Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Fluoroquinolone Resistance
2.2. Erythromycin Resistance
2.3. Tetracycline Resistance
2.4. Streptomycin Resistance
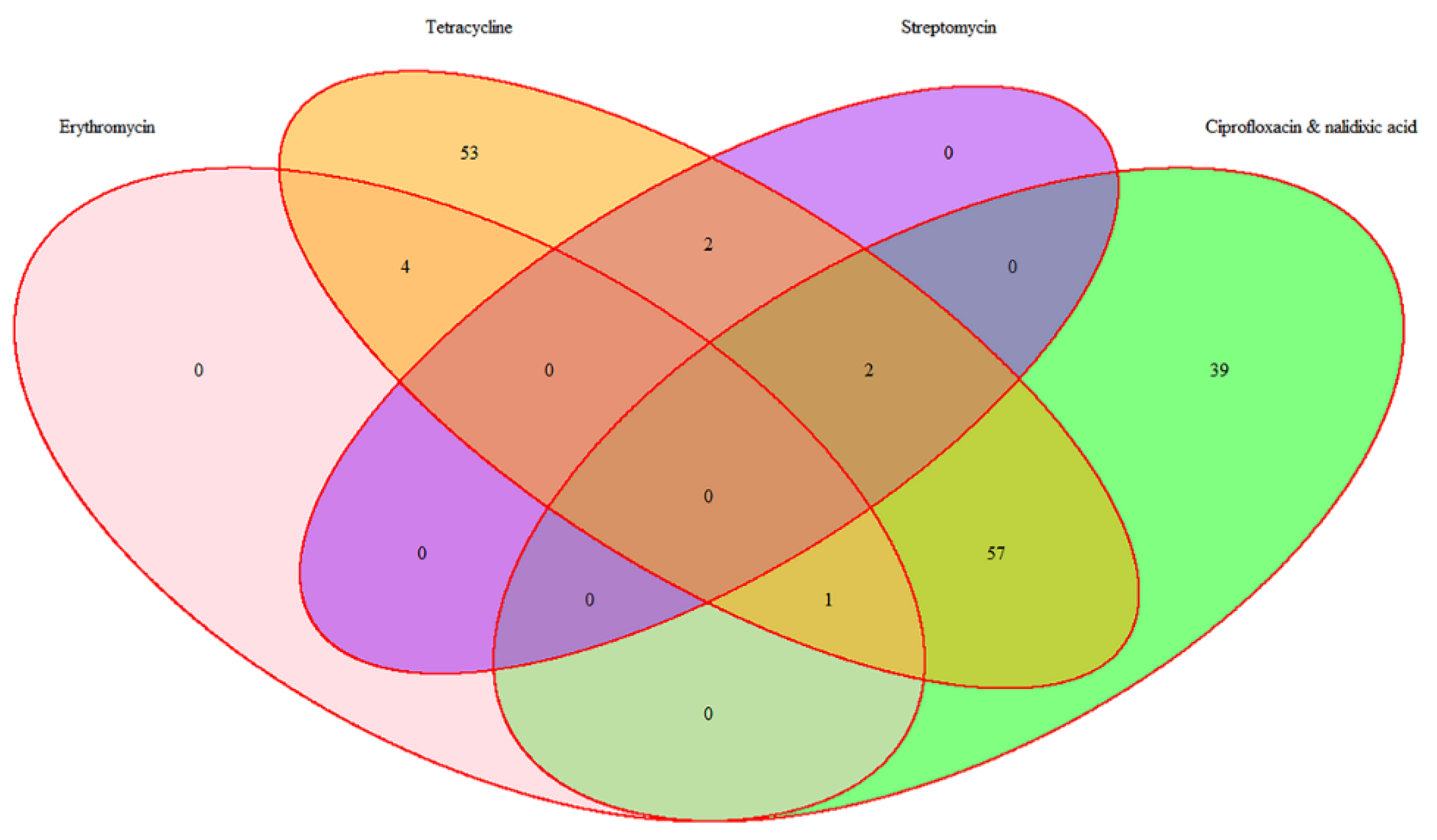
2.5. Overall Distribution of Antimicrobial Resistance
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolate Culture Conditions and Susceptibility Testing
4.2. DNA Extraction
4.3. Genotypic Characterisation of Antimicrobial Resistance—PCR Amplification and Sequencing
4.4. Moxifloxacin Minimum Inhibitory Concentration Testing
4.5. Genome Sequencing and Genomic Analysis
4.6. Data Availability
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority. Scientific Opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 94, 2105. [Google Scholar]
- European Food Safety Authority. Analysis of the baseline survey on the prevalence of Campylobacter in broiler batches and of Campylobacter and Salmonella on broiler carcasses in the EU. EFSA J. 2010, 8, 1522. [Google Scholar]
- World Health Organisation Critically Important Antimicrobials for Human Medicine 6th Revision. 2018. Available online: https://www.who.int/foodsafety/publications/antimicrobials-sixth/en/ (accessed on 1 April 2020).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, aimals and food in 2017. EFSA J. 2019, 17, e04694. [Google Scholar]
- European Food Safety Authority; European Centre for Disease Prevention and Control. European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, 4380. [Google Scholar]
- Duarte, A.; Santos, A.; Manageiro, V.; Martins, A.; Fraqueza, M.J.; Caniç, M.; Domingues, F.C.; Oleastro, M. Human, food and animal Campylobacter spp. isolated in Portugal: High genetic diversity and antibiotic resistance rates. Int. J. Antimicrob. Agents 2014, 44, 306–313. [Google Scholar] [CrossRef]
- Alfredson, D.A.; Korolik, V. Antibiotic resistance and resistance mechanisms in Campylobacter jejuni and Campylobacter coli. FEMS Microbiol. Lett. 2007, 227, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Bolinger, H.; Kathariou, S. The current state of macrolide resistance in Campylobacter spp.: Trends and impacts of resistance mechanisms. Appl. Environ. Microbiol. 2017, 83, e00416–e00417. [Google Scholar] [CrossRef] [Green Version]
- Aarestrup, F.M.; Engberg, J. Antimicrobial resistance of thermophilic Campylobacter. Vet. Res. 2001, 32, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, H.; Hansen, K.K.; Gradel, K.O.; Kristensen, B.; Ejlertsen, T.; Østergaard, C.; Schønheyder, H.C. Bacteraemia as a result of Campylobacter species: A population-based study of epidemiology and clinical risk factors. Clin. Microbiol. Infect. 2010, 16, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Payot, S.; Bolla, J.-M.; Corcoran, D.; Fanning, S.; Mégraud, F.; Zhang, Q. Mechanisms of fluoroquinolone and macrolide resistance in Campylobacter spp. Microbes Infect. 2006, 8, 1967–1971. [Google Scholar] [CrossRef]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol. Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Klostermeier, D. Why two? On the role of (A-)symmetry in negative supercoiling of DNA by gyrase. Int. J. Mol. Sci. 2018, 19, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nasht, H.A. DNA gyrase: An enzyme that introduces superhelical turns into DNA (Escherichia coli/ATP-dependent reaction/superhelix density). Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zirnstein, G.; Li, Y.; Swaminathan, B.; Angulo, F. Ciprofloxacin resistance in Campylobacter jejuni isolates: Detection of gyrA resistance mutations by mismatch amplification mutation assay PCR and DNA sequence analysis. J. Clin. Microbiol. 1999, 37, 3276–3280. [Google Scholar] [CrossRef] [Green Version]
- Siibak, T.; Peil, L.; Xiong, L.; Mankin, A.; Remme, J.; Tenson, T. Erythromycin- and chloramphenicol-induced ribosomal assembly defects are secondary effects of protein synthesis inhibition. Antimicrob. Agents Chemother. 2009, 53, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Cagliero, C.; Mouline, C.; Cloeckaert, A.; Payot, S. Synergy between efflux pump CmeABC and modifications in ribosomal proteins L4 and L22 in conferring macrolide resistance in Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 2006, 50, 3893–3896. [Google Scholar] [CrossRef] [Green Version]
- Zengel, J.M.; Jerauld, A.; Walker, A.; Wahl, M.C.; Lindahl, L. The extended loops of ribosomal proteins L4 and L22 are not required for ribosome assembly or L4-mediated autogenous control. Rna 2003, 9, 1188–1197. [Google Scholar] [CrossRef] [Green Version]
- Gong, F.; Yanofsky, C. Instruction of translating ribosome by nascent peptide. Science 2002, 297, 1864–1867. [Google Scholar] [CrossRef]
- Hao, H.; Yuan, Z.; Shen, Z.; Han, J.; Sahin, O.; Liu, P.; Zhang, Q. Mutational and transcriptomic changes involved in the development of macrolide resistance in Campylobacter jejuni. Antimicrob. Agents Chemother. 2013, 57, 1369–1378. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Wang, Y.; Zhang, Q.; Zhang, M.; Deng, F.; Shen, Z.; Wu, C.; Wang, S.; Zhang, J.; Shen, J. Report of ribosomal RNA methylase gene erm(B) in multidrug-resistant Campylobacter coli. J. Antimicrob. Chemother. 2014, 69, 964–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclercq, R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Lin, J.; Reynolds, D.L.; Zhang, Q. Contribution of the multidrug efflux transporter CmeABC to antibiotic resistance in different Campylobacter species. Foodborne Pathog. Dis. 2010, 7, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibreel, A.; Wetsch, N.M.; Taylor, D.E. Contribution of the CmeABC efflux pump to macrolide and tetracycline resistance in Campylobacter jejuni. Antimicrob. Agents Chemother. 2007, 51, 3212–3216. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Shen, Z.; Wang, Y.; Deng, F.; Liu, D.; Naren, G.; Dai, L.; Su, C.C.; Wang, B.; Wang, S.; et al. Emergence of a potent multidrug efflux pump variant that enhances Campylobacter resistance to multiple antibiotics. MBio 2016, 7, e01543-16. [Google Scholar] [CrossRef] [Green Version]
- Pumbwe, L.; Piddock, L.J.V. Identification and molecular characterisation of CmeB, a Campylobacter jejuni multidrug efflux pump. FEMS Microbiol. Lett. 2002, 206, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Boto, D.; López-Portolés, J.A.; Simón, C.; Valdezate, S.; Echeita, M.A. Study of the molecular mechanisms involved in high-level macrolide resistance of Spanish Campylobacter jejuni and Campylobacter coli strains. J. Antimicrob. Chemother. 2010, 65, 2083–2088. [Google Scholar] [CrossRef] [Green Version]
- Cagliero, C.; Maurel, M.-C.; Cloeckaert, A.; Payot, S. Regulation of the expression of the CmeABC efflux pump in Campylobacter jejuni: Identification of a point mutation abolishing the binding of the CmeR repressor in an in vitro -selected multidrug-resistant mutant. FEMS Microbiol. Lett. 2007, 267, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Cagliero, C.; Mouline, C.; Payot, S.; Cloeckaert, A. Involvement of the CmeABC efflux pump in the macrolide resistance of Campylobacter coli. J. Antimicrob. Chemother. 2005, 56, 948–950. [Google Scholar] [CrossRef] [Green Version]
- Connell, S.R.; Trieber, C.A.; Dinos, G.P.; Einfeldt, E.; Taylor, D.E.; Nierhaus, K.H. Mechanism of Tet(O)-mediated tetracycline resistance. EMBO J. 2003, 22, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C. Tetracycline Therapy: Update. Clin. Infect. Dis. 2003, 36, 462–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef]
- Warburton, P.J.; Amodeo, N.; Roberts, A.P. Mosaic tetracycline resistance genes encoding ribosomal protection proteins. J. Antimicrob. Chemother. 2016, 71, 3333–3339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibreel, A.; Tracz, D.M.; Nonaka, L.; Ngo, T.M.; Connell, S.R.; Taylor, D.E. Incidence of antibiotic resistance in Campylobacter jejuni isolated in Alberta, Canada, from 1999 to 2002, with special reference to tet(O)-mediated tetracycline resistance. Antimicrob. Agents Chemother. 2004, 48, 3442–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrob. Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Friis, L.M.; Pin, C.; Taylor, D.E.; Pearson, B.M.; Wells, J.M. A role for the tet(O) plasmid in maintaining Campylobacter plasticity. Plasmid 2007, 57, 18–28. [Google Scholar] [CrossRef]
- Kemper, N. Veterinary antibiotics in the aquatic and terrestrial environment. Ecol. Indic. 2008, 8, 1–13. [Google Scholar] [CrossRef]
- Crespo, M.D.; Altermann, E.; Olson, J.; Miller, W.G.; Chandrashekhar, K.; Kathariou, S. Novel plasmid conferring kanamycin and tetracycline resistance in the turkey-derived Campylobacter jejuni strain 11601MD. Plasmid 2016, 86, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Stanton, T.B.; McDowall, J.S.; Rasmussen, M.A. Diverse tetracycline resistance genotypes of Megasphaera elsdenii strains selectively cultured from swine feces. Appl. Environ. Microbiol. 2004, 70, 3754–3757. [Google Scholar] [CrossRef] [Green Version]
- Patterson, A.J.; Rincon, M.T.; Flint, H.J.; Scott, K.P. Mosaic tetracycline resistance genes are widespread in human and animal fecal samples. Antimicrob. Agents Chemother. 2007, 51, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- Melville, C.M.; Scott, K.P.; Mercer, D.K.; Flint, H.J. Novel tetracycline resistance gene, tet(32), in the Clostridium-related human colonic anaerobe K10 and its transmission in vitro to the rumen anaerobe Butyrivibrio fibrisolvens. Antimicrob. Agents Chemother. 2001, 45, 3246–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novais, C.; Freitas, A.R.; Silveira, E.; Baquero, F.; Peixe, L.; Roberts, A.P.; Coque, T.M. A tet(S/M) hybrid from CTn6000 and CTn916 recombination. Microbiology. 2012, 158, 2710–2711. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotra, L.P.; Haddad, J.; Mobashery, S. Aminoglycosides: Perspectives on mechanisms of action and resistance and strategies to counter resistance. Antimicrob. Agents Chemother. 2000, 44, 3249–3256. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Clowes, R.C.; Cohen, S.N.; Curtiss, R.I.; Datta, N.; Falkow, S. Uniform nomenclature for bacterial plasmids: A proposal. Bacteriol. Rev. 1976, 40, 168–189. [Google Scholar] [CrossRef] [Green Version]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef]
- Vanhoof, R.; Hannecart-Pokorni, E.; Content, J. Nomenclature of genes encoding aminoglycoside-modifying enzymes. Antimicrob. Agents Chemother. 1998, 42, 483. [Google Scholar] [CrossRef] [Green Version]
- Abril, C.; Brodard, I.; Perreten, V. Two novel antibiotic resistance genes, tet(44) and ant(6)-Ib, are located within a transferable pathogenicity island in Campylobacter fetus subsp. fetus. Antimicrob. Agents Chemother. 2010, 54, 3052–3055. [Google Scholar] [CrossRef] [Green Version]
- Hormeño, L.; Ugarte-Ruiz, M.; Palomo, G.; Borge, C.; Florez-Cuadrado, D.; Vadillo, S.; Píriz, S.; Domínguez, L.; Campos, M.J.; Quesada, A. ant(6)-I genes encoding aminoglycoside O-nucleotidyltransferases are widely spread among streptomycin resistant strains of Campylobacter jejuni and Campylobacter coli. Front. Microbiol. 2018, 9, 2515. [Google Scholar] [CrossRef] [Green Version]
- Werner, G.; Hildebrandt, B.; Witte, W. Linkage of erm(B) and aadE-sat4-aphA-3 in multiple-resistant Enterococcus faecium isolates of different ecological origins. Microb. Drug Resist. 2003, 9 (Suppl. 1), S9–S16. [Google Scholar] [CrossRef] [PubMed]
- Nirdnoy, W.; Mason, C.J.; Guerry, P. Mosaic structure of a multiple-drug-resistant, conjugative plasmid from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2454–2459. [Google Scholar] [CrossRef] [Green Version]
- Lucey, B.; Cryan, B.; O’Halloran, F.; Wall, P.G.; Buckley, T.; Fanning, S. Trends in antimicrobial susceptibility among isolates of Campylobacter species in Ireland and the emergence of resistance to ciprofloxacin. Vet. Rec. 2002, 151, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Bolton, D.; Patriarchi, A.; Fox, Á.; Fanning, S. A study of the molecular basis of quinolone and macrolide resistance in a selection of Campylobacter isolates from intensive poultry flocks. Food Control. 2013, 30, 222–226. [Google Scholar] [CrossRef]
- Lucey, B.; Crowley, D.; Moloney, P.; Cryan, B.; Daly, M.; O’ Halloran, F.; Threlfall, E.J.; Fanning, S. Integronlike structures in Campylobacter spp. of human and animal origin. Emerg. Infect. Dis. 2000, 6, 50–55. [Google Scholar]
- Corcoran, D.; Quinn, T.; Cotter, L.; O’Halloran, F.; Fanning, S. Characterization of a cmeABC operon in a quinolone-resistant Campylobacter coli isolate of Irish origin. Microb. Drug Resist. 2005, 11, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Oza, A.N.; McKenna, J.P.; McDowell, S.W.J.; Menzies, F.D.; Neill, S.D. Antimicrobial susceptibility of Campylobacter spp. isolated from broiler chickens in Northern Ireland. J. Antimicrob. Chemother. 2003, 52, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.E.; Mclernon, P.; Wareing, D.; Xu, J.; Murphy, P.G.; Mclernon, P.; Hospital, R.P.; Lane, S.G.; Pr, P. Characterisation of resistant Campylobacter species isolated from human beings and chickens. Vet. Rec. 2002, 150, 518–520. [Google Scholar] [CrossRef]
- Redondo, N.; Carroll, A.; Mcnamara, E. Molecular characterization of Campylobacter causing human clinical infection using whole-genome sequencing: Virulence, antimicrobial resistance and phylogeny in Ireland. PLoS ONE 2019, 14, e0219088. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, D.; Quinn, T.; Cotter, L.; Fanning, S. An investigation of the molecular mechanisms contributing to high-level erythromycin resistance in Campylobacter. Int. J. Antimicrob. Agents 2006, 27, 40–45. [Google Scholar] [CrossRef]
- Acke, E.; McGill, K.; Quinn, T.; Jones, B.R.; Fanning, S.; Whyte, P. Antimicrobial resistance profiles and mechanisms of resistance in Campylobacter jejuni isolates from pets. Foodborne Pathog. Dis. 2009, 6, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Minihan, D.; Whyte, P.; O’Mahony, M.; Cowley, D.; O’Halloran, F.; Corcoran, D.; Fanning, S.; Collins, J.D. Phenotypic and genotypic anti-microbial resistance profiles of Campylobacters from untreated feedlot cattle and their environment. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2006, 53, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kang, M. Molecular basis of macrolide resistance in Campylobacter strains isolated from poultry in South Korea. Biomed. Res. Int. 2018, 2018, 4526576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olkkola, S.; Culebro, A.; Juntunen, P.; Hänninen, M.L.; Rossi, M.M. Functional genomics in Campylobacter coli identified a novel streptomycin resistance gene located in a hypervariable genomic region. Microbiology 2016, 162, 1157–1166. [Google Scholar] [CrossRef]
- Fabre, A.; Oleastro, M.; Nunes, A.; Santos, A.; Ducournau, A.; Floch, P.; Lehours, P. Whole-genome sequence analysis of multidrug-resistant Campylobacter isolates: A focus on aminoglycoside resistance. J. Clin. Microbiol. 2018, 56, e00390-e18. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.T.; Lynch, H.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; Lucey, B. Antimicrobial resistance of Campylobacter isolates recovered from broilers in the Republic of Ireland in 2017 and 2018: An update. Br. Poult. Sci. 2020. [Google Scholar] [CrossRef]
- Moore, J.E.; Corcoran, D.; Dooley, J.S.G.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Megraud, F.; Millar, C.B.; O’ Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Zhang, M.; Zhou, J.; Pang, L.; Wang, G.; Hou, F. The molecular mechanisms of ciprofloxacin resistance in clinical Campylobacter jejuni and their genotyping characteristics in Beijing, China. Foodborne Pathog. Dis. 2017, 14, 386–392. [Google Scholar] [CrossRef]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Kielsznia, A.; Korzekwa, K.; Tobiasz, A.; Korzeniowska-Kowal, A.; Wieliczko, A. High prevalence of resistance to fluoroquinolones and tetracycline Campylobacter spp. isolated from poultry in Poland. Microb. Drug Resist. 2018, 24, 314–322. [Google Scholar] [CrossRef]
- Sierra-Arguello, Y.M.; Perdoncini, G.; Morgan, R.B.; Salle, C.T.P.; Moraes, H.L.S.; Gomes, M.J.P.; do Nascimento, V.P. Fluoroquinolone and macrolide resistance in Campylobacter jejuni isolated from broiler slaughterhouses in southern Brazil. Avian Pathol. 2016, 45, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Duarte, A.; Dierick, K.; Botteldoorn, N. Genetic basis and clonal population structure of antibiotic resistance in Campylobacter jejuni isolated from broiler carcasses in Belgium. Front. Microbiol. 2018, 9, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, N.; Pereira, S.; Sahin, O.; Lin, J.; Huang, S.; Michel, L.; Zhang, Q. Enhanced in vivo fitness of fluoroquinolone-resistant Campylobacter jejuni in the absence of antibiotic selection pressure. Proc. Natl. Acad. Sci. USA 2005, 102, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lin, J.; Pereira, S. Fluoroquinolone-resistant Campylobacter in animal reservoirs: Dynamics of development, resistance mechanisms and ecological fitness. Anim. Heal. Res. Rev. 2003, 4, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Whelan, M.X.; Ardill, L.; Koide, K.; Nakajima, C.; Suzuki, Y.; Simpson, J.C.; Ó Cróinín, T. Acquisition of fluoroquinolone resistance in Campylobacter jejuni leads to an increase in biofilm formation and virulence. Sci. Rep. 2019, 9, 18216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucey, B.; Feurer, C.; Greer, P.; Moloney, P.; Cryan, B.; Fanning, S. Antimicrobial resistance profiling and DNA Amplification Fingerprinting (DAF) of thermophilic Campylobacter spp. in human, poultry and porcine samples from the Cork region of Ireland. J. Appl. Microbiol. 2000, 89, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Health Products Regulatory Authority. Report on Consumption of Veterinary Antibiotics in Ireland during 2017. Available online: http://www.hpra.ie/docs/default-source/default-document-library/draft-report-on-consumption-of-veterinary-antibiotics-in-ireland-during-2017.pdf?sfvrsn=0 (accessed on 1 April 2020).
- Health Products Regulatory Authority. Report on consumption of veterinary antibiotics in Ireland during 2014. 2014. Available online: https://www.hpra.ie/docs/default-source/publications-forms/newsletters/report-on-consumption-of-veterinary-antibiotics-in-ireland-during-2014.pdf?sfvrsn=9 (accessed on 1 April 2020).
- Health Products Regulatory Authority. Report on Consumption of Veterinary Antibiotics in Ireland During 2018. Available online: https://www.hpra.ie/docs/default-source/default-document-library/report_consumption_vet_antimicrobials_2011.pdf?sfvrsn=0 (accessed on 1 April 2020).
- Ekkapobyotin, C.; Padungtod, P.; Chuanchuen, R. Antimicrobial resistance of Campylobacter coli isolates from swine. Int. J. Food Microbiol. 2008, 128, 525–528. [Google Scholar] [CrossRef]
- Kinana, A.D.; Cardinale, E.; Bahsoun, I.; Tall, F.; Sire, J.M.; Garin, B.; Boye, C.-B.; Dromigny, J.-A.; Perrier-Gros-Claude, J.-D. Analysis of topoisomerase mutations in fluoroquinolone-resistant and -susceptible Campylobacter jejuni strains isolated in Senegal. Int. J. Antimicrob. Agents 2007, 29, 397–401. [Google Scholar] [CrossRef]
- Dionisi, A.M.; Luzzi, I.; Carattoli, A. Identification of ciprofloxacin-resistant Campylobacter jejuni and analysis of the gyrA gene by the LightCycler mutation assay. Mol. Cell. Probes 2004, 18, 255–261. [Google Scholar] [CrossRef]
- Ragimbeau, C.; Colin, S.; Devaux, A.; Decruyenaere, F.; Cauchie, H.M.; Losch, S.; Penny, C.; Mossong, J. Investigating the host specificity of Campylobacter jejuni and Campylobacter coli by sequencing gyrase subunit A. BMC Microbiol. 2014, 14, 205. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.; Moreno, A.; Jimenez de Anta, M.T.; Vila, J. A double mutation in the gyrA gene is necessary to produce high levels of resistance to moxifloxacin in Campylobacter spp. clinical isolates. Int. J. Antimicrob. Agents 2005, 25, 542–545. [Google Scholar] [CrossRef]
- Bachoual, R.; Ouabdesselam, S.; Mory, F.; Lascols, C.; Soussy, C.-J.; Tankovic, J. Single or double mutational alterations of GyrA associated with fluoroquinolone resistance in Campylobacter jejuni and Campylobacter coli. Microb. Drug Resist. 2001, 7, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Tankovic, J.; Bachoual, R.; Ouabdesselam, S.; Boudjadja, A.; Soussy, C.-J. In-vitro activity of moxifloxacin against fluoroquinolone-resistant strains of aerobic Gram-negative bacilli and Enterococcus faecalis. J. Antimicrob. Chemother. 1999, 43, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piddock, L.J.V.; Ricci, V.; Pumbwe, L.; Everett, M.J.; Griggs, D.J. Fluoroquinolone resistance in Campylobacter species from man and animals: Detection of mutations in topoisomerase genes. J. Antimicrob. Chemother. 2003, 51, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakanen, A.; Jalava, J.; Kotilainen, P.; Jousimies-Somer, H.; Siitonen, A.; Huovinen, P. gyrA polymorphism in Campylobacter jejuni: Detection of gyrA mutations in 162 C. jejuni isolates by single-strand conformation polymorphism and DNA sequencing. Antimicrob. Agents Chemother. 2002, 46, 2644–2647. [Google Scholar] [CrossRef] [Green Version]
- Ge, B.; White, D.G.; Mcdermott, P.F.; Girard, W.; Zhao, S.; Hubert, S.; Meng, J. Antimicrobial-Resistant Campylobacter Species from Retail Raw Meats. Appl. Environ. Microbiol. 2003, 69, 3005–3007. [Google Scholar] [CrossRef] [Green Version]
- Thomrongsuwannakij, T.; Blackall, P.J.; Chansiripornchai, N. A study on Campylobacter jejuni and Campylobacter coli through commercial broiler production chains in Thailand. Avian Dis. 2017, 61, 186–197. [Google Scholar] [CrossRef]
- Oishi, A.; Murakami, K.; Etoh, Y.; Sera, N.; Horikawa, K. Antimicrobial susceptibility and resistance mutations in Campylobacter jejuni and C. coli isolates from human and meat sources. Kansenshogaku Zasshi. 2015, 89, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Chittum, H.S.; Champney, W.S. Ribosomal protein gene sequence changes in erythromycin-resistant mutants of Escherichia coli. J. Bacteriol. 1994, 176, 6192–6198. [Google Scholar] [CrossRef] [Green Version]
- Florez-Cuadrado, D.; Ugarte-Ruiz, M.; Quesada, A.; Palomo, G.; Domínguez, L.; Porrero, M.C. Description of an erm(B)-carrying Campylobacter coli isolate in Europe. J. Antimicrob. Chemother. 2016, 71, 841–843. [Google Scholar] [CrossRef] [Green Version]
- Florez-Cuadrado, D.; Ugarte-Ruiz, M.; Meric, G.; Quesada, A.; Porrero, M.C.; Pascoe, B.; Sáez-Llorente, J.L.; Orozco, G.L.; Domínguez, L.; Sheppard, S.K. Genome comparison of erythromycin resistant campylobacter from Turkeys identifies hosts and pathways for horizontal spread of erm(B) genes. Front. Microbiol. 2017, 8, 2240. [Google Scholar] [CrossRef] [PubMed]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Dierick, K.; Botteldoorn, N. Molecular epidemiology and antimicrobial resistance mechanisms of Campylobacter coli from diarrhoeal patients and broiler carcasses in Belgium. Transbound. Emerg. Dis. 2019, 66, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.; Tagg, K.A.; Joung, Y.J.; Bennett, C.; Watkins, L.F.; Eikmeier, D.; Folster, J.P. Report of erm(B) Campylobacter jejuni in the United States. Antimicrob. Agents Chemother. 2018, 62, e02615-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwaran, A.; Okoh, A.L. Campylobacteriosis agents in meat carcasses collected from two district municipalities in the Eastern cape province, South Africa. Foods 2020, 9, 203. [Google Scholar] [CrossRef] [Green Version]
- Luangtongkum, T.; Shen, Z.; Seng, V.W.; Sahin, O.; Jeon, B.; Liu, P.; Zhang, Q. Impaired fitness and transmission of macrolide-resistant Campylobacter jejuni in its natural host. Antimicrob. Agents Chemother. 2012, 56, 1300–1308. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Pu, S.; Wang, F.; Meng, J.; Ge, B. Fitness cost of macrolide resistance in Campylobacter jejuni. Int. J. Antimicrob. Agents 2009, 34, 462–466. [Google Scholar] [CrossRef]
- Zeitouni, S.; Collin, O.; Andraud, M.; Ermel, G.; Kempf, I. Fitness of macrolide resistant Campylobacter coli and Campylobacter jejuni. Microb. Drug Resist. 2012, 18, 101–108. [Google Scholar] [CrossRef]
- Stanton, T.B.; Humphrey, S.B. Isolation of tetracycline-resistant Megasphaera elsdenii strains with novel mosaic gene combinations of tet(O) and tet(W) from swine. Appl. Environ. Microbiol. 2003, 69, 3874–3882. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I.; Garrigues-Jeanjean, N.; Mackie, R.I. Molecular ecology of tetracycline resistance: Development and validation of primers for detection of tetracycline resistance genes encoding ribosomal protection proteins. Appl. Environ. Microbiol. 2001, 67, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, N.P.; Kuijf, M.L.; Ang, C.W.; Schiellerup, P.; Krogfelt, K.A.; Jacobs, B.C.; van Belkum, A.; Endtz, H.P.; Bergman, M.P. Sialylation of Campylobacter jejuni lipo-oligosaccharides is associated with severe gastro-enteritis and reactive arthritis. Microbes Infect. 2009, 11, 988–994. [Google Scholar] [CrossRef]
- Dearlove, B.L.; Cody, A.J.; Pascoe, B.; Méric, G.; Wilson, D.J.; Sheppard, S.K. Rapid host switching in generalist Campylobacter strains erodes the signal for tracing human infections. ISME J. 2016, 10, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llarena, A.K.; Zhang, J.; Vehkala, M.; Välimäki, N.; Hakkinen, M.; Hänninen, M.L.; Roasto, M.; Mäesaar, M.; Taboada, E.; Barker, D.; et al. Monomorphic genotypes within a generalist lineage of Campylobacter jejuni show signs of global dispersion. Microb. Genom. 2016, 2, e000088. [Google Scholar] [CrossRef] [PubMed]
- Mohan, V.; Stevenson, M.; Marshall, J.; Fearnhead, P.; Holland, B.R.; Hotter, G.; French, N.P. Campylobacter jejuni colonization and population structure in urban populations of ducks and starlings in New Zealand. Microbiologyopen 2013, 2, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kang, M.; Jang, H.K. Genetic characterization and epidemiological implications of Campylobacter isolates from wild birds in South Korea. Transbound. Emerg. Dis. 2019, 66, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäesaar, M.; Meremäe, K.; Ivanova, M.; Roasto, M. Antimicrobial resistance and multilocus sequence types of Campylobacter jejuni isolated from Baltic broiler chicken meat and Estonian human patients. Poult. Sci. 2018, 97, 3645–3651. [Google Scholar] [CrossRef] [PubMed]
- Aksomaitiene, J.; Ramonaite, S.; Tamuleviciene, E.; Novoslavskij, A.; Alter, T.; Malakauskas, M. Overlap of antibiotic resistant Campylobacter jejuni MLST genotypes isolated from humans, broiler products, dairy cattle and wild birds in Lithuania. Front. Microbiol. 2019, 10, 1377. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Deng, F.; Shen, Z.; Wu, C.; Zhang, J.; Zhang, Q.; Shen, J. Emergence of multidrug-resistant Campylobacter species isolates with a horizontally acquired rRNA methylase. Antimicrob. Agents Chemother. 2014, 58, 5405–5412. [Google Scholar] [CrossRef] [Green Version]
- Burckhardt, R.M.; Escalante-Semerena, J.C. Insights into the function of the N-acetyltransferase SatA that detoxifies streptothricin in Bacillus subtilis and Bacillus anthracis. Appl. Environ. Microbiol. 2019, 85, e03029-18. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Wang, Y.; Zhang, Q.; Chen, X.; Shen, Z.; Deng, F.; Wu, C.; Shen, J. Identification of a novel genomic island conferring resistance to multiple aminoglycoside antibiotics in Campylobacter coli. Antimicrob. Agents Chemother. 2012, 56, 5332–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PubMLST. Available online: https://pubmlst.org/ (accessed on 24 February 2020).
- Wright, S.; Wilson, S.; Miller, W.G.; Mandrell, R.E.; Siletzky, R.M.; Kathariou, S. Differences in methylation at GATC sites in genomic DNA of Campylobacter coli from turkeys and swine. Appl. Environ. Microbiol. 2010, 76, 7314–7317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadi, V.A.; Matthew-Belmar, V.; Subbarao, C.; Kashoma, I.; Rajashekara, G.; Sharma, R.; Hariharan, H.; Stone, D. Campylobacter species isolated from pigs in Grenada exhibited novel clones: Genotypes and antimicrobial resistance profiles of sequence types. Foodborne Pathog. Dis. 2017, 14, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cody, A.J.; McCarthy, N.D.; Van Rensburg, M.J.; Isinkaye, T.; Bentley, S.D.; Parkhill, J.; Dingle, K.E.; Bowler, I.C.J.W.; Jolley, K.A.; Maiden, M.C.J. Real-time genomic epidemiological evaluation of human Campylobacter isolates by use of whole-genome multilocus sequence typing. J. Clin. Microbiol. 2013, 51, 2526–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control. EU Protocol for Harmonised Monitoring of Antimicrobial Resistance in Human Salmonella and Campylobacter Isolates. Available online: https://www.ecdc.europa.eu/sites/default/files/media/en/publications/Publications/antimicrobial-resistance-Salmonella-Campylobacter-harmonised-monitoring.pdf (accessed on 7 June 2016).
- European Commission. Commission Implementing Decision of 12 November 2013 on the monitoring and reporting of antimicrobial resistance in zoonotic and commensal bacteria. Off. J. Eur. Union 2013, L 303, 26–39. [Google Scholar]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacher, S.; Ménard, A.; Bernard, E.; Mégraud, F. PCR-restriction fragment length polymorphism analysis for detection of point mutations associated with macrolide resistance in Campylobacter spp. Antimicrob. Agents Chemother. 2003, 47, 1125–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godschalk, P.C.R.; Kuijf, M.L.; Li, J.; Michael, F.S.; Ang, C.W.; Jacobs, B.C.; Karwaski, M.-F.; Brochu, D.; Moterassed, A.; Endtz, H.P.; et al. Structural characterization of Campylobacter jejuni lipooligosaccharide outer cores associated with Guillain-Barré and Miller Fisher Syndromes. Infect. Immun. 2007, 75, 1245–1254. [Google Scholar] [CrossRef] [Green Version]
- Guyard-Nicodème, M.; Rivoal, K.; Houard, E.; Rose, V.; Quesne, S.; Mourand, G.; Rouxel, S.; Kempf, I.; Guillier, L.; Gauchard, F.; et al. Prevalence and characterization of Campylobacter jejuni from chicken meat sold in French retail outlets. Int. J. Food Microbiol. 2015, 203, 8–14. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Genome analysis Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Genome analysis QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| GTJ | n= (%) | Nucleotide Position (Base Indicated in Brackets) of Wild-type Strain C. jejuni NCTC 11168 gyrA (GenBank Accession Number: L04566.1 and AL111168.1) | ||||||||||
| 64 (A) | 72 (T) | 243 (C) | 257 (C) | 357 (T) | 360 (C) | 471 (C) | 483 (T) | 608 (A) | 616 (G) | 622 (T) | ||
| GTJ-I | 40 (47.1%) | . | . | . | T | . | . | . | . | . | . | . |
| GTJ-II | 30 (35.3%) | G | C | T | T | C | T | T | C | G | . | T |
| GTJ-III | 15 (17.7%) | . | C | T | T | C | T | T | C | G | A | T |
| GTC | n= (%) | Nucleotide Position (Base Indicated in Brackets) of Wild-type Strain C. coli NCTC 11366 gyrA (GenBank Accession Number: AF092101.1 and NZ_UIGM01000003.1) | ||||||||||
| 117 (T) | 252 (C) | 257 (C) | 297 (C) | 342 (T) | 471 (T) | 498 (G) | ||||||
| GTC-I | 1 (7.1) | . | . | T | C | . | T | . | ||||
| GTC-II | 7 (50) | C | . | T | C | . | T | . | ||||
| GTC-III | 1 (7.1) | C | . | T | . | . | T | . | ||||
| GTC-IV | 1 (7.1) | . | . | T | C | . | . | . | ||||
| GTC-V | 2 (14.3) | . | T | T | C | . | T | . | ||||
| GTC-VI | 1 (7.1) | . | . | T | C | . | . | A | ||||
| GTC-VII | 1 (7.1) | . | . | T | C | C | T | . | ||||
| Species/Strain | C. jejuni CITCj625-18 tetO/32/O | C. jejuni CITC727-18 tetO/32/O | C. coli CITCc-3448-18 tetO/32/O | KY994102.1 S. suistetO/32/O |
|---|---|---|---|---|
| C. jejuni CITCj625-18 tetO/32/O | 100% (100%) | 100% (100%) | 99.78% (96%) | 99.73% (96%) |
| C. jejuni CITC727-18 tetO/32/O | 100% (100%) | 100% (100%) | 99.78% (96%) | 99.73% (96%) |
| C. coli CITCc-3448-18 tetO/32/O | 99.78% (100%) | 99.78% (100%) | 100% (100%) | 99.95% (100%) |
| KY994102.1 S. suis tetO/32/O | 99.73% (100%) | 99.73% (100%) | 99.95% (96%) | 100% (100%) |
| Locus Tag | Gene Annotation | InterProScan Protein Family or Domain | GenBank Accession Number of Closest (% Identity/% Coverage) | Predicted Function |
|---|---|---|---|---|
| HBF06_00624 | Holliday junction ATP-dependent DNA helicase, RuvB | IPR004605 | WP_002856258.1 100/100 | Holliday junction helicase (strand exchange reactions in homologous recombination) |
| HBF06_00625 | Histidine phosphotransferase | IPR036641 | MPA99107.1 100/90) | Signal transduction |
| HBF06_00627 | Autotransporter adhesion, CapC | IPR005546 | EAK6247206.1 99.16/94 | Protein secretion |
| HBF06_00628 | Truncated tetracycline resistance ribosomal protection protein, TetO | IPR035650 | AUA17601.1 99.59/82 | Ribosomal protection protein conferring tetracycline resistance (TetO) |
| HBF06_00629 | Aminoglycoside 6- nucleotidyltransferase, ANT(6)-Ib | IPR007530 | WP_001255868.1 100/100 | Adenylyltransferase activity conferring resistance to aminoglycosides |
| HBF06_00630 | Replication protein, RepB | IPR002631 | WP_052777339.1 100/100 | Plasmid replication protein |
| HBF06_00631 | DNA topoisomerase | IPR000380 | WP_139898553.1 99.52/100 | Topoisomerisation and single stranded breakage during transcription, DNA replication and recombination. |
| HBF06_00632 | Streptothricin acetyltransferase, SatA | IPR008125 | EOO12820.1 96.86/100 | Acetylation of streptothricin conferring resistance |
| HBF06_00633 | Mosaic tetracycline resistance ribosomal protection protein, TetO/32/O | IPR035650 | WP_052855148.1 100/100 | Ribosomal protection protein conferring tetracycline resistance |
| HBF06_00634 | Plasmid replication protein | None detected | EDP4862066.1 99.21/97 | Plasmid replication |
| Primer | Type | Sequence (5’ - 3’) | Target | Amplicon Size (bp) | Annealing (°C ) | Reference |
|---|---|---|---|---|---|---|
| GZgyrA5 | A + S | ATT TTT AGC AAA GAT TCT GAT | QRDR region of gyrA | 673 | 50 | [15] |
| GZgyrA6 | CCA TAA ATT ATT CCA CCT GT | |||||
| TetO-FW | A | ACG GAR AGT TTA TTG TAT ACC | tetO | 171 | 52 | [103] |
| TetO-RV | TGG CGT ATC TAT AAT GTT GAC | |||||
| TetO/W/O-F | A | ATC CAG ACA GCA GTG ACA TC | tetO/W/O | 489 | 50 | This study |
| TetO/W/O-R | ATG ATA GAC CGG AAA CAG GG | |||||
| TetO/32/O-i-F | A | GAT ACA ATG AAT TTG GAG CG | tetO/32/O type I | 545 | 48 | This study |
| TetO/32/O-i-R | AAT TGT CTT TTG CAC TCC C | |||||
| TetO/32/O-ii-F | A | CGG GCA GGT TTT TAAG ATT G | tetO/32/Oi type II | 365 | 50 | This study |
| TetO/32/O-ii-R | CTG TAT CAG CAA TCT CTG CG | |||||
| F2-campy-23S | A + S | AAT TGA TGG GGT TAG CAT TAG C | Domain V of 23S rRNA | 316 | 55 | [121] |
| R2-campy-23S | CAA CAA TGG CTC ATA TAC AAC TTG | |||||
| L4C-F | A + S | TTA TCC CTC TTT TGT AAT AGA TTC TAA | rplD | 614 | 48 | [61] |
| L4C-R | ATG AGT AAA GTA GTT GTT TTA AAT GAT | |||||
| L22C-F | A + S | TTA GCT TTC CTT TTT CAC TGT TGC TTT | rplV | 425 | 48 | [61] |
| L22C-R | ATG AGT AAA GCA TTA ATT AAA TTC ATA AG | |||||
| erm(B)-F | A | GGG CAT TTA ACG ACG AAA CTG G | ermB | 421 | 52 | [111] |
| erm(B)-R | CTG TGG TAT GGC GGG TAA GT | |||||
| CmecoliF3 | A + S | AATGTTTTAGCCGATACT | cmeABC | 428 | 45 | [28] |
| CmecoliR4 | AACACCGCTTACTTGAGG | |||||
| cst-II-F | A | ATG AAA AAA GTT ATT ATT GCT GGA AAT G | LOS locus class A/B | 885/876 | 50 | [122,123] |
| cst-II-R | TTA TTT TCC TTT GAA ATA ATG CTT TAT | |||||
| orf14c-F | A | CAA CTT TGC AAA ATG ATT TTA TCT ATC ATT | LOS locus class C | 995 | 50 | [123] |
| orf14c-R | ATG CAA ATA CAA CAA AAC AAT TC |
| Isolate | Gene | GenBank Accession Number | Sequence Length (bp) |
|---|---|---|---|
| C. coli (CITCc1303-18) | 23S ribosomal RNA, partial sequence | MT155934 | 262 |
| C. coli (CITCc1303-18) | 50S ribosomal protein L4 (rplD) gene, partial cds | MT155935 | 519 |
| C. coli (CITCc1303-18) | 50S ribosomal protein L22 (rplV) gene, partial cds | MT155936 | 319 |
| C. jejuni( CITCj4193-17) | gyrA (GTJ-I), partial cds | MT176407 | 644 |
| C. jejuni (CITCj999-18) | gyrA (GTJ-II), partial cds | MT176408 | 644 |
| C. jejuni (CITCj193-18) | gyrA (GTJ-III), partial cds | MT176409 | 644 |
| C. coli (CITCc3796-B-18) | gyrA (GTC-I), partial cds | MT176400 | 550 |
| C. coli (CITCc3636-B-17 | gyrA (GTC-II), partial cds | MT176401 | 550 |
| C. coli (CITCc3521-18) | gyrA (GTC-III), partial cds | MT176402 | 550 |
| C. coli (CITCc3318-17) | gyrA (GTC-IV), partial cds | MT176403 | 550 |
| C. coli (CITCc1631-18) | gyrA (GTC-V), partial cds | MT176404 | 550 |
| C. coli (CITCc3790-18) | gyrA (GTC-VI), partial cds | MT176405 | 550 |
| C. coli (CITCc1303-18) | gyrA (GTC-VII), partial cds | MT176406 | 550 |
| C. jejuni (CITCj625-18) | tetO/32/O type II, complete cds | MT176410 | 1920 |
| C. jejuni (CITcj727-18) | tetO/32/O type II, complete cds | MT176411 | 1920 |
| C. coli (CITCc3448-18) | tetO/32/O type II, complete cds | MT176412 | 1920 |
| C. jejuni (CITCj625-18) | ant(6)-Ib, complete cds | MT176413 | 867 |
| C. jejuni (CITcj727-18) | ant(6)-Ib, complete cds | MT176414 | 867 |
| C. coli (CITCc3448-18) | ant(6)-Ie, complete cds | MT176415 | 900 |
| C. coli (CITCc1631-18) | ant(6)-Ie, complete cds | MT176416 | 550 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, C.T.; Lynch, H.; Burke, S.; Hawkins, K.; Buttimer, C.; Mc Carthy, C.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; et al. Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics 2020, 9, 308. https://doi.org/10.3390/antibiotics9060308
Lynch CT, Lynch H, Burke S, Hawkins K, Buttimer C, Mc Carthy C, Egan J, Whyte P, Bolton D, Coffey A, et al. Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics. 2020; 9(6):308. https://doi.org/10.3390/antibiotics9060308
Chicago/Turabian StyleLynch, Caoimhe T., Helen Lynch, Sarah Burke, Kayleigh Hawkins, Colin Buttimer, Conor Mc Carthy, John Egan, Paul Whyte, Declan Bolton, Aidan Coffey, and et al. 2020. "Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period" Antibiotics 9, no. 6: 308. https://doi.org/10.3390/antibiotics9060308
APA StyleLynch, C. T., Lynch, H., Burke, S., Hawkins, K., Buttimer, C., Mc Carthy, C., Egan, J., Whyte, P., Bolton, D., Coffey, A., & Lucey, B. (2020). Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics, 9(6), 308. https://doi.org/10.3390/antibiotics9060308