A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility, Carbapenemase Phenotype Detection and MLST
2.2. Whole-Genome Sequencing and in Silico Data Analysis
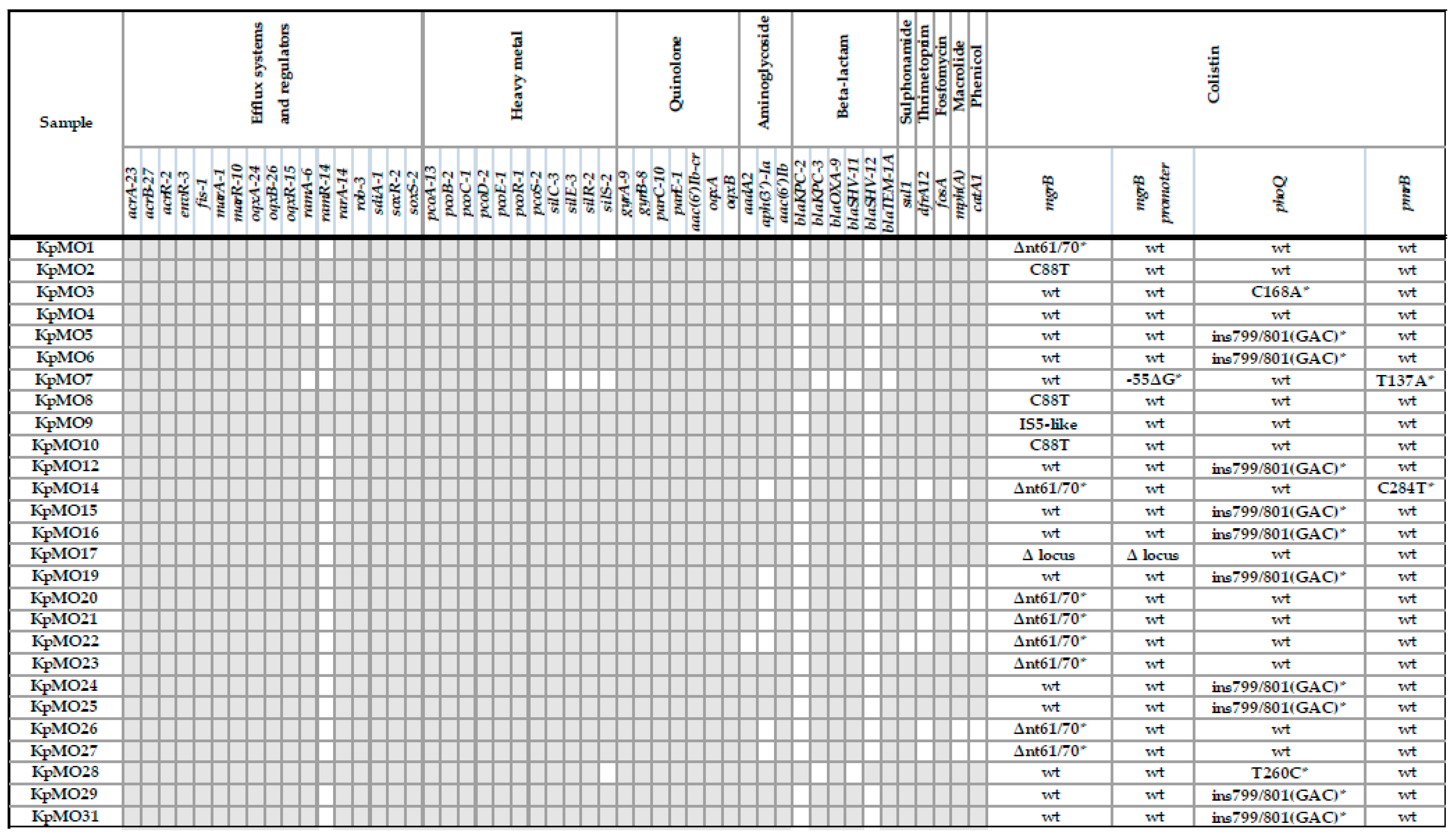
2.2.1. Resistome Analysis
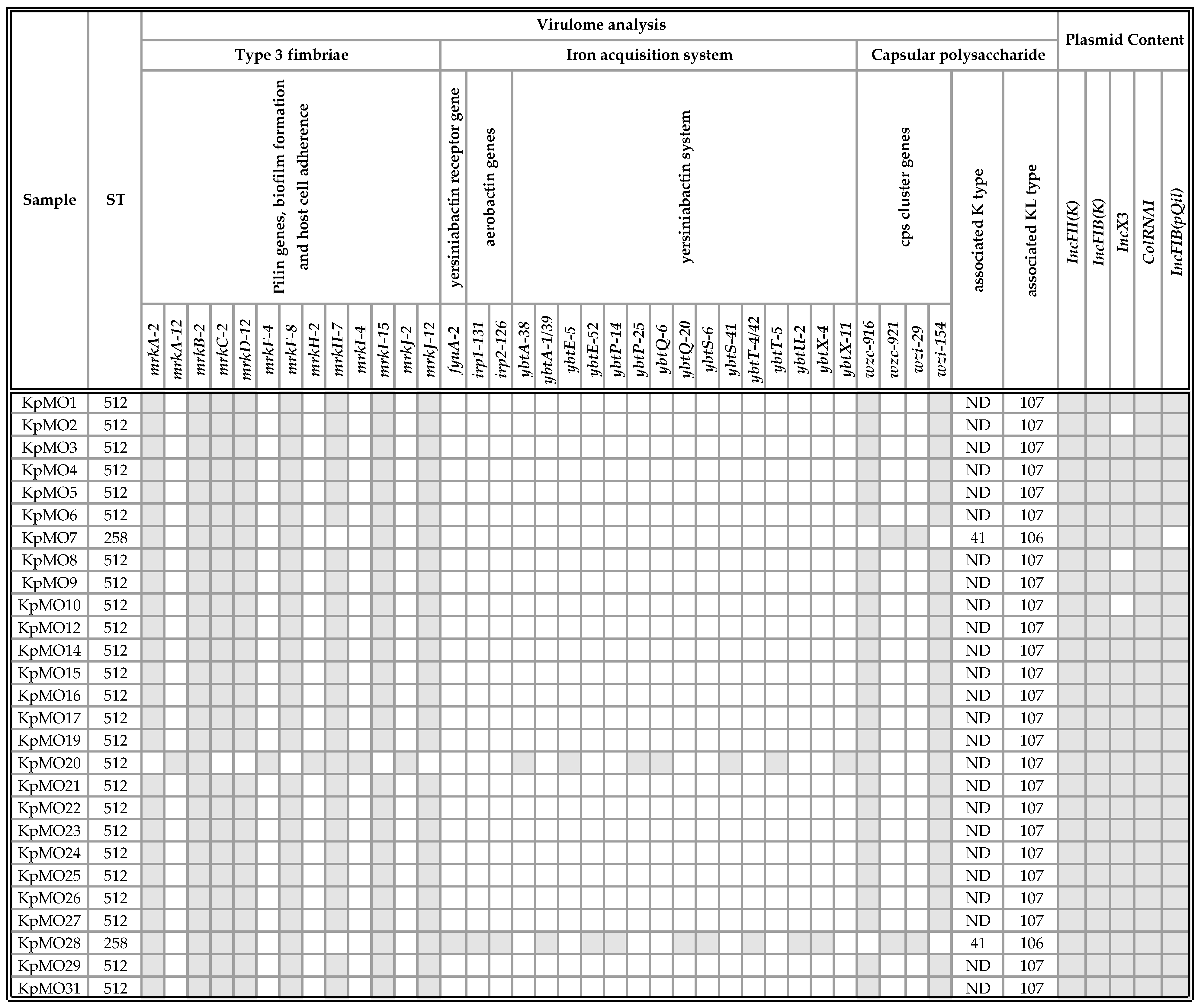
2.2.2. Virulome Analysis
2.2.3. Plasmid Content Analysis
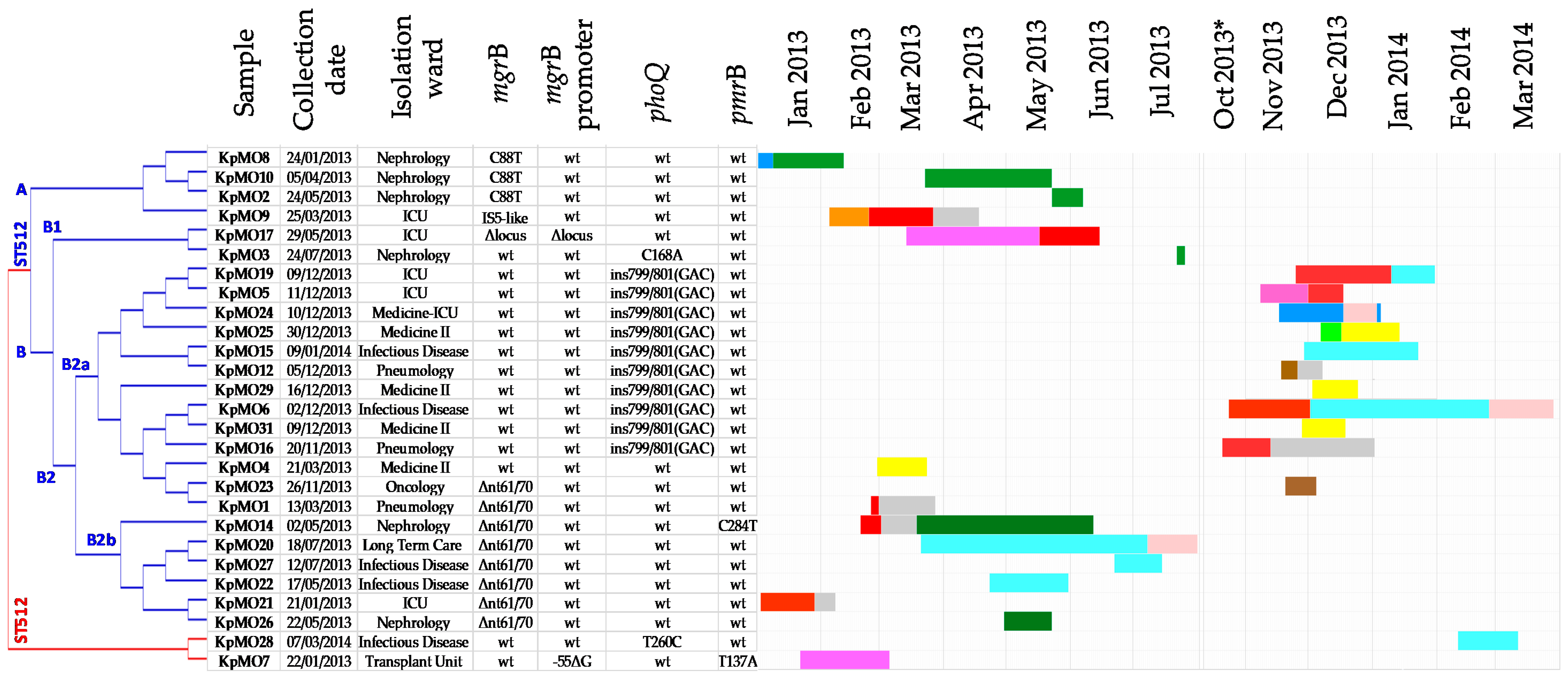
2.2.4. Phylogenetic Analysis
2.3. The Molecular Data in the Clinical–Epidemiologic Context
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Antimicrobial Susceptibility, Carbapenemase Phenotype Detection and MLST
4.3. Whole-Genome Sequencing and in Silico Data Analysis
4.3.1. Resistome, Virulome, and Plasmid Content Analysis
4.3.2. Phylogenetic Analysis
4.4. The Molecular Data in the Clinical–Epidemiologic Context
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-producing klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Gaiarsa, S.; Comandatore, F.; Gaibani, P.; Corbella, M.; Valle, C.D.; Epis, S.; Scaltriti, E.; Carretto, E.; Farina, C.; Labonia, M.; et al. Genomic epidemiology of Klebsiella pneumoniae in Italy and novel insights into the origin and global evolution of its resistance to carbapenem antibiotics. Antimicrob. Agents Chemother. 2015, 59, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Conte, V.; Monaco, M.; Giani, T.; D’Ancona, F.; Moro, M.L.; Arena, F. Molecular epidemiology of KPC-producing Klebsiella pneumoniae from invasive infections in Italy: Increasing diversity with predominance of the ST512 clade II sublineage. J. Antimicrob. Chemother. 2016, 71, 3386–3391. [Google Scholar] [CrossRef] [Green Version]
- Rimoldi, S.G.; Gentile, B.; Pagani, C.; Di Gregorio, A.; Anselmo, A.; Palozzi, A.M.; Fortunato, A.; Pittiglio, V.; Ridolfo, A.L.; Gismondo, M.; et al. Whole genome sequencing for the molecular characterization of carbapenemresistant Klebsiella pneumoniae strains isolated at the Italian ASST Fatebenefratelli Sacco Hospital, 2012–2014. BMC Infect. Dis. 2017, 17, 666. [Google Scholar] [CrossRef]
- Fasciana, T.; Gentile, B.; Aquilina, M.; Ciammaruconi, A.; Mascarella, C.; Anselmo, A.; Fortunato, A.; Fillo, S.; Petralito, G.; Lista, F.; et al. Co-existence of virulence factors and antibiotic resistance in new Klebsiella pneumoniae clones emerging in south of Italy. BMC Infect. Dis. 2019, 19, 928. [Google Scholar] [CrossRef] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rello, J.; Eshwara, V.K.; Lagunes, L.; Alves, J.; Wunderink, R.G.; Morris, A.C.; Rojas, J.N.; Alp, E.; Zhang, Z. A global priority list of the TOp TEn resistant Microorganisms (TOTEM) study at intensive care: A prioritization exercise based on multi-criteria decision analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Monaco, M.; Giani, T.; Raffone, M.; Arena, F.; Garcia-Fernandez, A.; Pollini, S.; Grundmann, H.; Pantosti, A.; Rossolini, G.M. Colistin resistance superimposed to endemic carbapenem-resistant Klebsiella pneumoniae: A rapidly evolving problem in Italy, November 2013 to April 2014. Eurosurveillance 2014, 19, 20939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, E.P.; Cervoni, M.; Bernardo, M.; Crivaro, V.; Cuccurullo, S.; Imperi, F.; Zarrilli, R. Molecular epidemiology and virulence profiles of colistin-resistant Klebsiella pneumoniae blood isolates from the hospital agency “Ospedale dei Colli”, Naples, Italy. Front Microbiol. 2018, 9, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaitan, A.; Diene, S.M.; Kempf, M.; Berrazeg, M.; Bakour, S.; Gupta, S.; Thongmalayvong, B.; Akkhavong, K.; Somphavong, S.; Paboriboune, P.; et al. Worldwide emergence of colistin resistance in Klebsiella pneumoniae from healthy humans and patients in Lao PDR, Thailand, Israel, Nigeria and France owing to inactivation of the PhoP/PhoQ regulator mgrB: An epidemiological and molecular study. Int. J. Antimicrob. Agents. 2014, 44, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefaniuk, E.M.; Tyski, S. Colistin Resistance in Enterobacterales Strains—A Current View. Pol. J. Microbiol. 2019, 68, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabat, A.J.; Budimir, A.; Nashev, D.; Sá-Leão, R.; van Dijl, J.M.; Laurent, F.; Grundmann, H.; Friedrich, A.W. Overview of molecular typing methods for outbreak detection and epidemiological surveillance. Eurosurveillance 2013, 18, 20380. [Google Scholar]
- Quainoo, S.; Coolen, J.P.M.; Van Hijum, S.A.; Huynen, M.A.; Melchers, W.J.G.; Van Schaik, W.; Wertheim, H.F.L. Whole-genome sequencing of bacterial pathogens: The future of nosocomial outbreak analysis. Clin. Microbiol. Rev. 2017, 30, 1015–1063. [Google Scholar] [CrossRef] [Green Version]
- Mavroidi, A.; Katsiari, M.; Likousi, S.; Palla, E.; Roussou, Z.; Nikolaou, C.; Maguina, A.; Platsouka, E.D. Characterization of ST258 Colistin-Resistant, blaKPC-Producing Klebsiella pneumoniae in a Greek Hospital. Microb. Drug Resist. 2016, 22, 392–398. [Google Scholar] [CrossRef]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [Green Version]
- Calia, C.; Pazzani, C.; Oliva, M.; Scrascia, M.; Lovreglio, P.; Capolongo, C.; Dionisi, A.M.; Chiarelli, A.; Monno, R. Carbapenemases-producing Klebsiella pneumoniae in hospitals of two regions of Southern Italy. APMIS 2017, 21, 45–498. [Google Scholar] [CrossRef]
- Del Bianco, F.; Morotti, M.; Pedna, M.F.; Farabegoli, P.; Sambri, V. Microbiological surveillance of plasmid mediated colistin resistance in human Enterobacteriaceae isolates in Romagna (Northern Italy): August 2016–July 2017. Int. J. Infect. Dis. 2018, 69, 96–98. [Google Scholar] [CrossRef] [Green Version]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB inactivation is a common mechanism of colistin resistance in KPC-producing Klebsiella pneumoniae of clinical origin. Antimicrob. Agents Chemother. 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Bontron, S.; Villegas, M.-V.; Ozdamar, M.; Nordmann, P.; Türkoglu, S. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2014, 70, 75–80. [Google Scholar] [CrossRef]
- Ong, C.-L.Y.; Beatson, S.A.; Totsika, M.; Forestier, C.; McEwan, A.G.; Schembri, M.A. Molecular analysis of type 3 fimbrial genes from Escherichia coli. Klebsiella and Citrobacter species. BMC Microbiol. 2010, 10, 183. [Google Scholar]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.-S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.-H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K. Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2016, 2, e000102. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Volby Larsen, M.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. PlasmidFinder and pMLST: In silico detection and typing of plasmids. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Phan, H.T.; Findlay, J.; Stoesser, N.; Pankhurst, L.; Navickaite, I.; De Maio, N.; Eyre, D.W.; Toogood, G.; Orsi, N.M.; et al. Covert dissemination of carbapenemase-producing Klebsiella pneumoniae (KPC) in a successfully controlled outbreak: Long-and short-read whole-genome sequencing demonstrate multiple genetic modes of transmission. J. Antimicrob. Chemother. 2017, 72, 3025–3034. [Google Scholar] [CrossRef] [PubMed]
- EUCAST (European Committee on Antimicrobial Susceptibility Testing). BreakpoInt. Tables for Interpretation of MICs and Zone Diameters. Version 2.0, valid from 2012–01-01. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/Breakpoint_table_v_2.0_120221.pdf (accessed on 31 March 2014).
- Simpson, J.T.; Wong, K.; Jackman, S.; Schein, J.E.; Jones, S.J.M.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Chan, A.P. Provean web server: A tool to predict the functional effect of amminoacid substitution and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.N.; Slezak, T.; Hall, B.G. kSNP3.0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Boil. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Schürch, A.C.; van Schaik, W. Challenges and opportunities for whole-genome sequencing–based surveillance of antibiotic resistance. Ann. N. Y. Acad. Sci. 2017, 1388, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Mellmann, A.; Bletz, S.; Böking, T.; Kipp, F.; Becker, K.; Schultes, A.; Prior, K.; Harmsen, D. Real-time genome sequencing of resistant bacteria provides precision infection control in an institutional setting. J. Clin. Microbiol. 2016, 54, 2874–2881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, H.C.D.; Allard, M.W.; Bopp, D.; Brown, E.W.; Fontana, J.; Iqbal, Z.; Kinney, A.; Limberger, R.; Musser, K.A.; Shudt, M.; et al. Rapid whole-genome sequencing for surveillance of Salmonella enterica serovar enteritidis. Emerg. Infect. Dis. 2014, 20, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Dallman, T.J.; Byrne, L.; Ashton, P.M.; Cowley, L.; Perry, N.T.; Adak, G.; Petrovska, L.; Ellis, R.J.; Elson, R.; Underwood, A.; et al. Whole-genome sequencing for national surveillance of Shiga toxinproducing Escherichia coli O157. Clin. Infect. Dis. 2015, 61, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, J.C.; Lane, C.R.; Romanes, F.; Da Silva, A.G.; Easton, M.; Cronin, K.; Waters, M.J.; Tomita, T.; Stevens, K.; Schultz, M.B.; et al. Translating genomics into practice for real-time surveillance and response to carbapenemase-producing Enterobacteriaceae: Evidence from a complex multi-institutional KPC outbreak. PeerJ 2018, 6, e4210. [Google Scholar] [CrossRef]
- Balloux, F.; Brynildsrud, O.; Van Dorp, L.; Shaw, L.P.; Chen, H.; Harris, K.; Wang, H.; Eldholm, V. From Theory to Practice: Translating Whole-Genome Sequencing (WGS) into the Clinic. Trends Microbiol. 2018, 26, 1035–1048. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Isolate ID | GEN | AMK | IPM | MEM | ETP | CIP | TMP/SMX | TGC | COL | AMC | TZP | AMP | CPM | CTX | FOX | CAZ | FOS | Carbapenemase Class (Enzyme) | MLST |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KpMO1 | A | 512 | |||||||||||||||||
| KpMO2 | A | 512 | |||||||||||||||||
| KpMO3 | A | 512 | |||||||||||||||||
| KpMO4 | A | 512 | |||||||||||||||||
| KpMO5 | A | 512 | |||||||||||||||||
| KpMO6 | A | 512 | |||||||||||||||||
| KpMO7 | A | 258 | |||||||||||||||||
| KpMO8 | A | 512 | |||||||||||||||||
| KpMO9 | A | 512 | |||||||||||||||||
| KpMO10 | A | 512 | |||||||||||||||||
| KpMO12 | A | 512 | |||||||||||||||||
| KpMO14 | A | 512 | |||||||||||||||||
| KpMO15 | A | 512 | |||||||||||||||||
| KpMO16 | A | 512 | |||||||||||||||||
| KpMO17 | A | 512 | |||||||||||||||||
| KpMO19 | A | 512 | |||||||||||||||||
| KpMO20 | A | 512 | |||||||||||||||||
| KpMO21 | A | 512 | |||||||||||||||||
| KpMO22 | A | 512 | |||||||||||||||||
| KpMO23 | A | 512 | |||||||||||||||||
| KpMO24 | A | 512 | |||||||||||||||||
| KpMO25 | A | 512 | |||||||||||||||||
| KpMO26 | A | 512 | |||||||||||||||||
| KpMO27 | A | 512 | |||||||||||||||||
| KpMO28 | A | 258 | |||||||||||||||||
| KpMO29 | A | 512 | |||||||||||||||||
| KpMO31 | A | 512 |
| Gene | Nucleotide Mutation | Protein Variant | PROVEAN Score | Prediction (Cutoff = −2.5) |
|---|---|---|---|---|
| mgrB | Δnt61/70 | T21L | −2996 | Deleterious |
| Q22T | −6000 | Deleterious | ||
| M23_W47del | −125,068 | Deleterious | ||
| phoQ | C168A | S56R | −3.359 | Deleterious |
| ins799/801(GAC) | D266_K267insD | −8.067 | Deleterious | |
| T260C | L87P | −4.65 | Deleterious | |
| pmrB | T137A | V46E | −4.063 | Deleterious |
| C284T | P95L | −9.604 | Deleterious |
| Isolate ID | Sample Type a | Collection Date | Age | Gender | CCI b | Colistin Use* (Days) | Admission Ward c (Date) | 2nd Admission Ward (Date) | 3rd Admission Ward (Date) | Discharge/Death (Date) | Clinical Outcome |
|---|---|---|---|---|---|---|---|---|---|---|---|
| KpMO1 | rectal swab | 13/03/2013 | 82 | M | 3 | no | ICU (25/02/2013) | Pneumology (01/03/2013) | Pneumology (27/03/2013) | recover | |
| KpMO2 | rectal swab | 24/05/2013 | 68 | F | 3 | no | Nephrology (23/05/2013) | − | Nephrology (06/06/2013) | chronic disease | |
| KpMO3 | rectal swab | 24/07/2013 | 63 | M | 5 | no | Nephrology (22/07/2013) | Nephrology (25/07/2013) | chronic disease | ||
| KpMO4 | rectal swab | 21/03/2013 | 77 | M | 2 | yes (13) | Medicine II (28/02/2013) | Medicine II (23/03/2013) | death for sepsis | ||
| KpMO5 | rectal swab | 11/12/2013 | 78 | F | 3 | no | Transplant Unit (08/11/2013) | ICU (01/12/2013) | ICU (17/12/2013) | death for non-infectious cause | |
| KpMO6 | rectal swab | 02/12/2013 | 67 | M | 4 | no | ICU (24/10/2013) | Infectious Disease (02/12/2013) | Long Term Care (26/02/2014) | Long Term Care (28/03/2014) | chronic disease |
| KpMO7 | rectal swab | 22/01/2013 | 71 | F | 2 | no | Transplant Unit (22/01/2013) | Transplant Unit (05/03/2013) | recover | ||
| KpMO8 | rectal swab | 24/01/2013 | 68 | M | 6 | no | Medicine-ICU (26/12/2012) | Nephrology (09/01/2013) | Nephrology (11/02/2013) | chronic disease | |
| KpMO9 | rectal swab | 25/03/2013 | 73 | F | 2 | no | Otolaryngology 05/02/2013 | ICU (24/02/2013) | Pneumology (27/03/2013) | Pneumology (17/04/2013) | recover |
| KpMO10 | urine | 05/04/2013 | 57 | M | 3 | no | Nephrology (23/03/2013) | Nephrology (22/05/2013) | recover | ||
| KpMO12 | rectal swab | 05/12/2013 | 70 | F | 8 | no | Oncology (18/11/2013) | Pneumology (26/11/2013) | Pneumology (07/12/2013) | death for non-infectious cause | |
| KpMO14 | rectal swab | 02/05/2013 | 55 | F | 4 | yes (30) | ICU (20/02/2013) | Pneumology (02/03/2013) | Nephrology (19/03/2013) | Nephrology (11/06/2013) | chronic disease |
| KpMO15 | rectal swab | 09/01/2014 | 46 | F | 6 | no | Infectious Disease (30/11/2013) | Infectious Disease (22/01/2014) | recover | ||
| KpMO16 | blood | 20/11/2013 | 73 | M | 3 | no | ICU (21/10/2013) | Pneumology (13/11/2013) | Pneumology (01/01/2014) | death for sepsis | |
| KpMO17 | BAL | 29/05/2013 | 47 | M | 3 | yes (12) | Transplant Unit (14/03/2013) | ICU (17/05/2013) | ICU (14/06/2013) | death for sepsis | |
| KpMO19 | rectal swab | 09/12/2013 | 66 | M | 1 | yes (14) | ICU (25/11/2013) | Infectious Disease (10/01/2014) | Infectious Disease (30/01/2014) | recover | |
| KpMO20 | urine | 18/07/2013 | 80 | F | 2 | no | Infectious Disease (21/03/2013) | Long Term Care (08/07/2013) | Long Term Care (30/07/2013) | chronic disease | |
| KpMO21 | rectal swab | 21/01/2013 | 43 | M | 2 | yes (26) | ICU (03/01/2013) | Pneumology (29/01/2013) | Pneumology (07/02/2013) | death for non-infectious cause | |
| KpMO22 | rectal swab | 17/05/2013 | 91 | M | 2 | no | Infectious Disease (23/04/2013) | Infectious Disease (30/05/2013) | chronic disease | ||
| KpMO23 | rectal swab | 26/11/2013 | 76 | F | 2 | no | Oncology (20/11/2013) | Oncology (04/12/2013) | chronic disease | ||
| KpMO24 | rectal swab | 10/12/2013 | 88 | M | 4 | no | Medicine-ICU (17/11/2013) | Long Term Care (18/12/2013) | Medicine-ICU (03/01/2014) | Medicine-ICU (04/01/2014) | death for sepsis |
| KpMO25 | rectal swab | 30/12/2013 | 91 | M | 11 | no | Orthopaedics (07/12/2013) | Medicine II (17/12/2013) | Medicine II (13/01/2014) | recover | |
| KpMO26 | rectal swab | 22/05/2013 | 89 | F | 2 | no | Nephrology (30/04/2013) | Nephrology (22/05/2013) | chronic disease | ||
| KpMO27 | rectal swab | 12/07/2013 | 94 | F | 4 | no | Infectious Disease (22/06/2013) | Infectious Disease (14/07/2013) | death for sepsis | ||
| KpMO28 | urine | 07/03/2014 | 24 | M | 6 | no | Infectious Disease (11/02/2014) | Infectious Disease (11/03/2014) | recover | ||
| KpMO29 | rectal swab | 16/12/2013 | 60 | F | 1 | no | Medicine II (03/12/2013) | Medicine II (24/12/2013) | recover | ||
| KpMO31 | rectal swab | 09/12/2013 | 85 | M | 4 | no | Medicine II (28/11/2013) | Medicine II (18/12/2013) | death for non-infectious cause |
| Head | Reference Sequences | GenBank Accession Number | |
|---|---|---|---|
| Chromosomal loci | acrAB | K. pneumoniae subsp. pneumoniae ST512-K30BO (Kp-ST512-K30BO) | NZ_CAJM00000000.2 |
| pmrHFIJKLM | |||
| crrAB | |||
| KpnEF | |||
| lpxM | − | − | |
| mgrB | K. pneumoniae subsp. pneumoniae HS11286 | CP003200.1 | |
| mgrB promoter | |||
| phoPQ | |||
| pmrABCD |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentile, B.; Grottola, A.; Orlando, G.; Fregni Serpini, G.; Venturelli, C.; Meschiari, M.; Anselmo, A.; Fillo, S.; Fortunato, A.; Lista, F.; et al. A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program. Antibiotics 2020, 9, 246. https://doi.org/10.3390/antibiotics9050246
Gentile B, Grottola A, Orlando G, Fregni Serpini G, Venturelli C, Meschiari M, Anselmo A, Fillo S, Fortunato A, Lista F, et al. A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program. Antibiotics. 2020; 9(5):246. https://doi.org/10.3390/antibiotics9050246
Chicago/Turabian StyleGentile, Bernardina, Antonella Grottola, Gabriella Orlando, Giulia Fregni Serpini, Claudia Venturelli, Marianna Meschiari, Anna Anselmo, Silvia Fillo, Antonella Fortunato, Florigio Lista, and et al. 2020. "A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program" Antibiotics 9, no. 5: 246. https://doi.org/10.3390/antibiotics9050246
APA StyleGentile, B., Grottola, A., Orlando, G., Fregni Serpini, G., Venturelli, C., Meschiari, M., Anselmo, A., Fillo, S., Fortunato, A., Lista, F., Pecorari, M., & Mussini, C. (2020). A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program. Antibiotics, 9(5), 246. https://doi.org/10.3390/antibiotics9050246