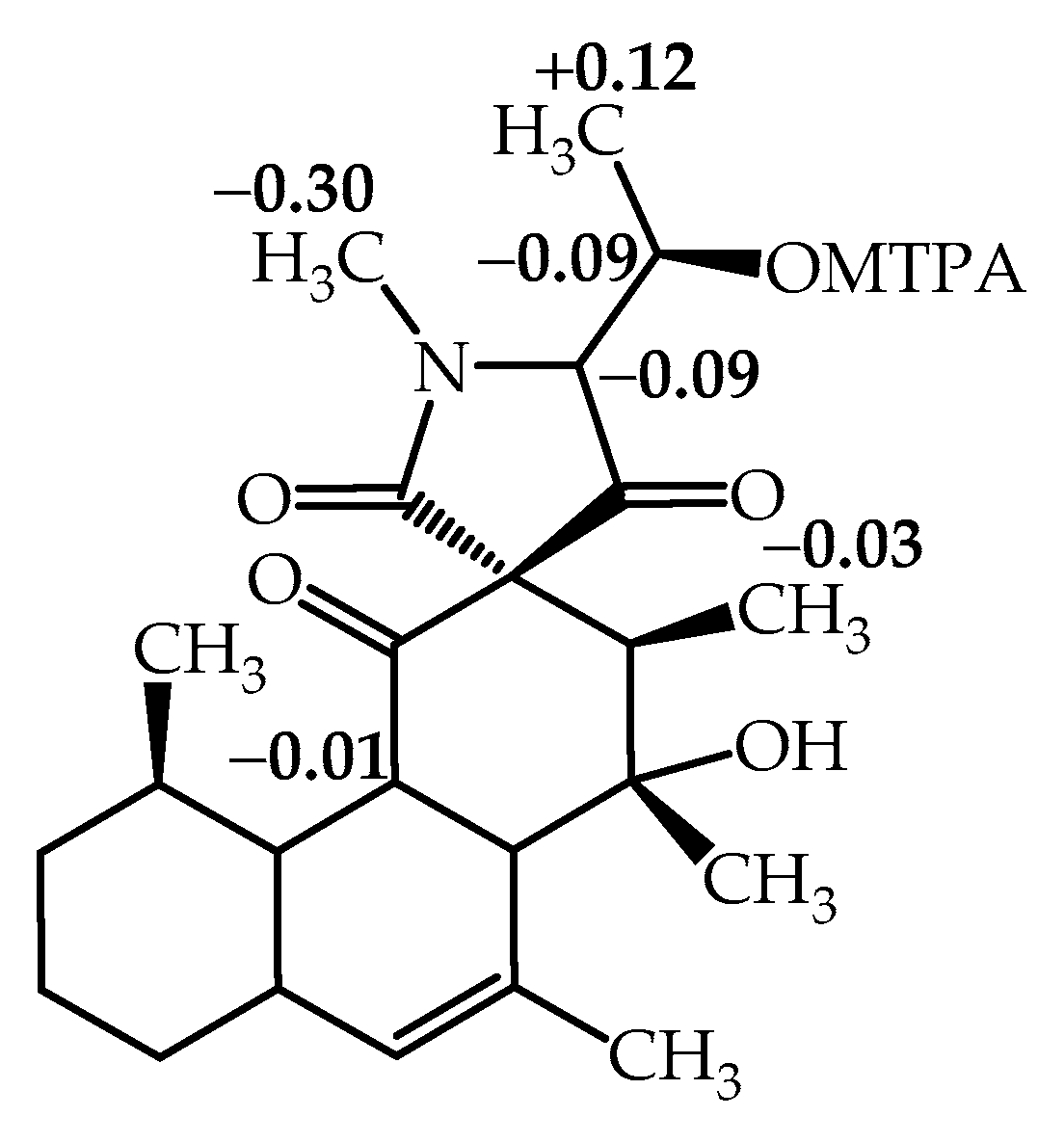
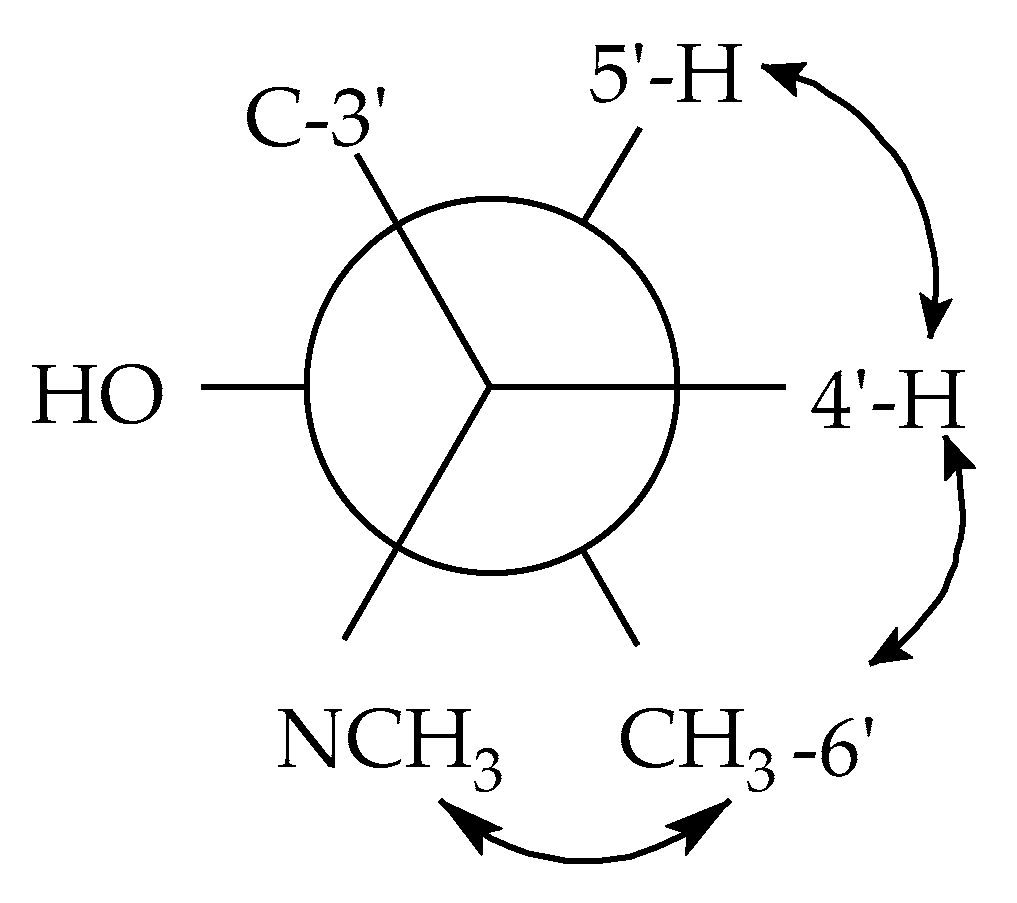
2.1. Structure Elucidation of Simplicilone A (1)
2.2. Structure Elucidation of Simplicilone B (2)
2.3. Biological Activity of Simplicilones A (1) and B (2)
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Isolation of Endophytic Fungus
4.3. Molecular Analysis, Sequencing and Phylogenetic Analysis of Endophytic Fungus
4.4. Fermentation, Extraction and Isolation
4.5. Bioassay for Cytotoxic Activity
4.6. Bioassay for Anti-Microbial Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef]
- Schulz, B.J.E.; Boyle, C.J.C.; Sieber, T.N. Microbial Root Endophytee; Springer: Berlin/Heidelberg, Germany, 2006; Volume 9, pp. 1–13. [Google Scholar]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites (1987 to 2000). Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Lòpez-Fernàndez, S.; Campisano, A.; Schulz, B.J.; Steinert, M.; Stadler, M.; Surup, F. Viriditins from Byssochlamys spectabilis, their stereochemistry and biosynthesis. Tetrahedron Lett. 2020, 61, 151446. [Google Scholar] [CrossRef]
- Halecker, S.; Surup, F.; Solheim, H.; Stadler, M. Albiducins A and B, salicylaldehyde antibiotics from the ash tree-associated saprotrophic fungus Hymenoscyphus albidus. J. Antibiot. 2017, 71, 339–341. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.-P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Happi, M.G.; Kouam, S.F.; Talontsi, F.M.; Nkenfou, C.; Longo, F.; Ngadjui, B.T.; Spiteller, M. New dimeric naphtopyrones with antimicrobial activity from Aspergillus niger, an Fungal Endophyte isolated from the Cameroonian plant Entandrophragma sp. Zeit Naturforschung 2013, 70, 625–630. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Lamshöft, M.; Douanla-Meli, C.; Kouam, S.F.; Spiteller, M. Antiplasmodial and cytotoxic dibenzofurans from Preussia sp. harboured in Enantia chlorantha Oliv. Fitoterapia 2014, 93, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Ngadjui, B.T.; Lontsi, D.; Ayafor, J.F.; Sondengam, B. Pachypophyllin and pachypostaudins A and B: Three bisnorlignans from pachypodanthium staudtii. Phytochemistry 1989, 28, 231–234. [Google Scholar] [CrossRef]
- Dieter, N. African Ethnobotany: Poisons and Drugs: Chemistry Pharmacology and Toxicology; CRC Press: Boca Raton, FL, USA, 1996; pp. 57–59. [Google Scholar]
- Bouquet, A. Féticheurs et Médecines Traditionnelles du Congo-Brazzaville; Mémoire O.R.S.T.O.M.: Paris, France, 1969; Volume 36. [Google Scholar]
- Wickens, G.E.; Burkill, H.M. The useful plants of West Tropical Africa. Kew Bull. 1986, 41, 471. [Google Scholar] [CrossRef]
- Ngouonpe, A.W.; Mbobda, A.S.W.; Happi, G.M.; Mbiantcha, M.; Tatuedom, O.K.; Ali, M.S.; Lateef, M.; Tchouankeu, J.C.; Kouam, S.F. Natural products from the medicinal plant Duguetia staudtii (Annonaceae). Biochem. Syst. Ecol. 2019, 83, 22–25. [Google Scholar] [CrossRef]
- Wei, D.-P.; Wanasinghe, D.N.; Hyde, K.D.; Mortimer, P.E.; Xu, J.; Xiao, Y.-P.; Bhunjun, C.S.; To-Anun, C. The genus Simplicillium. MycoKeys 2019, 60, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Someno, T.; Sawa, R.; Iinuna, H.; Naganawa, H.; Ishizuka, M.; Takeuchi, T. Delaminomycins, novel nonpeptide extracellular matrix receptor antagonist and a new class of potent immunomodulator. J. Antiot. 1993, 46, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Koyama, N.; Nagahiro, T.; Yamaguchi, Y.; Ohshiro, T.; Masuma, R.; Tomoda, H.; Omura, S. Spylidone, a novel inhibitor of lipid droplet accumulation in mouse macrophages produced by Phoma sp. FKI-1840. J. Antibiot. 2005, 58, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoye, T.R.; Jeffrey, C.S.; Shao, F. Mosher ester analysis for the determination of absolute configuration of stereogenic (chiral) carbinol carbons. Nat. Protoc. 2007, 2, 2451–2458. [Google Scholar] [CrossRef] [PubMed]
- Minowa, N.; Kodama, Y.; Harimaya, K.; Mikawa, T. A degradation study of vermisporin and determination of its absolute configuration. Heterocycles 1998, 48, 1639. [Google Scholar] [CrossRef]
- Takata, K.; Iwatsuki, M.; Yamamoto, T.; Shirahata, T.; Nonaka, K.; Masuma, R.; Hayakawa, Y.; Hanaki, H.; Kobayashi, Y.; Petersson, G.A.; et al. Aogacillins A and B Produced by Simplicillium sp. FKI-5985: New Circumventors of Arbekacin Resistance in MRSA. Org. Lett. 2013, 15, 4678–4681. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Dutta, T.; Sarkar, T.S.; Ghosh, S. Novel xylanases from Simplicillium obclavatum MTCC 9604: Comparative analysis of production, purification and characterization of enzyme from submerged and solid state fermentation. SpringerPlus 2013, 2, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Sudoh, Y.; Tsuchiya, Y.; Okuda, T.; Igarashi, Y. Isolation and biosynthesis of preussin B, a pyrrolidine alkaloid from Simplicillium lanosoniveum. J. Nat. Prod. 2014, 77, 813–817. [Google Scholar] [CrossRef]
- Dai, Y.; Lin, Y.; Pang, X.; Luo, X.-W.; Salendra, L.; Wang, J.; Zhou, X.; Lu, Y.; Yang, B.; Liu, Y. Peptides from the soft coral-associated fungus Simplicillium sp. SCSIO41209. Phytochemistry 2018, 154, 56–62. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, X.; Nong, X.; Wang, J.; Huang, Z.-H.; Qi, S.-H. Eight linear peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. Tetrahedron 2016, 72, 3092–3097. [Google Scholar] [CrossRef]
- Liang, X.; Nong, X.-H.; Huang, Z.-H.; Qi, S.-H. Antifungal and antiviral cyclic peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. J. Agric. Food Chem. 2017, 65, 5114–5121. [Google Scholar] [CrossRef] [PubMed]
- Uchida, R.; Kondo, A.; Yagi, A.; Nonaka, K.; Masuma, R.; Kobayashi, K.; Tomoda, H. Simpotentin, a new potentiator of amphotericin B activity against Candida albicans, produced by Simplicillium minatense FKI-4981. J. Antibiot. 2018, 72, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Petrini, O. Endophytic fungi in British Ericaceae: A preliminary study. Trans. Br. Mycol. Soc. 1984, 83, 510–512. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Surup, F.; Halecker, S.; Nimtz, M.; Rodrigo, S.; Schulz, B.; Steinert, M.; Stadler, M. Hyfraxins A and B, cytotoxic ergostane-type steroid and lanostane triterpenoid glycosides from the invasive ash dieback ascomycete Hymenoscyphus fraxineus. Steroids 2018, 135, 92–97. [Google Scholar] [CrossRef]
- Becker, K.; Pfütze, S.; Kuhnert, E.; Cox, R.; Stadler, M.; Surup, F. Hybridorubrins A−D, novel azaphilone heterodimers from stromata of Hypoxylon fragiforme and insights into the biosynthetic machinery for azaphilone diversification. Chem. A Eur. J. 2020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos | 1 | 2 | ||
|---|---|---|---|---|
| δC | δH (mult., J in Hz) | δC | δH (mult., J in Hz) | |
| 1 | 201.7 | - | 209.3 | - |
| 2 | 43.2 | 3.72 (pseudo t, 11.6) | 38.8 | 4.44 (dd, 11.6, 8.8) |
| 3 | 37.9 | 2.11 (dt, 3.6, 11.6) | 44.7 | 2.29 (m) |
| 4 | 39.5 | 1.65 (m) | 38.2 | 1.70 (m) |
| 5 | 30.9 | 1.29 (m) | 29.0 | 1.25 (m), 1.20 (m) |
| 6 | 27.8 | 1.73 (m), 1.30 (m) | 28.6 | 1.76 (m), 1.26 (m) |
| 7 | 31.5 | 1.66 (m), 1.21 (m) | 31.4 | 1.28 (m) |
| 8 | 40.3 | 1.99 (m) | 39.9 | 1.98 (m) |
| 9 | 134.0 | 5.72 (br d, 6.7) | 131.5 | 5.73 (br d, 6.7) |
| 10 | 132.9 | - | 131.9 | - |
| 11 | 59.2 | 2.30 (d, 11.6) | 46.4 | 3.22 (dd, 2.5, 11.6) |
| 12 | 77.3 | - | 47.4 | 2.67 (dq, 2.5, 7.0) |
| 13 | 53.7 | 2.51 (q, 7.0) | 211.5 | - |
| 14 | 23.6 | 0.72 (d, 7.3) | 21.4 | 0.81 (d, 7.6) |
| 15 | 25.5 | 1.84 (s) | 21.6 | 1.65 (s) |
| 16 | 17.7 | 1.37 (s) | 10.1 | 1.18 (d, 7.3) |
| 17 | 11.4 | 0.88 (d, 7.0) | 28.4 | 2.07 (s) |
| 1’ | 172.8 | - | 163.8 | - |
| 2’ | 75.5 | - | 104.9 | - |
| 3’ | 205.8 | - | 191.4 | - |
| 4’ | 74.3 | 3.78 (d, 4.3) | 72.8 | 3.18 (d, 7.6) |
| 5’ | 69.8 | 4.04 (dq, 4.3, 7.0) | 65.9 | 4.13 (dq, 6.4, 7.6) |
| 6’ | 17.6 | 1.34 (d, 7.0) | 18.5 | 1.41 (d, 6.4) |
| 7’ | 31.7 | 3.19 (s) | 45.0 | 2.97 (s) |
| Cell Line | 1 | 2 | Reference (IC50) |
|---|---|---|---|
| L929 (IC50) | - | - | Epothilone B (0.00062) |
| KB3.1 (IC50) | 25 | 29 | Epothilone B (0.00003) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anoumedem, E.G.M.; Mountessou, B.Y.G.; Kouam, S.F.; Narmani, A.; Surup, F. Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3. Antibiotics 2020, 9, 753. https://doi.org/10.3390/antibiotics9110753
Anoumedem EGM, Mountessou BYG, Kouam SF, Narmani A, Surup F. Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3. Antibiotics. 2020; 9(11):753. https://doi.org/10.3390/antibiotics9110753
Chicago/Turabian StyleAnoumedem, Elodie Gisèle M., Bel Youssouf G. Mountessou, Simeon F. Kouam, Abolfazl Narmani, and Frank Surup. 2020. "Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3" Antibiotics 9, no. 11: 753. https://doi.org/10.3390/antibiotics9110753
APA StyleAnoumedem, E. G. M., Mountessou, B. Y. G., Kouam, S. F., Narmani, A., & Surup, F. (2020). Simplicilones A and B Isolated from the Endophytic Fungus Simplicillium subtropicum SPC3. Antibiotics, 9(11), 753. https://doi.org/10.3390/antibiotics9110753