The Inhibitory Effect of Plant Extracts on Growth of the Foodborne Pathogen, Listeria monocytogenes






Abstract
:1. Introduction
2. Results
2.1. Effect of Plant Extract on Growth of L. monocytogenes and Determination of Minimum Inhibitory Concentrations (MICs)
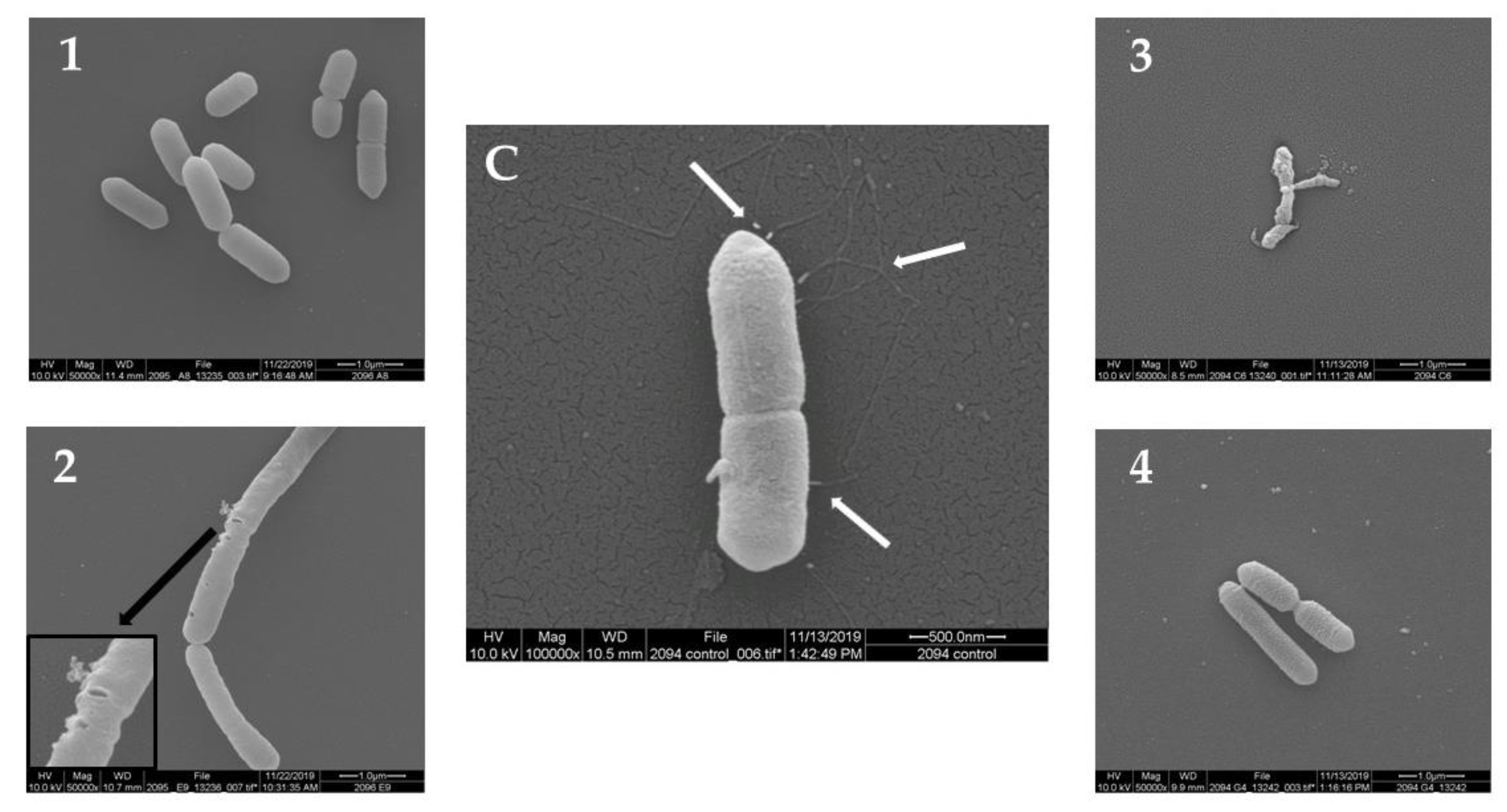
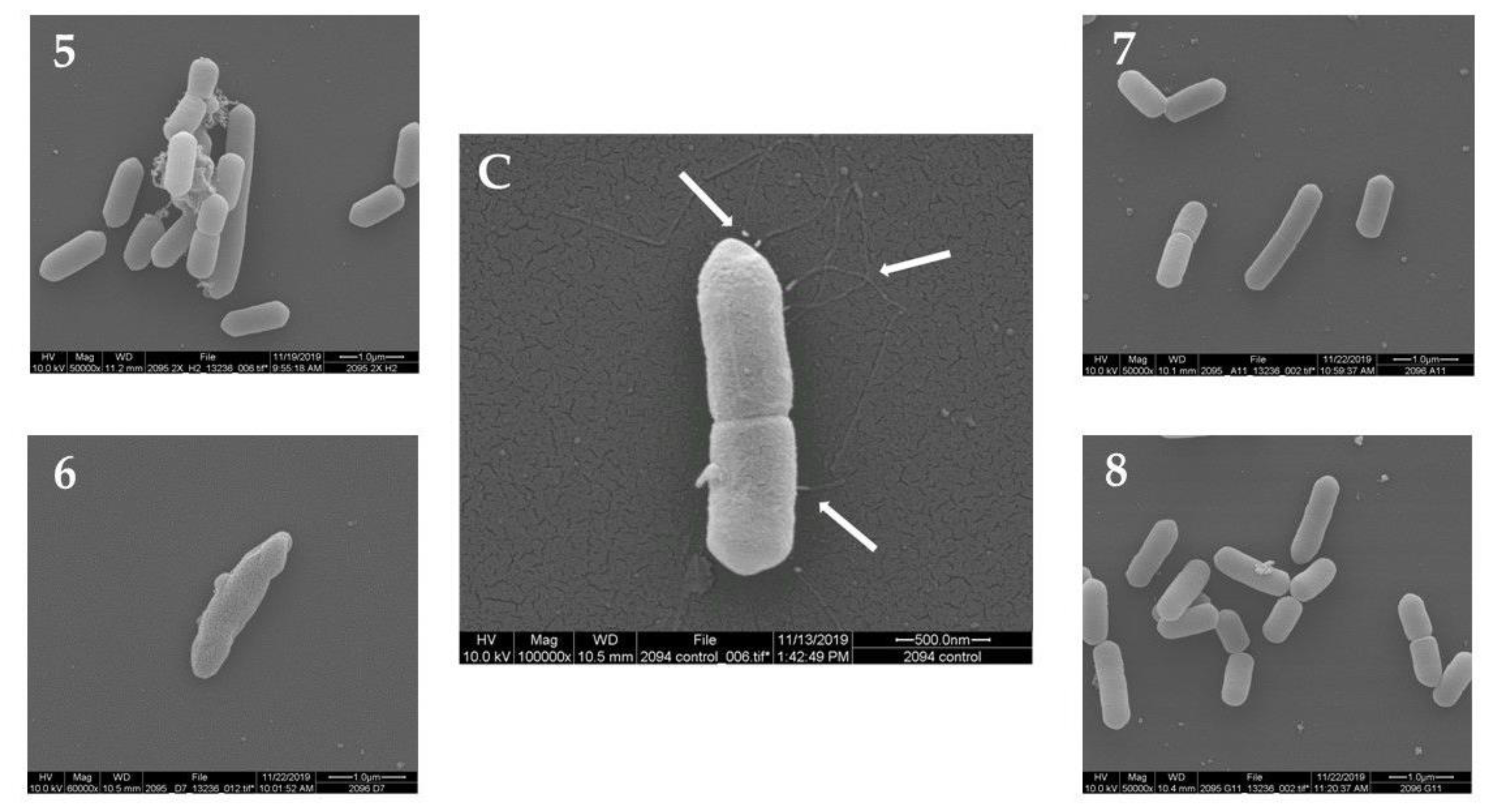
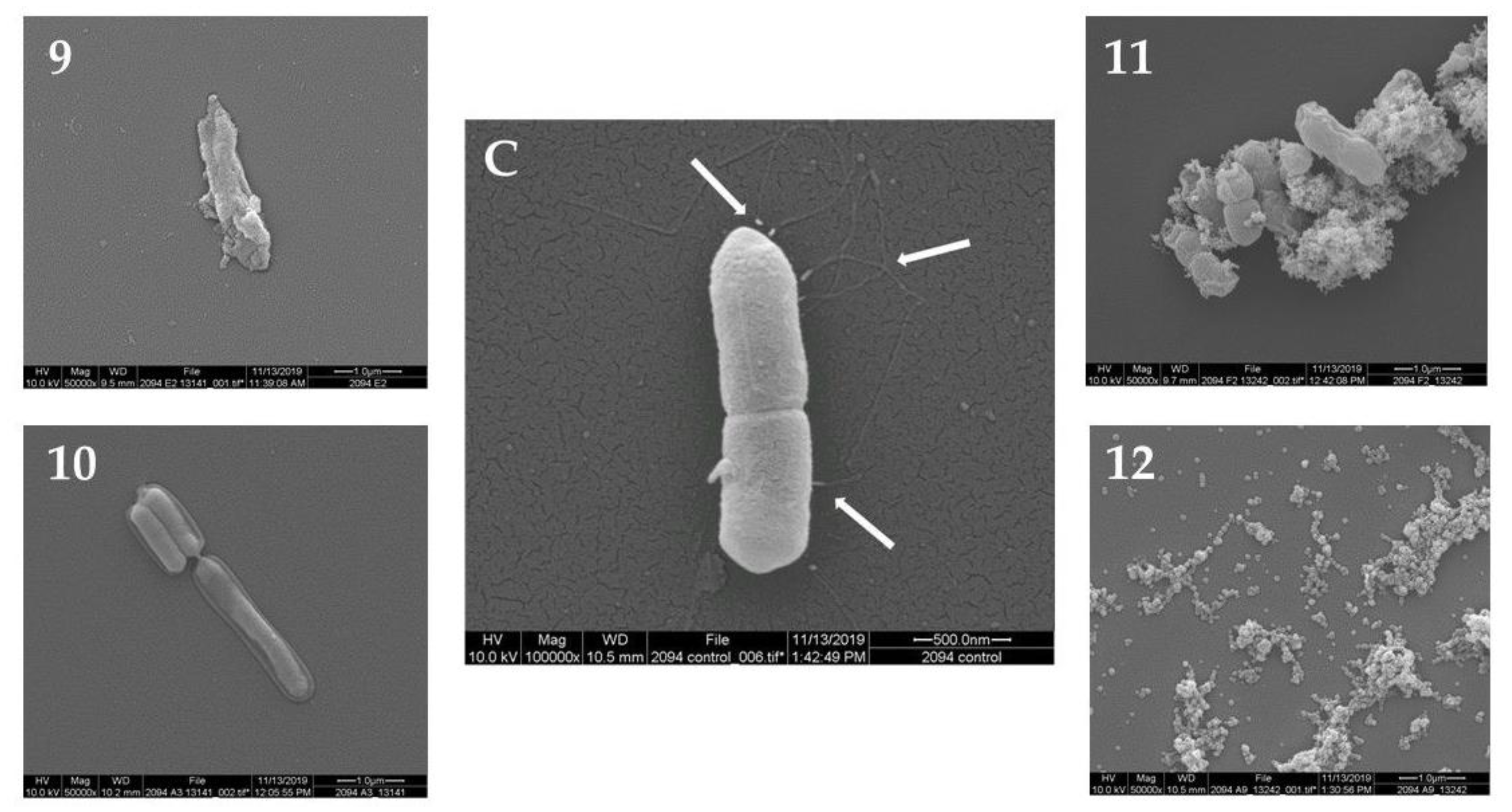
2.2. Scanning Electron Microscopy (SEM)
3. Discussion
4. Materials and Methods
4.1. Plant Extracts and Crude Fractionation
4.2. L. monocytogenes Strain and Culture Conditions
4.3. Plant Extracts Effect on L. monocytogenes Growth and Determination of MICs
4.4. Scanning Electron Microscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar] [CrossRef] [Green Version]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar] [CrossRef]
- Allen, K.; Wałecka-Zacharska, E.; Kosek-Paszkowska, K.; Devlieghere, F.; Van Meervenne, E.; Osek, J.; Wieczorek, K.; Bania, J. Listeria monocytogenes–An examination of food chain factors potentially contributing to antimicrobial resistance. Food Microbiol. 2015, 54, 178–189. [Google Scholar] [CrossRef]
- Bahrami, A.; Baboli, Z.M.; Schimmel, K.; Jafari, S.M.; Williams, L. Efficiency of novel processing technologies for the control of Listeria monocytogenes in food products. Trends Food Sci. Technol. 2020, 96, 61–78. [Google Scholar] [CrossRef]
- Rothrock, M.J., Jr.; Micciche, A.C.; Bodie, A.R.; Ricke, S.C. Listeria occurrence and potential control strategies in alternative and conventional poultry processing and retail. Front. Sustain. Food Syst. 2019, 3, 33. [Google Scholar] [CrossRef]
- Pouillot, R.; Hoelzer, K.; Jackson, K.A.; Henao, O.L.; Silk, B.J. Relative risk of listeriosis in Foodborne Diseases Active Surveillance Network (FoodNet) sites according to age, pregnancy, and ethnicity. Clin. Infect. Dis. 2012, 54, S405–S410. [Google Scholar] [CrossRef] [Green Version]
- Fei, P.; Ali, M.A.; Gong, S.; Sun, Q.; Bi, X.; Liu, S.; Guo, L. Antimicrobial activity and mechanism of action of olive oil polyphenols extract against Cronobacter sakazakii. Food Control 2018, 94, 289–294. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; Vincken, J.-P.; van Ederen, R.; den Besten, H.M.W.; Gruppen, H. Rapid membrane permeabilization of Listeria monocytogenes and Escherichia coli induced by antibacterial prenylated phenolic compounds from legumes. Food Chem. 2018, 240, 147–155. [Google Scholar] [CrossRef]
- Guo, L.; Sun, Q.; Gong, S.; Bi, X.; Jiang, W.; Xue, W.; Fei, P. Antimicrobial activity and action approach of the olive oil polyphenol extract against Listeria monocytogenes. Front. Microbiol. 2019, 10, 1586. [Google Scholar] [CrossRef] [Green Version]
- Xylia, P.; Chrysargyris, A.; Botsaris, G.; Tzortzakis, N. Mint and pomegranate extracts/oils as antibacterial agents against Escherichia coli O157:H7 and Listeria monocytogenes on shredded carrots. J. Food Safety 2018, 38, e12423. [Google Scholar] [CrossRef]
- Germanò, M.P.; D’Angelo, V.; Sanogo, R.; Catania, S.; Alma, R.; De Pasquale, R.; Bisignano, G. Hepatoprotective and antibacterial effects of extracts from Trichilia emetica Vahl. (Meliaceae). J. Ethnopharmacol. 2005, 96, 227–232. [Google Scholar] [CrossRef]
- Moyo, P.; Botha, M.E.; Nondaba, S.; Niemand, J.; Maharaj, V.J.; Eloff, J.N.; Louw, A.I.; Birkholtz, L. In vitro inhibition of Plasmodium falciparum early and late stage gametocyte viability by extracts from eight traditionally used South African plant species. J. Ethnopharmacol. 2016, 185, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, K.; Dhawan, S. Sharma, A. Passiflora: A review update. J. Ethnopharmacol. 2004, 94, 1–23. [Google Scholar] [CrossRef]
- Sasikala, V.; Saravanan, S.; Parimelazhagan, T. Analgesic and anti-inflammatory activities of Passiflora foetida L. Asian Pac. J. Trop. Med. 2011, 4, 600–603. [Google Scholar] [CrossRef] [Green Version]
- Bendini, A.; Cerretani, L.; Pizzolante, L.; Toschi, T.G.; Guzzo, F.; Ceoldo, S.; Marconi, A.M.; Andreetta, F.; Levi, M. Phenol content related to antioxidant and antimicrobial activities of Passiflora spp. extracts. Eur. Food Res. Technol. 2006, 223, 102–109. [Google Scholar] [CrossRef]
- Simão, J.M.; Barboza, T.; Vianna, M.; Garcia, R.; Mansur, E.; Ignacio, A.C.; Pacheco, G. A comparative study of phytoconstituents and antibacterial activity of in vitro derived materials of four Passiflora species. An. Acad. Bras. Cienc. 2018, 90, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Mnayer, D.; Roointan, A.; Shahri, F.; Ayatollahi, S.A.; Sharifi-Rad, M.; Molaee, N.; Sharifi-Rad, M. Antibacterial activities of essential oils from Iranian medicinal plants on extended-spectrum β-lactamase-producing Escherichia coli. Cell Mol. Biol. (Noisy-le-grand) 2016, 62, 75–82. [Google Scholar]
- Lu, Y.; Foo, L.Y. Polyphenolics of Salvia—a review. Phytochemistry 2002, 59, 117–140. [Google Scholar] [CrossRef]
- Cocan, I.; Alexa, E.; Danciu, C.; Radulov, I.; Galuscan, A.; Obistioiu, D.; Morvay, A.A.; Sumalan, R.M.; Poiana, M.A.; Pop, G.; et al. Phytochemical screening and biological activity of Lamiaceae family plant extracts. Exp. Ther. Med. 2018, 15, 1863–1870. [Google Scholar] [CrossRef] [Green Version]
- Firuzi, O.; Miri, R.; Asadollahi, M.; Eslami, S.; Jassbi, A.R. Cytotoxic, antioxidant and antimicrobial activities and phenolic contents of eleven salvia species from Iran. Iran. J. Pharm. Res. 2013, 12, 801–810. [Google Scholar]
- Jabbari, M.; Daneshfard, B.; Emtiazy, M.; Khiveh, A.; Hashempur, M.H. Biological Effects and Clinical Applications of Dwarf Elder (Sambucus ebulus L): A Review. J. Evid. Based Complementary Altern. Med. 2017, 22, 996–1001. [Google Scholar] [CrossRef] [Green Version]
- Süntar, I.P.; Akkol, E.K.; Yalçin, F.N.; Koca, U.; Keleş, H.; Yesilada, E. Wound healing potential of Sambucus ebulus L. leaves and isolation of an active component, quercetin 3-O-glucoside. J. Ethnopharmacol. 2010, 129, 106–114. [Google Scholar] [CrossRef]
- Schwaiger, S.; Zeller, I.; Pölzelbauer, P.; Frotschnig, S.; Laufer, G.; Messner, B.; Pieri, V.; Stuppner, H.; Bernhard, D. Identification and pharmacological characterization of the anti-inflammatory principal of the leaves of dwarf elder (Sambucus ebulus L.). J. Ethnopharmacol. 2011, 133, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimzadeh, M.A.; Nabavi, S.F.; Nabavi, S.M. Antioxidant activities of methanol extract of Sambucus ebulus L. flower. Pak. J. Biol. Sci. 2009, 12, 447–450. [Google Scholar] [CrossRef]
- Hosseinimehr, S.J.; Pourmorad, F.; Shahabimajd, N.; Shahrbandy, K.; Hosseinzadeh, R.; Pak, J. In vitro antioxidant activity of Polygonium hyrcanicum, Centaurea depressa, Sambucus ebulus, Mentha spicata and Phytolacca americana. Biol. Sci. 2007, 10, 637–640. [Google Scholar] [CrossRef] [Green Version]
- Salehzadeh, A.; Asadpour, L.; Naeemi, A.S.; Houshmand, E. Antimicrobial activity of methanolic extracts of Sambucus ebulus and Urtica dioica against clinical isolates of methicillin resistant Staphylococcus aureus. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 38–40. [Google Scholar] [CrossRef] [Green Version]
- Quave, C.L.; Plano, L.R.; Bennett, B.C. Quorum sensing inhibitors of Staphylococcus aureus from Italian medicinal plants. Planta Med. 2011, 77, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Bello, I.C.; Dorling, P.; Fellows, L.; Winchester, B. Specific inhibition of human beta-D-glucuronidase and alpha-L-iduronidase by a trihydroxy pipecolic acid of plant origin. FEBS Lett. 1984, 176, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Nazeer, M.A.; Khoshoo, T.N. Cytology of some species of Sansevieria Thunb. Cytologia 1984, 49, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Wyk, B.E.; Oudtshoorn, B.; Gericke, N. Medicinal Plants of South Africa, 2nd ed.; Briza Publications: Pretoria, South Africa, 2009. [Google Scholar]
- Giovannini, P.; Howes, M.R. Medicinal plants used to treat snakebite in Central America: Review and assessment of scientific evidence. J. Ethnopharmacol. 2017, 199, 240–256. [Google Scholar] [CrossRef]
- Sriprapat, W.; Boraphech, P.; Thiravetyan, P. Factors affecting xylene-contaminated air removal by the ornamental plant Zamioculcas zamiifolia. Environ. Sci. Pollut. Res. Int. 2014, 21, 2603–2610. [Google Scholar] [CrossRef]
- Sriprapat, W.; Suksabye, P.; Areephak, S.; Klantup, P.; Waraha, A.; Sawattan, A.; Thiravetyan, P. Uptake of toluene and ethylbenzene by plants: Removal of volatile indoor air contaminants. Ecotoxicol. Environ. Saf. 2014, 102, 147–151. [Google Scholar] [CrossRef]
- Amoateng, P.; Adjei, S.; Osei-Safo, D.; Kukuia, K.K.E.; Karikari, T.K.; Nyarko, A.K. An ethanolic extract of Desmodium adscendens exhibits antipsychotic-like activity in mice. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Awouafack, M.D.; McGaw, L.J.; Gottfried, S.; Mbouangouere, R.; Tane, P.; Spiteller, M.; Eloff, J.N. Antimicrobial activity and cytotoxicity of the ethanol extract, fractions and eight compounds isolated from Eriosema robustum (Fabaceae). BMC Complement. Altern. Med. 2013, 13, 289. [Google Scholar] [CrossRef] [Green Version]
- Awouafack, M.D.; Tane, P.; Spiteller, M.; Eloff, J.N. Eriosema (Fabaceae) species represent a rich source of flavonoids with interesting pharmacological activities. Nat. Prod. Commun. 2015, 10, 1325–1330. [Google Scholar] [CrossRef] [Green Version]
- Hotti, H.; Gopalacharyulu, P.; Seppänen-Laakso, T.; Rischer, H. Metabolite profiling of the carnivorous pitcher plants Darlingtonia and Sarracenia. PLoS ONE 2017, 12, e0171078. [Google Scholar] [CrossRef]
- Riffle, M.S.; Waller, G.R.; Murray, D.S.; Sgaramello, R.P. Devil’s-claw (Proboscidea louisianica), essential oil and its components: Potential allelochemical agents on cotton and wheat. J. Chem. Ecol. 1990, 16, 1927–1940. [Google Scholar] [CrossRef]
- Connor, S.E.; Kvavadze, E.V. Modelling late quaternary changes in plant distribution, vegetation and climate using pollen data from Georgia, Caucasus. J. Biogeogr. 2009, 36, 529–545. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant polyphenols as antioxidant and antibacterial agents for shelf-life extension of meat and meat products: Classification, structures, sources, and action mechanisms. Compr. Rev. Food Sci. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [Green Version]
- Okuda, T. Systematics and health effects of chemically distinct tannins in medicinal plants. Phytochemistry 2005, 66, 2012–2031. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative Structure-Activity Relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Valdramidis, V.P.; O’Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P.J. Application of natural antimicrobials for food preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef] [Green Version]
- Sultanbawa, Y. Plant antimicrobials in food applications: Minireview. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; Volume 2, pp. 1084–1099. [Google Scholar]
- Field, J.A.; Lettinga, G. Toxicity of tannic compounds to microorganisms. In Plant Polyphenols; Hemingway, R.W., Laks, P.E., Eds.; Springer US: London, UK, 1992; pp. 673–692. [Google Scholar]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. World Sci. J. 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kumar, Y.; Yadav, D.N.; Ahmad, T.; Narsaiah, K. Recent trends in the use of natural antioxidants for meat and meat products. Compr. Rev. Food Sci. Food Saf. 2015, 14, 796–812. [Google Scholar] [CrossRef] [Green Version]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta. 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Cho, Y.S.; Schiller, N.L.; Kahng, H.Y.; Oh, K.H. Cellular responses and proteomic analysis of Escherichia coli exposed to green tea polyphenols. Curr. Microbiol. 2007, 55, 501–506. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kaihatsu, K.; Nishino, K.; Ogawa, M.; Kato, N.; Yamaguchi, A. Antibacterial and antifungal activities of new acylated derivatives of epigallocatechin gallate. Front. Microbiol. 2012, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals (Basel) 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Yi, S.; Wang, W.; Bai, F.; Zhu, J.; Li, J.; Li, X.; Xu, I.; Sun, T.; He, Y. Antimicrobial effect and membrane-activ mechanism of tea polyphenols against Serratia marcescens. World J. Microbiol. Biotechnol. 2014, 30, 451–460. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Liu, Y.; McKeever, C.L.; Malik, N.S.A. Assessment of the antimicrobial activity of olive leaf extract against foodborne bacterial pathogens. Front. Microbiol. 2017, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, A.; Saavedra, M.J.; Simões, M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling 2012, 28, 755–767. [Google Scholar] [CrossRef]
- Carraro, L.; Fasolato, L.; Montemurro, F.; Martino, M.E.; Balzan, S.; Servili, M.; Novelli, E.; Cardazzo, B. Polyphenols from olive mill waste affect biofilm formation and motility in Escherichia coli K-12. Microb. Biotechnol. 2014, 7, 265–275. [Google Scholar] [CrossRef]
- Du, R.; Qu, Y.; Qi, P.X.; Sun, X.; Liu, Y.; Zhao, M. Natural flagella-templated Au nanowires as a novel adjuvant against Listeria monocytogenes. Nanoscale 2020, 12, 5627–5635. [Google Scholar] [CrossRef]
- Linnan, M.J.; Mascola, L.; Lou, X.D.; Goulet, V.; May, S.; Salminen, C.; Hird, D.W.; Yonekura, M.L.; Hayes, P.; Weaver, R.; et al. Epidemic listeriosis associated with Mexican-style cheese. N. Engl. J. Med. 1988, 319, 823–828. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
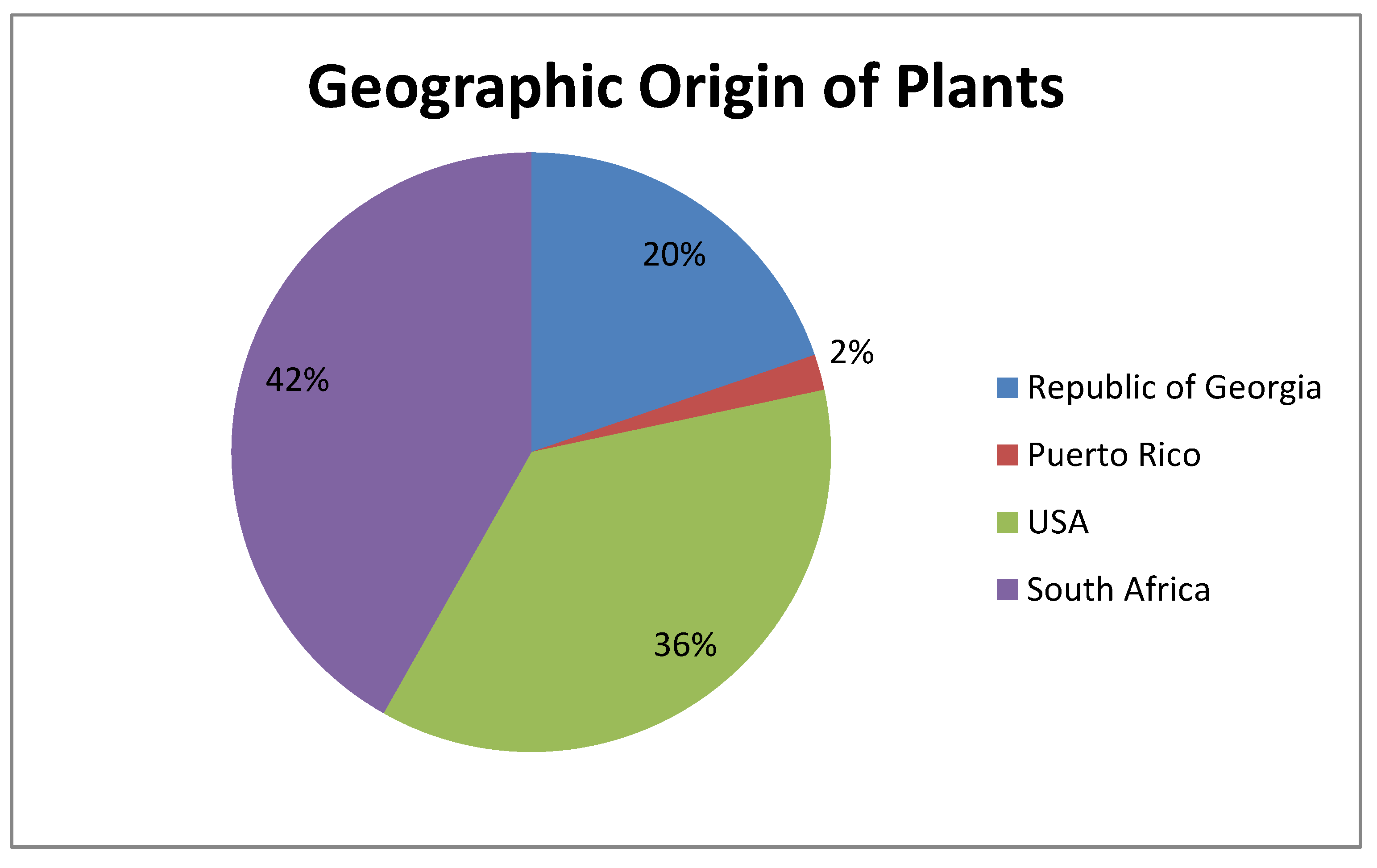
| Group | No. | Family | Genus | Species | Extraction Origin (Plant Part) | Geographicorigin | MIC (mg/mL) |
|---|---|---|---|---|---|---|---|
| A | 1 | Meliaceae | Trichilia | emetica | Leaves | South Africa | 10 |
| 2 | Passifloraceae | Passiflora | foetida | Whole plant | South Africa | 5 | |
| 3 | Lamiaceae | Salvia | nemorosa | Whole plant | Georgia | 5 | |
| 4 | Sambucaceae | Sambucus | ebulus | Whole plant | Georgia | 10 | |
| B | 5 | Fabaceae | Baphia | racemosa | Root | South Africa | 2.5 |
| 6 | Dracaenaceae | Sansevieria | hyacinthoides | Root | South Africa | 2.5 | |
| 7 | Fabaceae | Desmodium | adscendens | Whole plant | South Africa | 5 | |
| 8 | Fabaceae | Eriosema | preptum | Whole plant | South Africa | 10 | |
| C | 9 | Sarraceniaceae | Darlingtonia | californica | Leaves | USA | 10 |
| 10 | Pedaliaceae | Proboscidea | louisianica | Seed pod | USA | 10 | |
| 11 | Betulaceae | Alnus | barbata | Leaves + twigs | Georgia | 5 | |
| 12 | Ophioglossaceae | Botrychium | multifidum | Root | USA | 5 |
| Group | No. | Genus | Species | MIC (mg/mL) | Cell Damage (SEM) |
|---|---|---|---|---|---|
| A | 1 | Trichilia | emetica | 10 | Loss of flagella |
| 2 | Passiflora | foetida | 5 | Loss of flagella, holes in the external bacteria wall | |
| 3 | Salvia | nemorosa | 5 | Loss of flagella, severe cell collapse and deformation | |
| 4 | Sambucus | ebulus | 10 | Loss of flagella | |
| B | 5 | Baphia | racemosa | 2.5 | Loss of flagella |
| 6 | Sansevieria | hyacinthoides | 2.5 | Loss of flagella, cell deformation | |
| 7 | Desmodium | adscendens | 5 | Loss of flagella | |
| 8 | Eriosema | preptum | 10 | Loss of flagella | |
| C | 9 | Darlingtonia | californica | 10 | Loss of flagella, severe cell collapse and deformation |
| 10 | Proboscidea | louisianica | 10 | Loss of flagella, leakage of intracellular components | |
| 11 | Alnus | barbata | 5 | Loss of flagella, severe cell collapse and deformation | |
| 12 | Botrychium | multifidum | 5 | Cell destruction |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceruso, M.; Clement, J.A.; Todd, M.J.; Zhang, F.; Huang, Z.; Anastasio, A.; Pepe, T.; Liu, Y. The Inhibitory Effect of Plant Extracts on Growth of the Foodborne Pathogen, Listeria monocytogenes. Antibiotics 2020, 9, 319. https://doi.org/10.3390/antibiotics9060319
Ceruso M, Clement JA, Todd MJ, Zhang F, Huang Z, Anastasio A, Pepe T, Liu Y. The Inhibitory Effect of Plant Extracts on Growth of the Foodborne Pathogen, Listeria monocytogenes. Antibiotics. 2020; 9(6):319. https://doi.org/10.3390/antibiotics9060319
Chicago/Turabian StyleCeruso, Marina, Jason A. Clement, Matthew J. Todd, Fangyuan Zhang, Zuyi Huang, Aniello Anastasio, Tiziana Pepe, and Yanhong Liu. 2020. "The Inhibitory Effect of Plant Extracts on Growth of the Foodborne Pathogen, Listeria monocytogenes" Antibiotics 9, no. 6: 319. https://doi.org/10.3390/antibiotics9060319
APA StyleCeruso, M., Clement, J. A., Todd, M. J., Zhang, F., Huang, Z., Anastasio, A., Pepe, T., & Liu, Y. (2020). The Inhibitory Effect of Plant Extracts on Growth of the Foodborne Pathogen, Listeria monocytogenes. Antibiotics, 9(6), 319. https://doi.org/10.3390/antibiotics9060319