1. Introduction
Fungal infections are widely recognized as one of the main causes of morbidity and mortality, particularly those infections caused by opportunistic pathogenic fungi, such as the
Candida species [
1]. The incidence of
Candida infections (Candidosis) has increased remarkably in the last years [
1].
On human pathology,
Candida species have an important role as colonizers of the mucosal membranes of the oral cavity and gastrointestinal tract, as well as normal components of the skin and vaginal flora; so, in normal conditions,
Candida are non-pathogenic commensal microorganisms in humans [
2,
3]. However, modifications in human host defenses may lead to a disproportional growth of
Candida and consequently to a pathogenic colonization by these species [
4,
5]. In general, transition from commensal to pathogen in
Candida is facilitated by a series of virulence factors, such as hemolytic activity, secretion of extracellular hydrolytic enzymes (coagulase, phospholipase, and proteases), and producing specific adhesins (for example, fibrinogen and fibronectin), which appear to play an important role in adhesion, penetration, invasion, and destruction of human tissues. Also, the ability of
Candida to adhere to medical devices or host tissues with the formation of more resistant structures (biofilms) is a particularly important virulence factor [
6,
7]. In normal environments, biofilm formation represents the most predominant type of microbial growth and is frequently associated to persistent clinical infections [
8,
9]. While each
Candida species possesses unique features, biofilms in nature are formed by more than one microbial species, both bacteria and fungi, which confers an even higher resistant phenotype to the biofilm [
10]. Although the study of the structure and properties of single-species biofilms is an important step for understanding infectious diseases, the elucidation of communal behavior of microorganisms in biofilms composed of different species may have a high impact for understanding infectious diseases and to develop new therapeutic strategies [
10]. Indeed,
Candida species and
Pseudomonas aeruginosa microorganisms tend to form polymicrobial biofilms and, as such, they are often responsible for nosocomial infections in immunocompromised individuals [
11,
12].
Candida albicans remains as the most prevalent species of these infections, but a clear rise in the proportion of non-
Candida albicans Candida (NCAC) species has been noted [
6,
7,
13]. According to some epidemiological studies,
Candida tropicalis stands out in the NCAC species group and is correlated with other forms of
Candida mortality [
14,
15,
16]. For example, a predominance of NCACs species was observed in the south of America, where
C. albicans accounted for 40.9% of cases, followed by
C. tropicalis (20.9%),
C. parapsilosis (20.5%), and
C. glabrata (4.9%) [
7,
17,
18]. A study by Kontoyiannis et al. has shown that
C. tropicalis is more persistent, leading this process to situations of uselessness in cases of infection [
14]. This may imply increased virulence and resistance to antifungals compared to
C. albicans, for example, particularly in the oral cavity [
19]. As a consequence of the lack of knowledge on NCAC species virulence, namely in
C. tropicalis, the rising levels of resistance to the traditional antifungal therapies, and the association of Candidosis to high levels of mortality, there is an urgent need to develop new strategies to fight these infections [
1]. In this sense, an alternative approach for the treatment of Candidosis is the use of natural compounds as antifungal agents, among which is honey. Honey is a natural product which has been known for its biological and pharmacological properties for centuries. It has been extensively used in traditional medicine and also complementary medicine because of its antibacterial, antifungal, antimycobacterial, and antiviral activities [
20]. These antimicrobial activities may be caused by honey’s acidity (low pH), osmotic effect, high sugar concentration, presence of bacteriostatic and bactericidal factors, increase in cytokine release, as well as immune modulating and anti-inflammatory properties [
21,
22]. Besides that, one of the most important factors is the presence of hydrogen peroxide (H
2O
2), the primary antimicrobial component in most honeys, produced by the enzyme glucose oxidase, and its action consists in the destruction of the essential components of the cells [
21,
23]. Some types of honey contain additional antimicrobial activity by methylglyoxal (MGO), bee defensin-1, and other bee-derived compounds, such as phenolic compounds of floral origin and lysozyme, among other compounds [
23].
The present work aims to evaluate the antimicrobial effect of two different honeys (Portuguese Heather (PH) and manuka) alone, or in combination with a commercial antimicrobial agent, in mixed biofilms of C. tropicalis with P. aeruginosa.
3. Discussion
Analyzing the MIC and MFC values of the two honeys under study for
C. tropicalis, it was verified that this strain is susceptible to both honeys, especially the PH. Similar results were obtained by Khosravi et al. for
C. tropicalis (MIC of 38.5% and MFC of 43.7%) [
24].
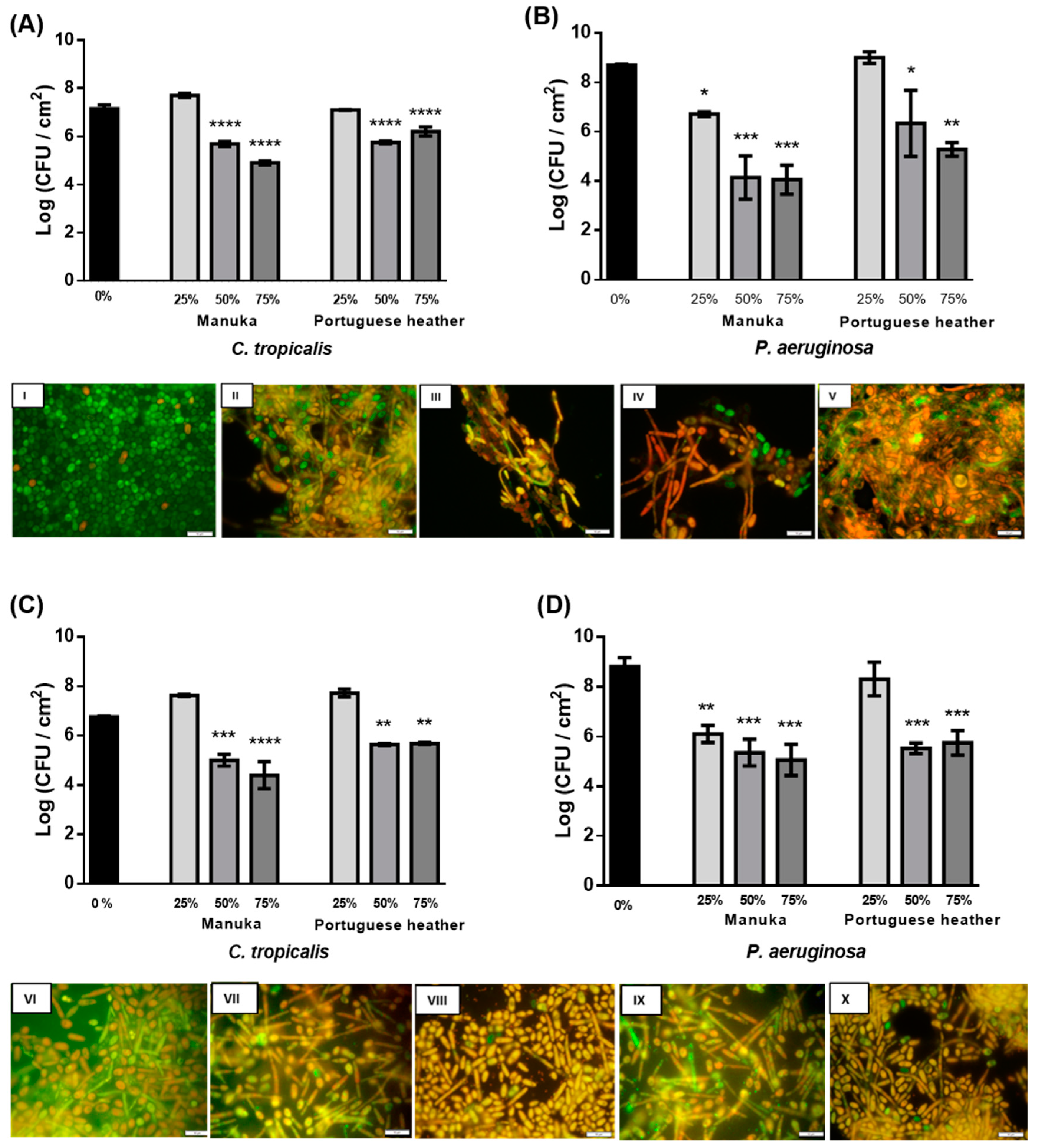
The antimicrobial effect of PH and manuka honeys in 24 h biofilms of
C. tropicalis and
P. aeruginosa was determined through quantification of viable cells (CFU) per unit of area (cm
2) (
Figure 1). Regarding
C. tropicalis single biofilms, after 24 h of treatment, none of the honeys at 25% (w/v) caused cell reduction (
Figure 1A). This may be a consequence of the fungal cells taking advantage of the lower sugar concentration present in honey at 25% to promote their own growth. Still, significant reductions were obtained with both honeys at 50% (w/v) (about 1.5 Log-reduction (CFU/cm
2)) and with manuka honey at 75% (w/v) (
p < 0.0001) (about 2 Log (CFU/cm
2) (
Figure 1A)). A significant reduction was only observed with 75% (w/v) PH (
p < 0.01). Some properties of honey may be involved in the antimicrobial effect, particularly in the antifungal effect [
21,
22], such as the presence of metilglioxal (MGO) [
25]. an effective antimicrobial agent against planktonic and biofilm cells [
26]. The presence of aromatic acids or special proteins with antifungal activity, such as flavonoids, polyphenois, and defensin-1 [
25], and the production of H
2O
2 by the enzyme glucose oxidase [
26]. Indeed, in a previous study, the authors have explored the analysis of these different characteristics of honeys, and the results showed that PH honey was one of the most interesting values indicating the possible high antimicrobial potential (data not shown).
In
C. tropicalis single and mixed biofilms, after 24 h of treatment, 75% (w/v) manuka honey significantly reduced viable cells in 2 Log (CFU/cm
2) (
p < 0.0001) (
Figure 1A,C). For PH, similar results were obtained with 50% (w/v) and 75% (w/v) honey, with a significant reduction of 1 Log (CFU/cm
2) (
p < 0.0001) (
Figure 1A–C). These data suggest that, after 24 h of treatment, manuka honey exerts an antifungal effect both in the presence and absence of
P. aeruginosa, while PH exerts a slightly higher effect in single species biofilms of
C. tropicalis.
The images obtained by fluorescence microscopy (
Figure 1I–X), which are only representative of the cell form and structure, revealed a direct relationship between the presence of honey or
P. aeruginosa and the hyphae. In
Candida species, the morphological transition involving the formation of hyphae is an important virulence factor, which is associated with cell stress, which could be caused by the presence of
P. aeruginosa or the several compounds of honey (such as H
2O
2 and defensin-1) [
27]. Also, from the microscopy images (
Figure 1I–X), it was observed that, curiously, the untreated mixed biofilm presented several damaged cells of
C. tropicalis (
Figure 1VI), compared to single biofilms (
Figure 1I). This is supported by the experiments carried out by Bandara et al. that investigated the interactions between
Candida species and
P. aeruginosa, showing a reduction of 88% (w/v) of
C. tropicalis after a 24 h-incubation [
10]. It was confirmed in the same study that, in general,
Candida species and
P. aeruginosa have mutually suppressive effects at all stages of biofilm formation. However, most of the previous studies on interactions between
Candida species and bacteria in mixed biofilms are concentrated in
C. albicans and only a few are related to NCAC [
10].
Regarding the results obtained for
P. aeruginosa in single and mixed biofilms, significant reductions could be observed after 6 h of treatment for both honeys at all concentrations tested. In this period of time, the manuka honey allowed for a reduction of the single biofilm to half of the viable cells (
p < 0.0001) and the PH managed to reduce in average 2–3 Log (CFU/cm
2). In mixed biofilms, there was also a significant reduction, but it was not as pronounced as in single biofilms (
p < 0.001). With the results obtained, it can be observed that the honey has a faster and superior antibacterial effect compared to the results obtained for
C. tropicalis. Comparing the two honeys tested, it was observed that the highest reductions were obtained with manuka honey: 4 and 3 Log (CFU/cm
2) with 50% (w/v) and 75% (w/v) (
p < 0.001) in single (
Figure 1B) and mixed (
Figure 1D) biofilms, respectively. However, the reduction occurred from 25% (w/v) of manuka honey, both in single (
p < 0.1) (
Figure 1B) and in mixed biofilms (
p < 0.01) (
Figure 1D). The effect of lower concentrations of manuka on
P. aeruginosa was already reported by Cooper and Molan [
28]. According to the literature, honey has broad-spectrum antibacterial activity. Regarding the effect of manuka honey on bacteria, a study by Roberts, Moddocks, and Cooper found that honey acts by inhibiting the flogging of bacteria, which limits their mobility and prevents the formation of biofilms [
29]. In summary, after 24 h of treatment, honey promoted cell reduction in biofilms of both species simultaneously, suggesting honey as a promising agent for the treatment of polymicrobial infections of
C. tropicalis and
P. aeruginosa. Very few treatments were reported as being able to reduce more than one species in a mixed biofilm.
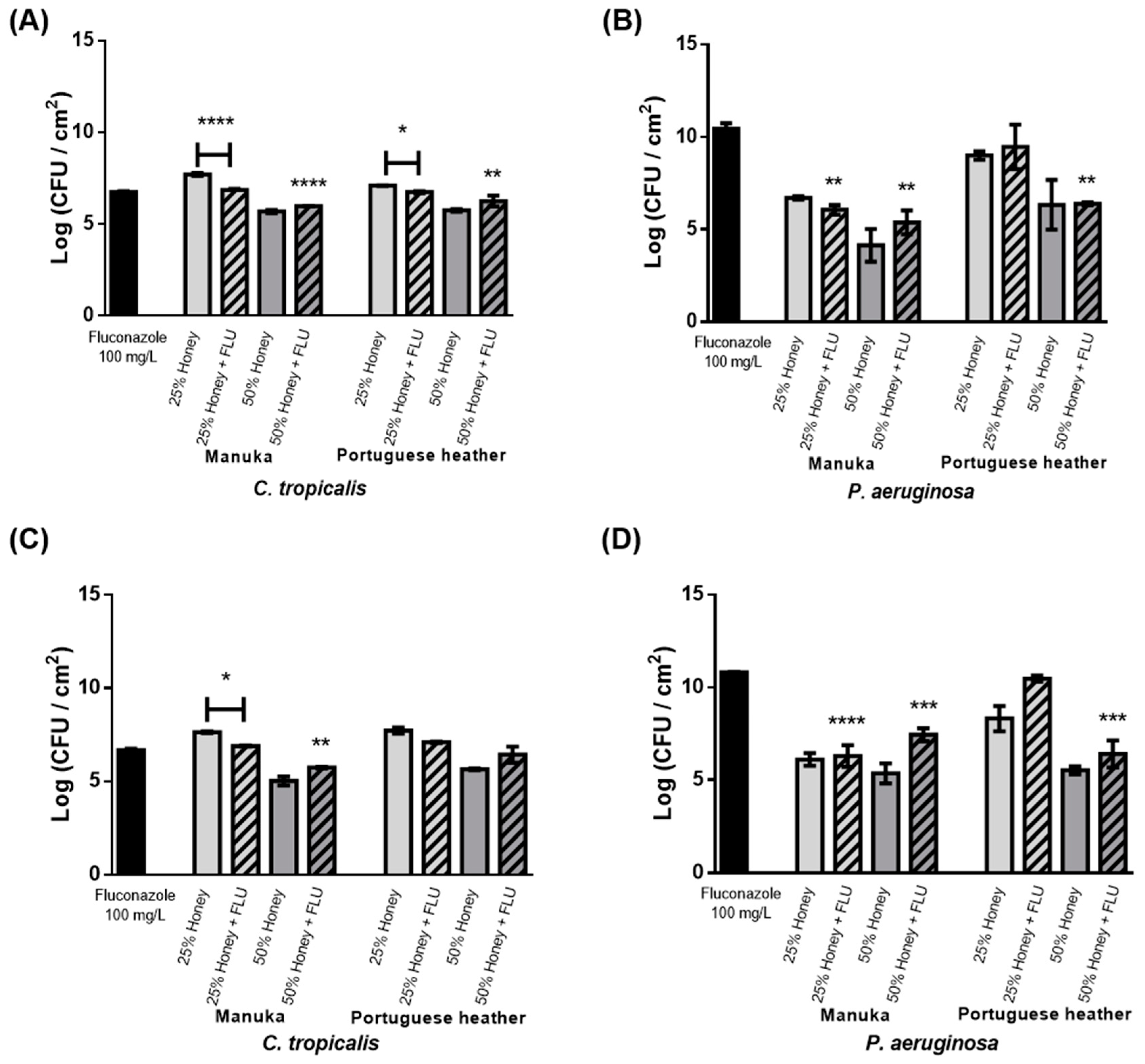
The effect of the combination of honey (PH or manuka honeys at 25% (w/v) and 50% (w/v)) with FLU (100 mg L
−1) was then evaluated in single and mixed biofilms (
Figure 2). For
C. tropicalis, the cell reduction obtained with the combination (honey and FLU) was higher than with only honey at 25% (w/v) (1 Log reduction for manuka honey,
p < 0.0001; 0.5 Log for PH,
p < 0.1) (
Figure 2A). Conversely, combinations of honey at 50% (w/v) and FLU had no advantage over honey treatment alone; nevertheless, an improved reduction was observed when compared to the effect of FLU alone (
p < 0.0001 for manuka honey and
p < 0.01 for heather honey). Comparing results obtained for honey combined with an antifungal agent, it was observed that it had advantages over single honey or antifungal treatment. The combined treatments allowed for a 50% reduction of the dosage of antifungal typically required in clinical settings, i.e., 200 mg L
−1.
In mixed biofilms, a slight reduction in biofilm was observed after 12 h compared to the individual treatment with either honey or FLU. After 24 h of combinational therapy, a significant reduction was only obtained in comparison to treatment with manuka honey alone at 25% (w/v) (
p < 0.1) (
Figure 2C), similar to what was observed for single biofilms (
Figure 2A). This demonstrates that the combination of
C. tropicalis and
P. aeruginosa is indifferent to treatment with honey and FLU, and that
C. tropicalis remains less tolerant to FLU. Furthermore, adding FLU to honey substantially increased the number of
P. aeruginosa viable cells in mixed biofilms (
Figure 2D) compared with treatment with only honey.
In summary, honey by itself allows for a superior reduction in comparison with the combination of honey and FLU in all conditions tested in mixed and single biofilms, with the exception of 25% (w/v) honey.
C. tropicalis remains highly resistant to FLU, even when combined with honey, both in single and mixed biofilms. This is in accordance with several studies stating that resistance to FLU is increasing in clinical isolates of this species [
29].
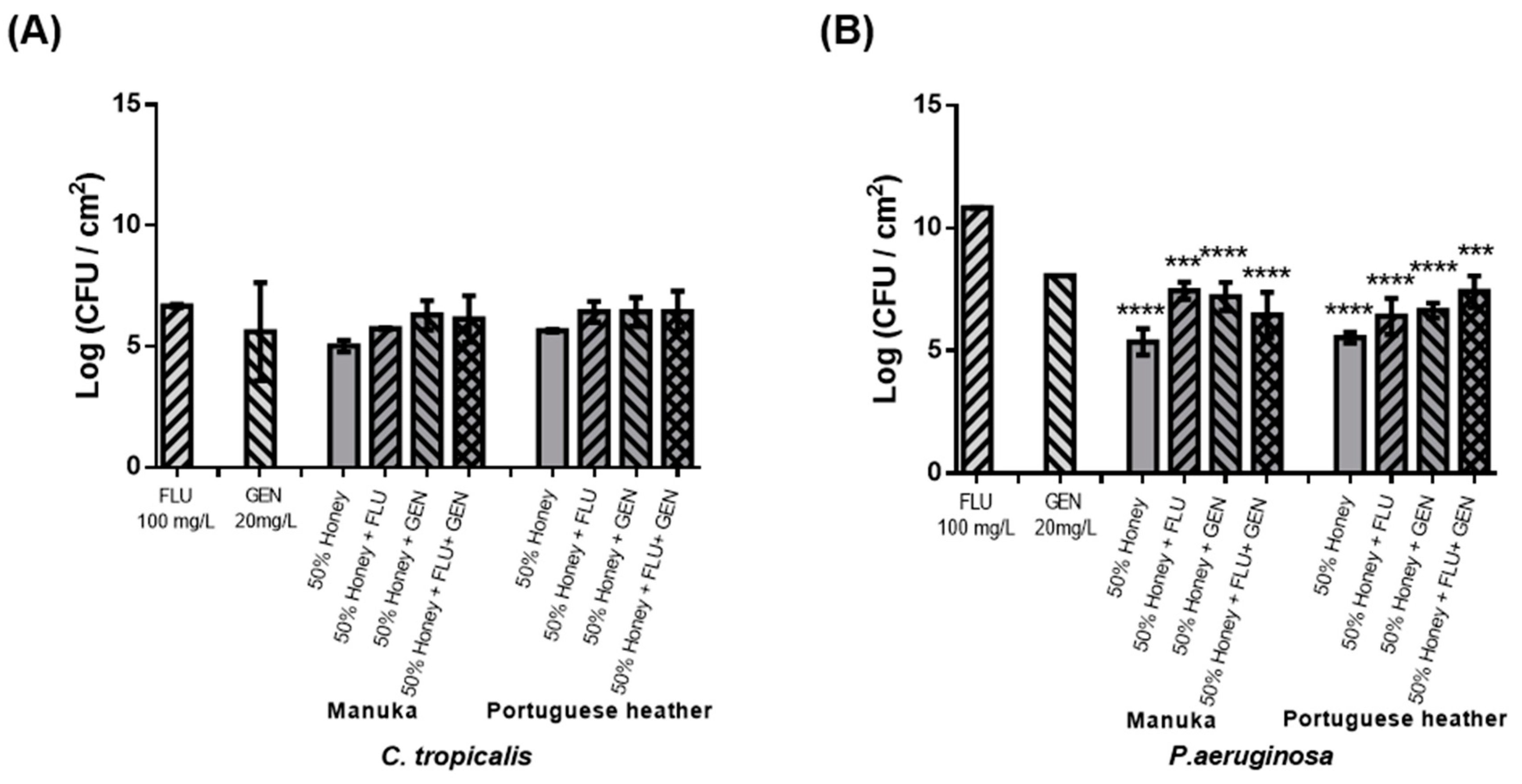
Considering the results previously obtained, a third antimicrobial agent was added (GEN) to the combinations tested. So, combinational therapy was evaluated using honey 50% (w/v), FLU 100 mg L
−1, and GEN 20 mg L
−1. However, this resulted in an increased number of viable cells in biofilms compared to treatment with honey alone, for all conditions tested (
Figure 3). Indeed, the combination of these three elements had no advantage over treatment with honey alone, both for the fungal and bacterial species. The interaction between FLU and GEN was investigated by Thomas et al. as a treatment of pre-formed biofilms of 4, 8, and 12 h of
C. albicans, where synergism was observed only against pre-formed biofilms of 4 and 8 h, with no synergism observed at 24 h [
30].
4. Material and Methods
4.1. Microorganisms and Culture Conditions
C. tropicalis ATCC 750 and P. aeruginosa DSM 22644 were stored at –80 ± 2 °C in broth medium with 20% (v/v) glycerol. Prior to each assay, C. tropicalis and P. aeruginosa strains were subcultured from the frozen stock preparations onto Sabouraud Dextrose Agar (SDA) and Tryptic Soy Agar (TSA) plates, respectively. SDA and TSA were prepared from Sabouraud Dextrose Broth (SDB; Liofilchem, Roseto degli Abruzzi, Italy) or Tryptic Soy Broth (TSB; Liofilchem, Roseto degli Abruzz, Italy), supplemented with 2% (w/v) agar (Liofilchem, Roseto degli Abruzzi, Italy). The plates were then incubated aerobically at 37 °C for 18–24 h.
Pure liquid cultures (pre-inocula) of C. tropicalis were maintained in SDB, whereas P. aeruginosa was grown overnight in TSB. For planktonic and biofilm assays, 0.22 µm filter-sterilized Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco® by Life Technologies TM, Grand Island, NY, USA) at pH 7.0 was used. Unless otherwise stated, all rinse steps were performed either by using 0.9% (w/v) saline solution (NaCl; J.T. Baker, Deventer, The Netherlands) or ultrapure (UP) sterile water.
4.2. Antimicrobial Agents
Stock solutions of two commercial antimicrobial agents, Fluconazole (FLU, Sigma-Aldrich, St. Louis, MO, USA) and Gentamicin sulfate (GEN, Sigma-Aldrich, St. Louis, MO, USA) were prepared and stored according to the manufacturer’s instructions. Also, two different honeys, Portuguese heather honey (raw dark amber honey whose main plant nectar is heather (30%, APISMaia company (Porto Portugal)) collected by a beekeeper in the North of Portugal) and manuka (commercial Medihoney
®) were stored at 4 °C, and the dilutions were prepared with RPMI 6420 medium. All the concentrations of different antimicrobial agents (natural and commercial) tested in this work are presented in
Table 1.
4.3. Planktonic Antimicrobial Susceptibilities
Susceptibilities of
C. tropicalis planktonic-cell cultures were evaluated by determining the minimum inhibitory concentration (MIC) and the minimum fungicidal concentration (MFC). The MIC values were determined according to standard European Committee on Antimicrobial Susceptibility Testing (EUCAST), through the broth microdilution method [
32]. Briefly, the initial cell concentration for both microorganisms was adjusted for 1 × 10
6 CFU/mL and dispensed into 96-well plates in a proportion of 1:2 (the final inoculum concentration was 5 × 10
5 CFU/mL) with the working antimicrobial solutions (previously diluted in RPMI 1640 broth with double of the desired final concentration). Wells containing only broth medium (antimicrobial-free medium) were used as negative controls and wells containing
C. tropicalis culture without antimicrobial agent were used as positive controls. Plates were incubated overnight at 37 °C. MIC was obtained by visual observation of the turbidity gradient. The minimum concentration where growth inhibition occurs is equivalent to the MIC value.
For the determination of MFC values, 10 μL was removed from each well of the microdilution trays, after incubation, and plated onto SDA plates and incubated at 37 °C. The lowest antimicrobial concentration that yielded no colony growth after 12–24 h was considered as the MFC.
4.4. Biofilms Antimicrobial Susceptibilities
Biofilms were developed according to the modified microtiter plate test proposed by Stepanović et al., with some modifications [
33]. Briefly, different cultures were centrifuged twice (3000
g, 4 °C, 10 min) and the pellet was resuspended in RPMI 1640, until reaching 1 × 10
7 cells/mL. Yeast cells (
C. tropicalis) were enumerated by microscopy using a Neubauer counting chamber. Bacteria (
P. aeruginosa) concentration was adjusted to 0.13 (corresponding between 2–3 × 10
−8 CFU·mL
−1), using an ELISA microtiter plate reader with a wavelength of 640 nm (Sunrise-Basic Tecan, Männedorf, Switzerland). For mixed-species cultures, a combination of 50% of the suspended inoculum of each species was used.
The cellular suspensions were transferred, under aseptic conditions, to 96-well flat tissue culture plates (polystyrene, Orange Scientific, Braine-L’Alleud, Belgium) (200 μL per well). To promote biofilm formation, microtiter plates were incubated aerobically for 24 h on a horizontal shaker at 120 rpm and 37 °C.
The effect of each honey (PH or manuka) alone was evaluated in single and mixed-species biofilms of C. tropicalis and P. aeruginosa. For this, 24 h-old biofilms were exposed to increasing concentrations of each agent (25% (w/v), 50% (w/v), and 75% (w/v)). Briefly, after biofilm formation, 200 μl of cell suspension were replaced by the antimicrobial solutions prepared at 2-fold the desired concentration. Plates were then incubated aerobically at 37 °C for 24 h. After 6, 12, and 24 h, the treated biofilms were removed to assess biofilm-cells cultivability through CFU enumeration.
After biofilm formation, the wells were washed twice with saline solution after discarding the planktonic fraction. In order to estimate the number of cultivable biofilm-entrapped cells in single- and mixed-species, the microdrop technique was used. Briefly, 200 μL of fresh saline solution was added to each well and the biofilms were scraped. The resulting biofilm-cells suspensions were then serially diluted in saline solution and plated onto non-selective agar (SDA for C. tropicalis and TSA for P. aeruginosa pure cultures) plates. Selective agar was also used for colony-forming units (CFU) determination of Candida species (SDA supplemented with 20 mg L−1 gentamycin (GEN), to suppress the growth of P. aeruginosa) and P. aeruginosa (Pseudomonas Isolation Agar, (PIA)). Agar plates were incubated aerobically at 37 °C for 24 h for cultivable cell counting. Values of cultivable sessile cells were expressed as Log CFU per area (cm2). All negative (wells containing only broth medium) and positive (wells containing C. tropicalis, P. aeruginosa, C. tropicalis, and P. aeruginosa cultures without antimicrobial agents) controls were performed.
4.5. Combinatorial Effect of Antimicrobial Agents (Honey and Commercial Agents) on Biofilms
The combinatorial effect of honeys (25% (w/v) and 50% (w/v) of PH or manuka honey with 100 mg L−1 FLU was assessed against 24 h-old C. tropicalis biofilms, following a procedure similar to the individual application of the antimicrobials, and against 24 h-old dual-species (C. tropicalis and P. aeruginosa) biofilms. Also, the triple combinatorial effect of antimicrobial agent (100 mg L−1 FLU and 20 mg L−1 GEN) and 50% (w/v) of honey (PH or manuka) was assessed against 24 h-old dual-species (C. tropicalis and P. aeruginosa) biofilms. Biofilm cells were removed after 6, 12, and 24 h to assess biofilm-cells cultivability through CFU enumeration. Also, all negative (wells containing only broth medium) and positive (wells containing C. tropicalis, P. aeruginosa, C. tropicalis, and P. aeruginosa cultures without antimicrobial agents) controls were performed.
4.6. Cell Viability Assessment of Biofilm-Embedded Cells
In order to evaluate the cell morphology and viability of polymicrobial biofilms after treatment in a qualitative way, the Live/Dead® BacLight™ Bacterial Viability Kit (Molecular Probes, Leiden, The Netherlands) was employed. Basically, biofilms were formed on polystyrene coupons, as described above, and were then stained for 15 min in the dark with a mixture of the SYTO 9 and Propidium Iodide, both prepared at 3 μl/mL in saline solution. For microscopic observation, an Olympus BX51 microscope fitted with fluorescence illumination was used. The optical filter combination consisted of 470 to 490 nm in combination with 530 to 550 nm excitation filters.
4.7. Statistical Analysis
Data were analyzed using the Prism software package (GraphPad Software version 6.01 for Macintosh). One-way analysis of variance (ANOVA) tests were performed, and means were compared by applying Tukey’s multiple comparison test. The statistical analyses performed were considered significant when p < 0.1. For all assays, at least three independent experiments were carried out in triplicate.






{kind=link}
{kind=link}
{kind=link}