Modified Aminoglycosides Bind Nucleic Acids in High-Molecular-Weight Complexes

 and
and
Abstract
:1. Introduction
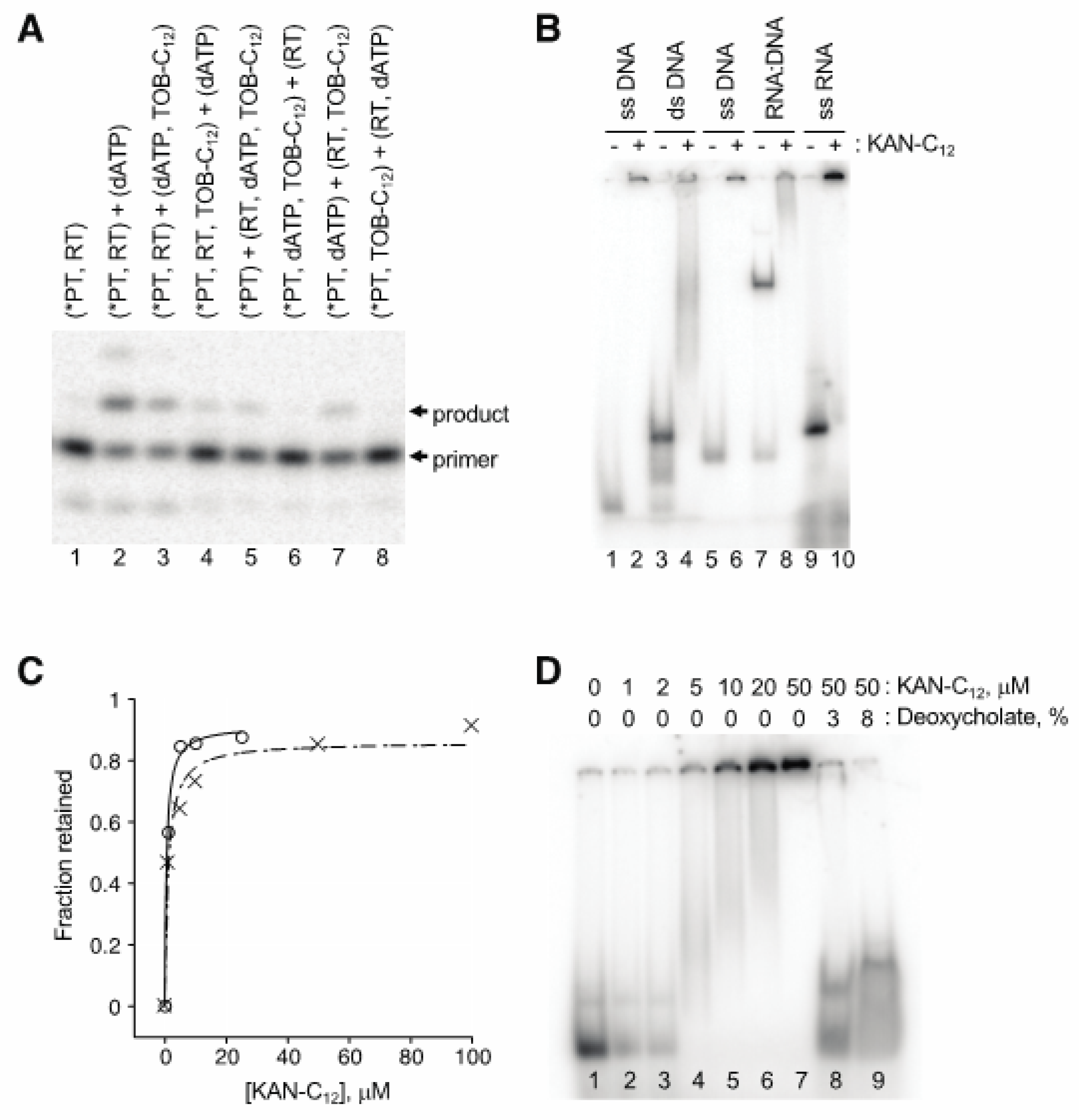
2. Results
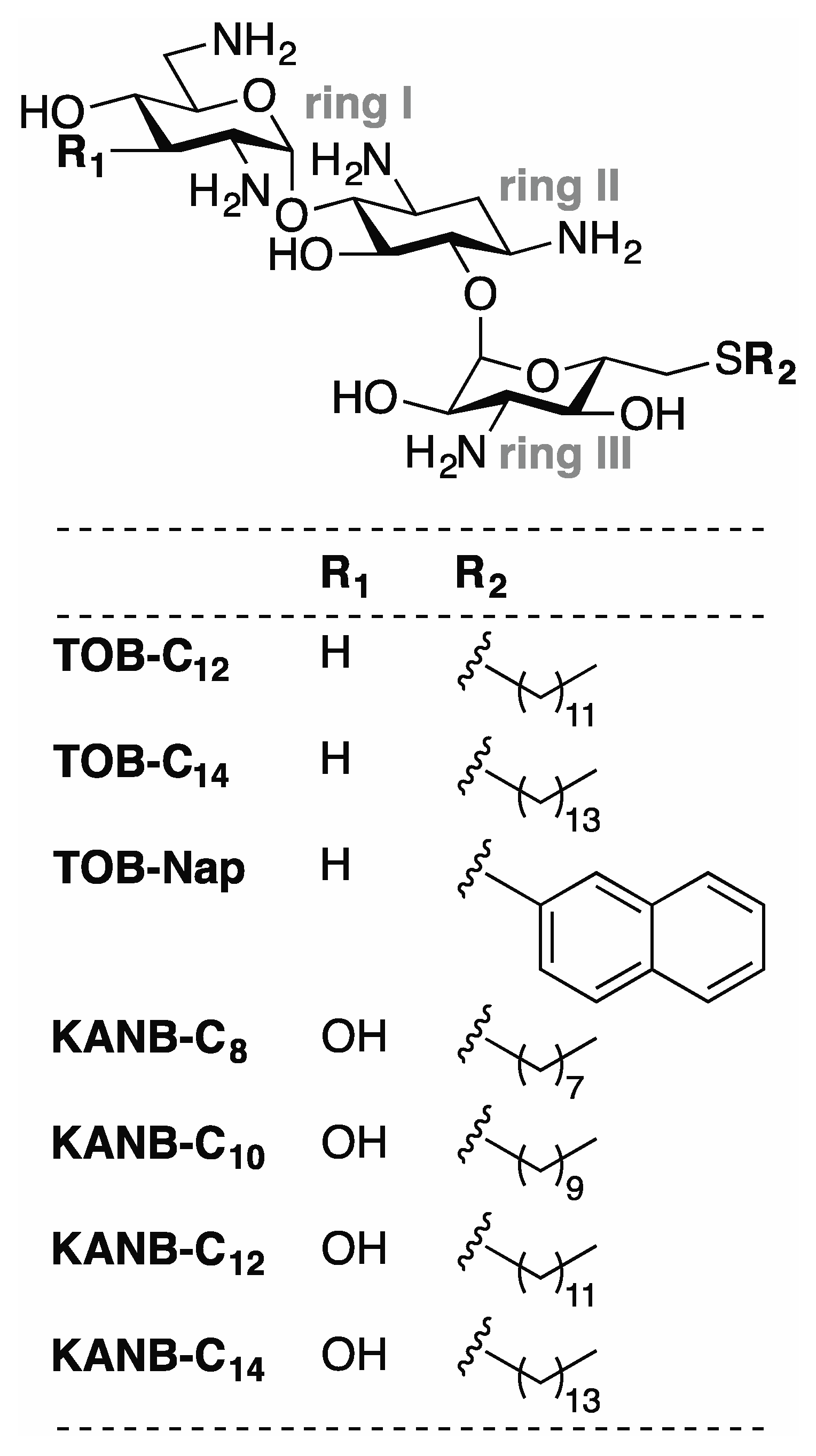
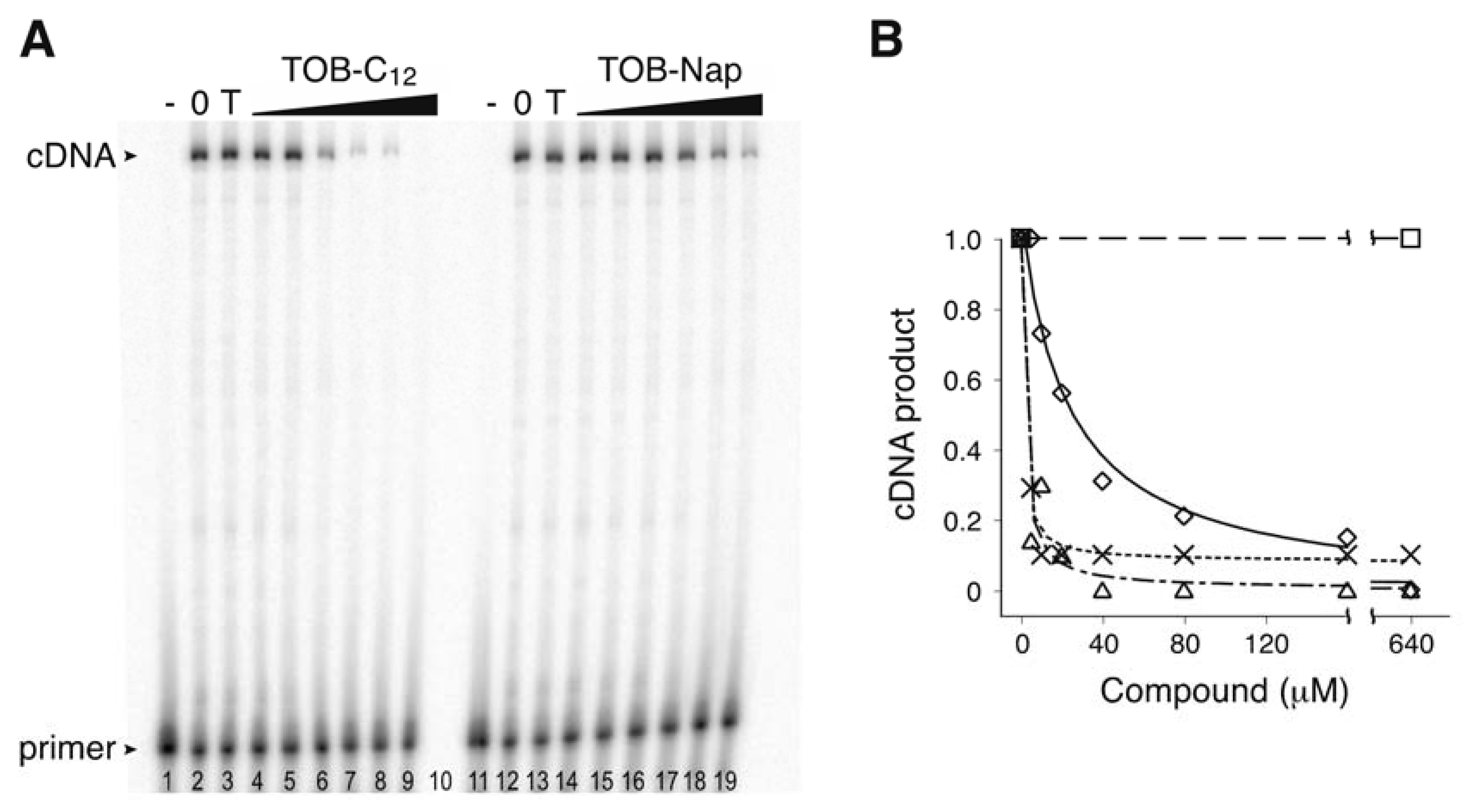
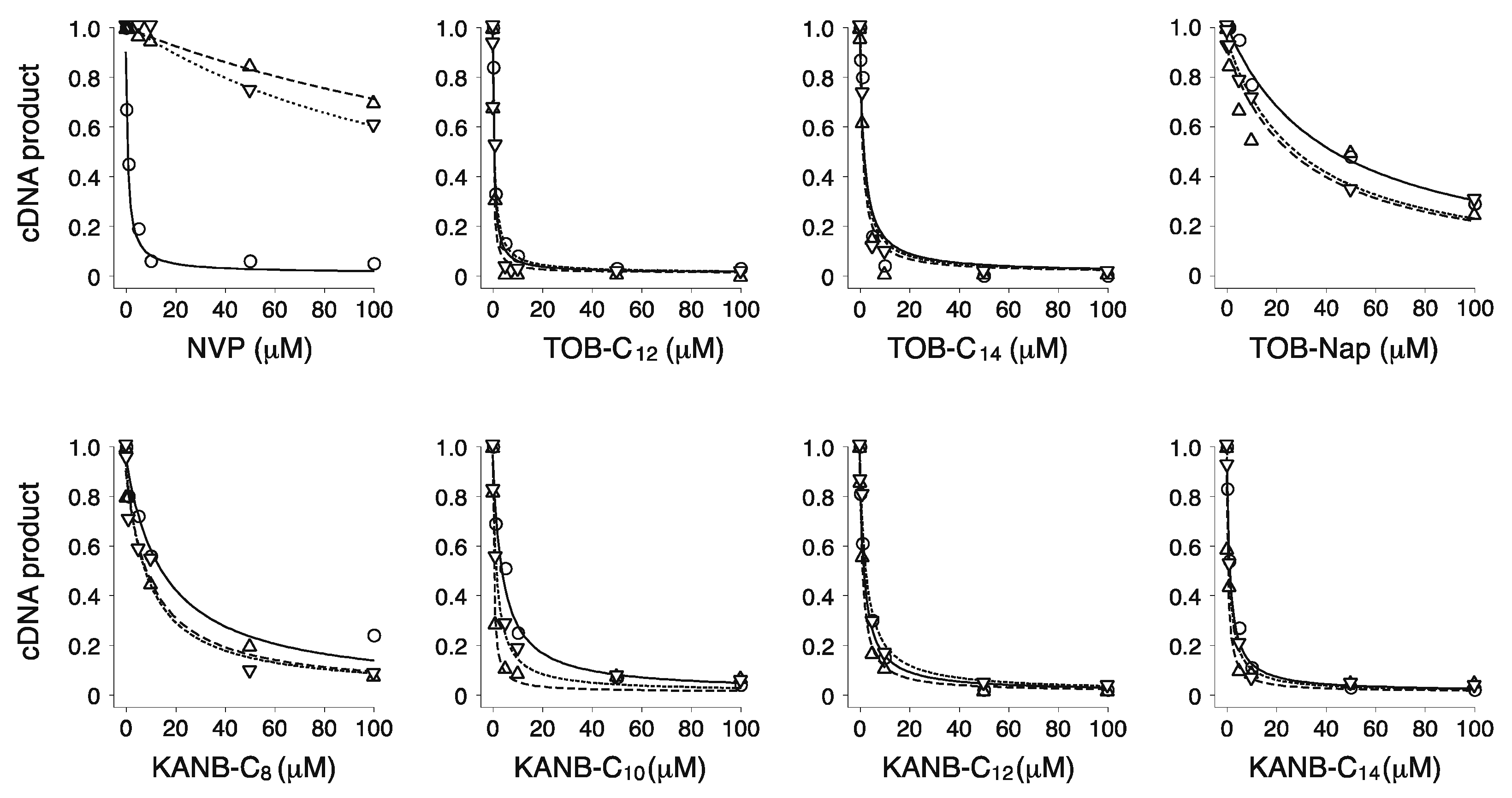
2.1. Potent Inhibition of Reverse Transcriptase by Certain Aminoglycoside Variants
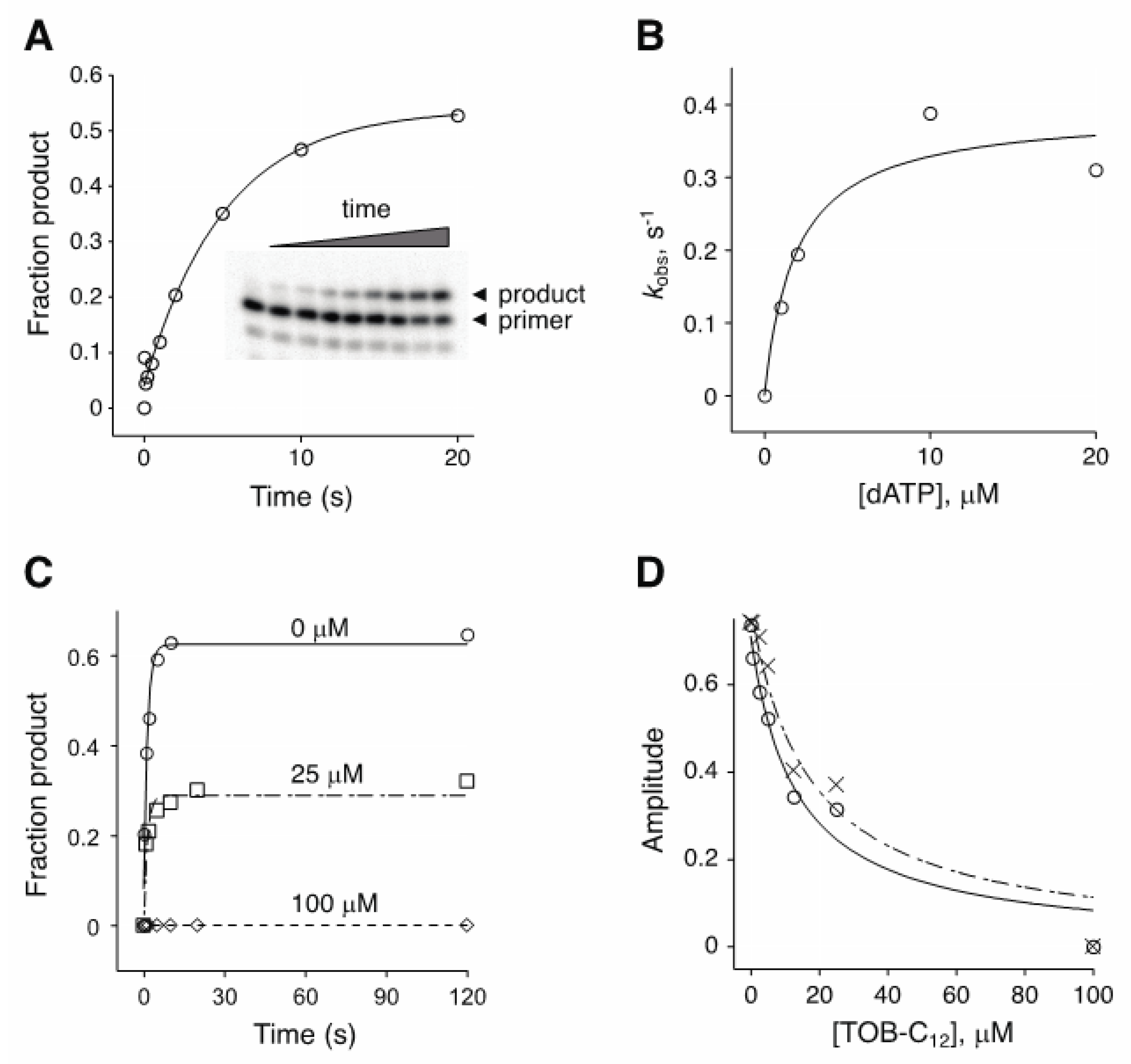
2.2. AG variants Bind Nucleic Acids in High-Molecular-Weight Complexes
3. Discussion
4. Materials and Methods
4.1. Biochemical Reagents
4.2. Primer Extension Assays
4.3. Binding Experiments
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carter, A.P.; Clemons, W.M.; Brodersen, D.E.; Morgan-Warren, R.J.; Wimberly, B.T.; Ramakrishnan, V. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 2000, 407, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Zhu, H.; Shoji, S.; Fredrick, K. Roles of specific aminoglycoside-ribosome interactions in the inhibition of translation. RNA 2019, 25, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Borovinskaya, M.A.; Pai, R.D.; Zhang, W.; Schuwirth, B.S.; Holton, J.M.; Hirokawa, G.; Kaji, H.; Kaji, A.; Cate, J.H. Structural basis for aminoglycoside inhibition of bacterial ribosome recycling. Nat. Struct. Mol. Biol. 2007, 14, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zhang, J.; Sun, W.; He, W.; Jiang, H.; Chen, D.; Murchie, A.I. Riboswitch control of aminoglycoside antibiotic resistance. Cell 2013, 152, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Von Ahsen, U.; Davies, J.; Schroeder, R. Antibiotic inhibition of group I ribozyme function. Nature 1991, 353, 368–370. [Google Scholar] [CrossRef]
- Ennifar, E.; Paillart, J.C.; Marquet, R.; Ehresmann, B.; Ehresmann, C.; Dumas, P.; Walter, P. HIV-1 RNA dimerization initiation site is structurally similar to the ribosomal A site and binds aminoglycoside antibiotics. J. Biol. Chem. 2003, 278, 2723–2730. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Huber, P.W.; Cui, M.; Czarnik, A.W.; Mei, H.Y. Binding of neomycin to the TAR element of HIV-1 RNA induces dissociation of Tat protein by an allosteric mechanism. Biochemistry 1998, 37, 5549–5557. [Google Scholar] [CrossRef]
- Kumar, S.; Ranjan, N.; Kellish, P.; Gong, C.; Watkins, D.; Arya, D.P. Multivalency in the recognition and antagonism of a HIV TAR RNA-TAT assembly using an aminoglycoside benzimidazole scaffold. Org. Biomol. Chem. 2016, 14, 2052–2056. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hamasaki, K.; Rando, R.R. Specificity of aminoglycoside binding to RNA constructs derived from the 16S rRNA decoding region and the HIV-RRE activator region. Biochemistry 1997, 36, 768–779. [Google Scholar] [CrossRef]
- Walter, F.; Vicens, Q.; Westhof, E. Aminoglycoside-RNA interactions. Curr. Opin. Chem. Biol. 1999, 3, 694–704. [Google Scholar] [CrossRef]
- Jiang, L.; Patel, D.J. Solution structure of the tobramycin-RNA aptamer complex. Nat. Struct. Biol. 1998, 5, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Blount, K.F.; Zhao, F.; Hermann, T.; Tor, Y. Conformational constraint as a means for understanding RNA-aminoglycoside specificity. J. Am. Chem. Soc. 2005, 127, 9818–9829. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.H.; Rando, R.R. Decoding region bubble size and aminoglycoside antibiotic binding. Bioorgan. Med. Chem. Lett. 2002, 12, 2241–2244. [Google Scholar] [CrossRef]
- Litovchick, A.; Evdokimov, A.G.; Lapidot, A. Arginine-aminoglycoside conjugates that bind to HIV transactivation responsive element RNA in vitro. FEBS Lett. 1999, 445, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Borkow, G.; Lara, H.H.; Lapidot, A. Mutations in gp41 and gp120 of HIV-1 isolates resistant to hexa-arginine neomycin B conjugate. Biochem. Biophys. Res. Commun. 2003, 312, 1047–1052. [Google Scholar] [CrossRef]
- Borkow, G.; Vijayabaskar, V.; Lara, H.H.; Kalinkovich, A.; Lapidot, A. Structure-activity relationship of neomycin, paromomycin, and neamine-arginine conjugates, targeting HIV-1 gp120-CXCR4 binding step. Antivir. Res. 2003, 60, 181–192. [Google Scholar] [CrossRef]
- Cabrera, C.; Gutierrez, A.; Blanco, J.; Barretina, J.; Litovchick, A.; Lapidot, A.; Evdokimov, A.G.; Clotet, B.; Este, J.A. Anti-human immunodeficiency virus activity of novel aminoglycoside-arginine conjugates at early stages of infection. Aids Res. Hum. Retrovir. 2000, 16, 627–634. [Google Scholar] [CrossRef]
- Cabrera, C.; Gutierrez, A.; Barretina, J.; Blanco, J.; Litovchick, A.; Lapidot, A.; Clotet, B.; Este, J.A. Anti-HIV activity of a novel aminoglycoside-arginine conjugate. Antivir. Res. 2002, 53, 1–8. [Google Scholar] [CrossRef]
- Hartz, D.; McPheeters, D.S.; Gold, L. Selection of the initiator tRNA by Escherichia coli initiation factors. Genes Dev. 1989, 3, 1899–1912. [Google Scholar] [CrossRef] [Green Version]
- Fosso, M.Y.; Zhu, H.; Green, K.D.; Garneau-Tsodikova, S.; Fredrick, K. Tobramycin Variants with Enhanced Ribosome-Targeting Activity. ChemBioChem 2015, 16, 1565–1570. [Google Scholar] [CrossRef]
- Brautigam, C.A.; Steitz, T.A. Structural and functional insights provided by crystal structures of DNA polymerases and their substrate complexes. Curr. Opin. Struct. Biol. 1998, 8, 54–63. [Google Scholar] [CrossRef]
- Doublie, S.; Tabor, S.; Long, A.M.; Richardson, C.C.; Ellenberger, T. Crystal structure of a bacteriophage T7 DNA replication complex at 2.2 A resolution. Nature 1998, 391, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Koonin, E.V.; Leipe, D.D.; Aravind, L. Origin and evolution of the archaeo-eukaryotic primase superfamily and related palm-domain proteins: Structural insights and new members. Nucleic Acids Res. 2005, 33, 3875–3896. [Google Scholar] [CrossRef] [PubMed]
- Jeruzalmi, D.; Steitz, T.A. Structure of T7 RNA polymerase complexed to the transcriptional inhibitor T7 lysozyme. EMBO J. 1998, 17, 4101–4113. [Google Scholar] [CrossRef] [Green Version]
- Reuman, E.C.; Rhee, S.Y.; Holmes, S.P.; Shafer, R.W. Constrained patterns of covariation and clustering of HIV-1 non-nucleoside reverse transcriptase inhibitor resistance mutations. J. Antimicrob. Chemother. 2010, 65, 1477–1485. [Google Scholar] [CrossRef]
- Xi, H.; Davis, E.; Ranjan, N.; Xue, L.; Hyde-Volpe, D.; Arya, D.P. Thermodynamics of nucleic acid "shape readout" by an aminosugar. Biochemistry 2011, 50, 9088–9113. [Google Scholar] [CrossRef] [Green Version]
- Sainlos, M.; Hauchecorne, M.; Oudrhiri, N.; Zertal-Zidani, S.; Aissaoui, A.; Vigneron, J.P.; Lehn, J.M.; Lehn, P. Kanamycin A-derived cationic lipids as vectors for gene transfection. ChemBioChem 2005, 6, 1023–1033. [Google Scholar] [CrossRef]
- Habrant, D.; Peuziat, P.; Colombani, T.; Dallet, L.; Gehin, J.; Goudeau, E.; Evrard, B.; Lambert, O.; Haudebourg, T.; Pitard, B. Design of Ionizable Lipids To Overcome the Limiting Step of Endosomal Escape: Application in the Intracellular Delivery of mRNA, DNA, and siRNA. J. Med. Chem. 2016, 59, 3046–3062. [Google Scholar] [CrossRef]
- Desigaux, L.; Sainlos, M.; Lambert, O.; Chevre, R.; Letrou-Bonneval, E.; Vigneron, J.P.; Lehn, P.; Lehn, J.M.; Pitard, B. Self-assembled lamellar complexes of siRNA with lipidic aminoglycoside derivatives promote efficient siRNA delivery and interference. Proc. Natl. Acad. Sci. USA 2007, 104, 16534–16539. [Google Scholar] [CrossRef] [Green Version]
- Hou, E.W.; Prasad, R.; Beard, W.A.; Wilson, S.H. High-level expression and purification of untagged and histidine-tagged HIV-1 reverse transcriptase. Protein Expr. Purif. 2004, 34, 75–86. [Google Scholar] [CrossRef]
- Shoji, S.; Walker, S.E.; Fredrick, K. Reverse translocation of tRNA in the ribosome. Mol. Cell 2006, 24, 931–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, I.M.; Green, K.D.; Berkov-Zrihen, Y.; Feldman, M.; Vidavski, R.R.; Eldar-Boock, A.; Satchi-Fainaro, R.; Eldar, A.; Garneau-Tsodikova, S.; Fridman, M. 6”-Thioether tobramycin analogues: Towards selective targeting of bacterial membranes. Angew. Chem. Int. Ed. 2012, 51, 5652–5656. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (µM) | ||
|---|---|---|---|
| WT | K103N | Y181C | |
| NVP | 0.3 ± 0.2 | 180 ± 40 | 200 ± 30 |
| TOB-C12 | 0.3 ± 0.1 | 0.6 ± 0.1 | 0.5 ± 0.1 |
| TOB-C14 | 1.9 ± 0.3 | 1.2 ± 0.6 | 1.4 ± 0.4 |
| TOB-Nap | 40 ± 5 | 40 ± 11 | 12 ± 4 |
| KANB-C8 | 8 ± 3 | 7 ± 1 | 8 ± 2 |
| KANB-C10 | 3.8 ± 0.3 | 1.9 ± 0.3 | 1.0 ± 0.6 |
| KANB-C12 | 1.9 ± 0.3 | 2.0 ± 0.2 | 0.5 ± 0.3 |
| KANB-C14 | 1.0 ± 0.1 | 0.9 ± 0.1 | 0.7 ± 0.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ying, L.; Zhu, H.; Fosso, M.Y.; Garneau-Tsodikova, S.; Fredrick, K. Modified Aminoglycosides Bind Nucleic Acids in High-Molecular-Weight Complexes. Antibiotics 2020, 9, 93. https://doi.org/10.3390/antibiotics9020093
Ying L, Zhu H, Fosso MY, Garneau-Tsodikova S, Fredrick K. Modified Aminoglycosides Bind Nucleic Acids in High-Molecular-Weight Complexes. Antibiotics. 2020; 9(2):93. https://doi.org/10.3390/antibiotics9020093
Chicago/Turabian StyleYing, Lanqing, Hongkun Zhu, Marina Y. Fosso, Sylvie Garneau-Tsodikova, and Kurt Fredrick. 2020. "Modified Aminoglycosides Bind Nucleic Acids in High-Molecular-Weight Complexes" Antibiotics 9, no. 2: 93. https://doi.org/10.3390/antibiotics9020093
APA StyleYing, L., Zhu, H., Fosso, M. Y., Garneau-Tsodikova, S., & Fredrick, K. (2020). Modified Aminoglycosides Bind Nucleic Acids in High-Molecular-Weight Complexes. Antibiotics, 9(2), 93. https://doi.org/10.3390/antibiotics9020093