Acacia senegal Extract Rejuvenates the Activity of Phenicols on Selected Enterobacteriaceae Multi Drug Resistant Strains

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants Material and Extraction
2.1.1. Hydroethanolic Extracts (HE)
2.1.2. Aqueous Extract
2.1.3. Soxhlet Extraction
2.2. Phytochemical Composition
2.2.1. High-Performance Thin Layer Chromatography (HPTLC) Screening
2.2.2. Determination of Total Phenolic Content
2.2.3. Determination of Total Flavonoids Content
2.2.4. Determination of Tannin Content
Hydrolyzable Tannins
Condensed Tannins
2.3. Bacterial Strains
2.4. Determination of Minimal Inhibitory Concentration (MIC) of the Plant Extracts
2.5. Combination with Antibiotics
2.6. Outer Membrane Permeation Assay
2.7. Statistical Analysis
3. Results
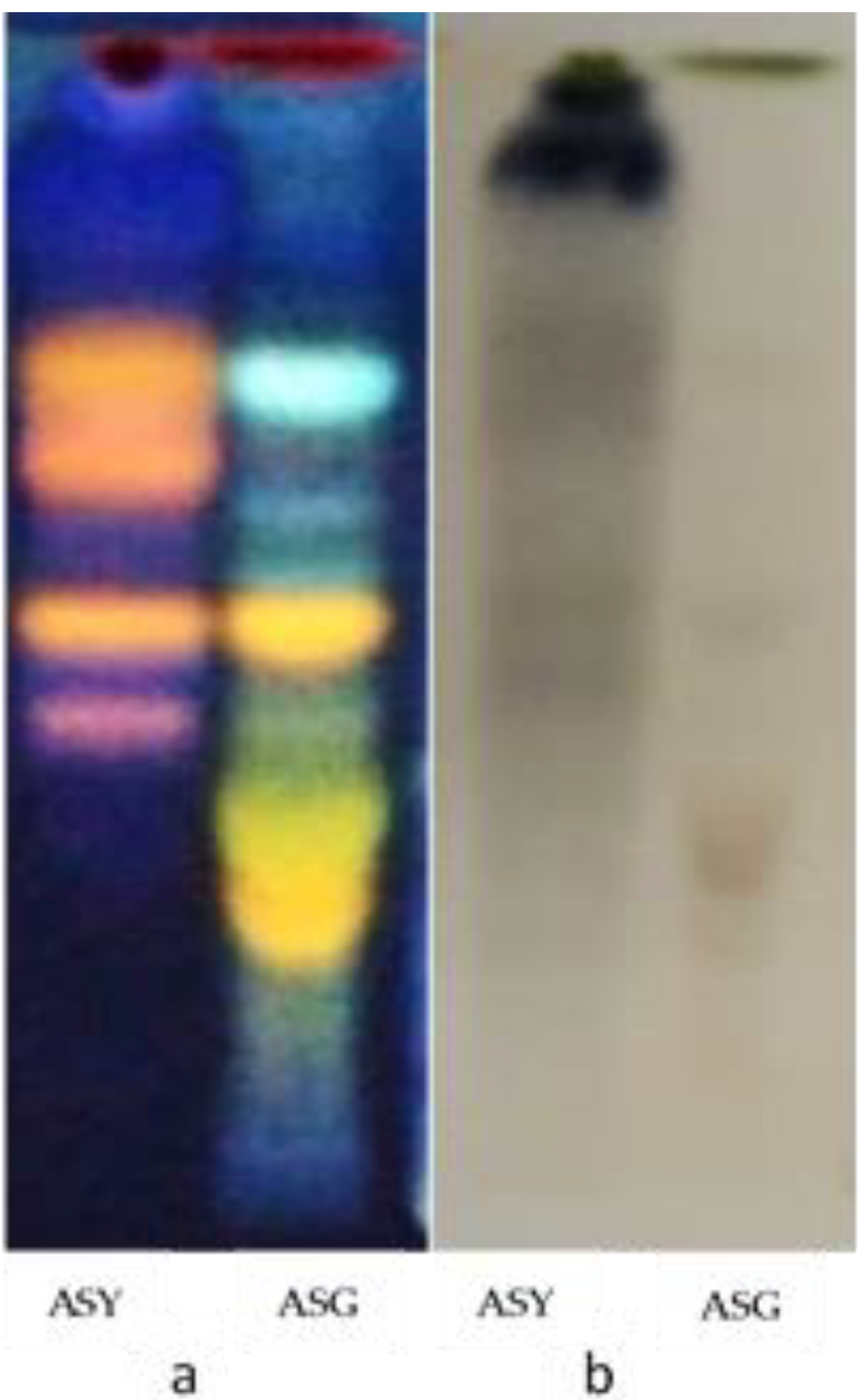
3.1. Phytochemical Screening by HPTLC
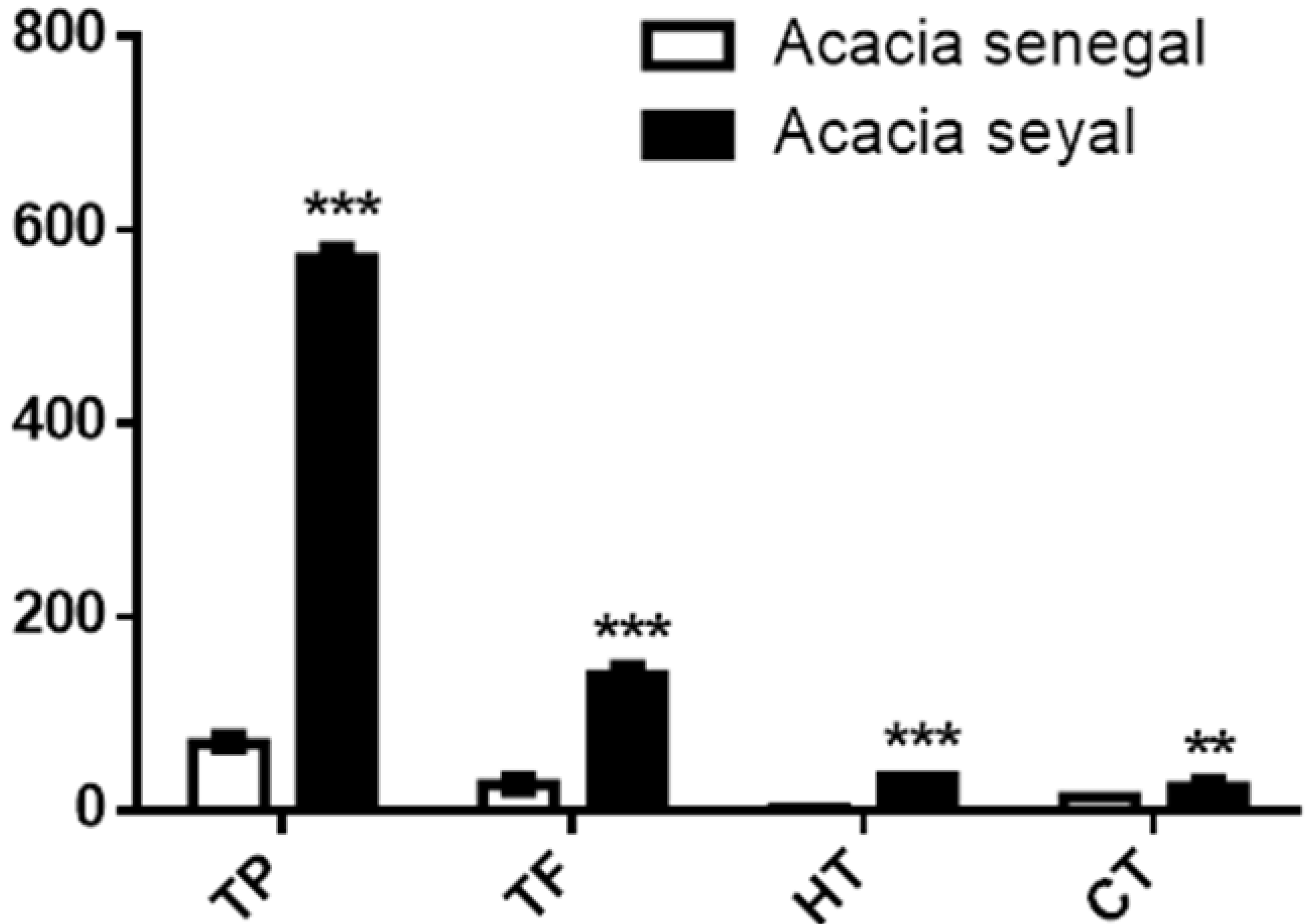
3.2. Phytochemical Contents
3.3. Antibacterial Effect of the Various Extract
3.4. Combination with Different Antibiotics
3.5. Effect on Membrane Permeability
3.6. Combination Tests with Phenicols
3.7. FloR Pump and A. senegal HE
4. Discussion
5. Conclusions
Ethics Approval and Consent to Participate
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Principes méthodologiques généraux pour la recherche et l’évaluation relatives à la médecine traditionnelle; Genève. Available online: https://apps.who.int/iris/handle/10665/68476 (accessed on 6 August 2019).
- Stratégie de l’OMS pour la médecine traditionnelle pour 2002–2005. Available online: http://archives.who.int/tbs/trm/s2298f.pdf (accessed on 17 June 2019).
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Aspects Med. 2006, 27, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Guinoiseau, E.; Luciani, A.; Rossi, P.G.; Quilichini, Y.; Ternengo, S.; Bradesi, P.; Berti, L. Cellular effects induced by Inula graveolens and Santolina corsica essential oils on Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 873–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosa, L.; Cañizares, E. Obtención, caracterización y evaluación de la actividad antimicrobiana de extractos de propóleos de Campeche. Ars Pharm. Internet 2002, 43, 187–204. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Silber, J.; Kramer, A.; Labes, A.; Tasdemir, D. From discovery to production: Biotechnology of marine fungi for the production of new antibiotics. Mar. Drugs 2016, 14, 137. [Google Scholar] [CrossRef] [Green Version]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Traditional Medicinal Plants. Available online: http://www.ethnopharmacologia.org/prelude2018/pdf/biblio-hr-17-ruffo.pdf (accessed on 6 August 2019).
- Tapsoba, H.; Deschamps, J.-P. Use of medicinal plants for the treatment of oral diseases in Burkina Faso. J. Ethnopharmacol. 2006, 104, 68–78. [Google Scholar] [CrossRef]
- Iyamah, P.C.; Idu, M. Ethnomedicinal survey of plants used in the treatment of malaria in Southern Nigeria. J. Ethnopharmacol. 2015, 173, 287–302. [Google Scholar] [CrossRef]
- Magnini, R.D. A Review on ethnobotanical uses, biological activities and phytochemical aspects of Acacia senegal (L.) Willd. and Acacia seyal Delile.(Fabaceae). Int. J. Plant Sci. Hor. 2020, 2, 32–55. [Google Scholar] [CrossRef]
- Hammiche, V.; Maiza, K. Traditional medicine in Central Sahara: Pharmacopoeia of Tassili N’ajjer. J. Ethnopharmacol. 2006, 105, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Teklehaymanot, T. An ethnobotanical survey of medicinal and edible plants of Yalo Woreda in Afar regional state, Ethiopia. J. Ethnobiol. Ethnomed. 2017, 13, 40. [Google Scholar] [CrossRef]
- Mariod, A.A.; Fadle, N.; Hasan, A.A. Antimicrobial screening of wood extracts of Combretum hartmannianum, Acacia seyal and Terminalia brownie. Eur. J. Mol. Biol. Biochem. 2014, 1, 77–80. [Google Scholar]
- Samrot, A.V.; Sahiti, K.; Raji, P.; Rohan, B.; Kumar, D.; Sharma, K. TLC bio-autography guided identification of antioxidant and antibacterial activity of Acacia senegal. Pharm. Lett. 2016, 8, 41–47. [Google Scholar]
- Abdllha, H.B.; Mohamed, A.I.; Almoniem, K.A.; Adam, N.I.; Alhaadi, W.; Elshikh, A.A.; Ali, A.J.; Makuar, I.G.; Elnazeer, A.M.; Elrofaei, N.A. Evolution of antimicrobial, antioxidant potentials and phytochemical studies of three solvent extracts of five species from acacia used in Sudanese ethnomedicine. Adv. Microbiol. 2016, 6, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Satish, A.; Asna, U. Quantification of flavonoids by UPLC-MS and its antibacterial activity from Brassica oleracea var. Capitata L. GSC Biol. Pharm. Sci. 2018, 5, 109–114. [Google Scholar] [CrossRef]
- Bagre, I.; Bahi, C.; Gnahoue, G.; Djaman, A.J.; Guede, G.F. Composition phytochimique et évaluation in-vitro de l’activité antifongique des extraits des feuilles de Morinda morindoïdes (baker) Milne-Redhead (Rubiaceae) sur Aspergillus fumigatus et candida albicans. Int. J. Sci. Pharm. Biol. Sci. 2007, 8, 15–23. [Google Scholar]
- Bhadoriya, S.S.; Mishra, V.; Raut, S.; Ganeshpurkar, A.; Jain, S.K. Anti-inflammatory and antinociceptive activities of a hydroethanolic extract of Tamarindus indica leaves. Sci. Pharm. 2012, 80, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Simao, A.A.; Marques, T.R.; Marcussi, S.; Correa, A.D. Aqueous extract of Psidium guajava leaves: Phenolic compounds and inhibitory potential on digestive enzymes. An. Acad. Bras. Ciênc. 2017, 89, 2155–2165. [Google Scholar] [CrossRef]
- Nitiéma, M.; Soleti, R.; Koffi, C.; Belemnaba, L.; Mallegol, P.; Ouédraogo, N.; Kini, F.B.; Ouédraogo, S.; Guissou, I.P.; Andriantsitohaina, R. Ethyl Acetate Fraction of Lannea microcarpa Engl. and K. Krause (Anacardiaceae) Trunk Barks Corrects Angiotensin II-Induced Hypertension and Endothelial Dysfunction in Mice. Oxid. Med. Cell. Longev. 2019, 2019, 9464608. [Google Scholar]
- Mandana, B.; Russly, A.R.; Farah, S.T.; Noranizan, M.A.; Zaidul, I.S.; Ali, G. Antioxidant activity of winter melon (Benincasa hispida) seeds using conventional Soxhlet extraction technique. Int. Food Res. J. 2012, 19, 229–234. [Google Scholar]
- Hassim, N.; Markom, M.; Anuar, N.; Dewi, K.H.; Baharum, S.N.; Mohd Noor, N. Antioxidant and antibacterial assays on polygonum minus extracts: Different extraction methods. Int. J. Chem. Eng. 2015, 2015, 826709. [Google Scholar] [CrossRef]
- Nitiema, L.W.; Sombié, P.A.; Koala, M.; Del Fiore, A. Phytochemical composition and antioxidant activity of Balanites aegyptiaca, Securidaca longepedunculata and Acacia gourmaensis used against seed-borne fungi in Burkina Faso. Curr. J. Appl. Sci. Technol. 2020, 39, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Alothman, M.; Bhat, R.; Karim, A.A. Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvents. Food Chem. 2009, 115, 785–788. [Google Scholar] [CrossRef]
- Mole, S.; Butler, L.G.; Hagerman, A.E.; Waterman, P.G. Ecological tannin assays: A critique. Oecologia 1989, 78, 93–96. [Google Scholar] [CrossRef]
- Ba, K.; Tine, E.; Destain, J.; Cissé, N.; Thonart, P. Étude comparative des composés phénoliques, du pouvoir antioxydant de différentes variétés de sorgho sénégalais et des enzymes amylolytiques de leur malt. Biotechnol. Agron. Société Environ. 2010, 14, 131–139. [Google Scholar]
- Vergalli, J.; Dumont, E.; Cinquin, B.; Maigre, L.; Pajovic, J.; Bacqué, E.; Mourez, M.; Réfrégiers, M.; Pagès, J.-M. Fluoroquinolone structure and translocation flux across bacterial membrane. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Braibant, M.; Chevalier, J.; Chaslus-Dancla, E.; Pages, J.-M.; Cloeckaert, A. Structural and functional study of the phenicol-specific efflux pump FloR belonging to the major facilitator superfamily. Antimicrob. Agents Chemother. 2005, 49, 2965–2971. [Google Scholar] [CrossRef] [Green Version]
- Ghisalberti, D.; Masi, M.; Pagès, J.-M.; Chevalier, J. Chloramphenicol and expression of multidrug efflux pump in Enterobacter aerogenes. Biochem. Biophys. Res. Commun. 2005, 328, 1113–1118. [Google Scholar] [CrossRef]
- Masi, M.; Pagès, J.-M.; Pradel, E. Overexpression and purification of the three components of the Enterobacter aerogenes AcrA–AcrB–TolC multidrug efflux pump. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 786, 197–205. [Google Scholar] [CrossRef]
- Mallea, M.; Chevalier, J.; Bornet, C.; Eyraud, A.; Davin-Regli, A.; Bollet, C.; Pages, J.-M. Porin alteration and active efflux: Two in vivo drug resistance strategies used by Enterobacter aerogenes. Microbiology 1998, 144, 3003–3009. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, J.; Mahamoud, A.; Baitiche, M.; Adam, E.; Viveiros, M.; Smarandache, A.; Militaru, A.; Pascu, M.L.; Amaral, L.; Pagès, J.-M. Quinazoline derivatives are efficient chemosensitizers of antibiotic activity in Enterobacter aerogenes, Klebsiella pneumoniae and Pseudomonas aeruginosa resistant strains. Int. J. Antimicrob. Agents 2010, 36, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pages, J.-M.; Lavigne, J.-P.; Leflon-Guibout, V.; Marcon, E.; Bert, F.; Noussair, L.; Nicolas-Chanoine, M.-H. Efflux pump, the masked side of beta-lactam resistance in Klebsiella pneumoniae clinical isolates. PLoS ONE 2009, 4, e4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, J.H.; Turnidge, J.D. Susceptibility test methods: Dilution and disk diffusion methods. In Manual of Clinical Microbiology, 8th ed.; ASM Press: Washington, DC, USA, 2003; Volume 2, pp. 1108–1127. [Google Scholar]
- Fadli, M.; Saad, A.; Sayadi, S.; Chevalier, J.; Mezrioui, N.-E.; Pagès, J.-M.; Hassani, L. Antibacterial activity of Thymus maroccanus and Thymus broussonetii essential oils against nosocomial infection–bacteria and their synergistic potential with antibiotics. Phytomedicine 2012, 19, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Borselli, D.; Lieutaud, A.; Thefenne, H.; Garnotel, E.; Pagès, J.-M.; Brunel, J.M.; Bolla, J.-M. Polyamino-isoprenic derivatives block intrinsic resistance of P. aeruginosa to doxycycline and chloramphenicol in vitro. PLoS ONE 2016, 11, e0154490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infe,ctions: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaara, M.; Siikanen, O.; Apajalahti, J.; Fox, J.; Frimodt-Møller, N.; He, H.; Poudyal, A.; Li, J.; Nation, R.L.; Vaara, T. A novel polymyxin derivative that lacks the fatty acid tail and carries only three positive charges has strong synergism with agents excluded by the intact outer membrane. Antimicrob. Agents Chemother. 2010, 54, 3341–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolla, J.-M.; Alibert-Franco, S.; Handzlik, J.; Chevalier, J.; Mahamoud, A.; Boyer, G.; Kieć-Kononowicz, K.; Pagès, J.-M. Strategies for bypassing the membrane barrier in multidrug resistant Gram-negative bacteria. FEBS Lett. 2011, 585, 1682–1690. [Google Scholar] [CrossRef]
- Masi, M.; Réfregiers, M.; Pos, K.M.; Pagès, J.-M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Sharma, P.; Bhagchandani, T.; Jain, S.C. Phytochemical investigation and antimicrobial activity of Acacia senegal root heartwood. J. Pharm. Res. 2012, 5, 4934–4938. [Google Scholar]
- Mudi, S.Y.; Salisu, A. Studies on brine shrimp lethality and activity of stem bark extract of Acacia senegal L. on respiratory tract pathogenic bacteria. Int. J. Biomed. Hlth. Sci. 2009, 5, 139–143. [Google Scholar]
- Sibanda, T.; Okoh, A.I. The challenges of overcoming antibiotic resistance: Plant extracts as potential sources of antimicrobial and resistance modifying agents. Afr. J. Biotechnol. 2007, 6, 25. [Google Scholar]
- Radulovic, N.S.; Blagojevic, P.D.; Stojanovic-Radic, Z.Z.; Stojanovic, N.M. Antimicrobial plant metabolites: Structural diversity and mechanism of action. Curr. Med. Chem. 2013, 20, 932–952. [Google Scholar] [PubMed]
- Perumal, S.; Mahmud, R.; Ismail, S. Mechanism of action of isolated caffeic acid and epicatechin 3-gallate from Euphorbia hirta against Pseudomonas aeruginosa. Pharmacogn. Mag. 2017, 13, S311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadiq, M.B.; Tarning, J.; Aye Cho, T.Z.; Anal, A.K. Antibacterial activities and possible modes of action of Acacia nilotica (L.) Del. against multidrug-resistant Escherichia coli and Salmonella. Molecules 2017, 22, 47. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.-M.; Amaral, L.; Bolla, J.-M. Geraniol restores antibiotic activities against multidrug-resistant isolates from gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [Green Version]
- Dzotam, J.K.; Touani, F.K.; Kuete, V. Antibacterial activities of the methanol extracts of Canarium schweinfurthii and four other Cameroonian dietary plants against multi-drug resistant Gram-negative bacteria. Saudi J. Biol. Sci. 2016, 23, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.; Nazir, M.; Ali, M.S.; Hussain, H.; Lee, Y.S.; Riaz, N.; Jabbar, A. Antimicrobial natural products: An update on future antibiotic drug candidates. Nat. Prod. Rep. 2010, 27, 238–254. [Google Scholar] [CrossRef]
- Siriyong, T.; Srimanote, P.; Chusri, S.; Yingyongnarongkul, B.; Suaisom, C.; Tipmanee, V.; Voravuthikunchai, S.P. Conessine as a novel inhibitor of multidrug efflux pump systems in Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2017, 17, 405. [Google Scholar] [CrossRef]
- Belofsky, G.; Percivill, D.; Lewis, K.; Tegos, G.P.; Ekart, J. Phenolic metabolites of dalea v ersicolor that enhance antibiotic activity against model pathogenic bacteria. J. Nat. Prod. 2004, 67, 481–484. [Google Scholar] [CrossRef]
- Holler, J.G.; Christensen, S.B.; Slotved, H.-C.; Rasmussen, H.B.; Gúzman, A.; Olsen, C.-E.; Petersen, B.; Mølgaard, P. Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Oluwatuyi, M.; Kaatz, G.W.; Gibbons, S. Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 2004, 65, 3249–3254. [Google Scholar] [CrossRef] [PubMed]
- Lôme, V.; Brunel, J.-M.; Bolla, J.-M. Multiparametric profiling for identification of chemosensitizers against Gram-negative bacteria. Front. Microbiol. 2018, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Malmir, S.; Bahreinian, M.; Zahiri Yeganeh, S.; Mirnejad, R.; Moosazadeh Moghaddam, M.; Saberi, F. Molecular mechanisms of resistance to conventional antibiotics in bacteria. Int. J. Med. Rev. 2018, 5, 118–129. [Google Scholar] [CrossRef]
- Milhaud, G. Les résidus de chloramphénicol et leur toxicité. Ann. Rech. Vét. 1985, 16, 133–148. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Strains | Description | Reference |
|---|---|---|
| E. coli | ||
| AG100 | Parental E. coli K-12 Porin +; basal efflux | [29] |
| AG100A | AG100 acrAB::Kanr non (AcrAB-); Porin + | [29] |
| AG102 | AG100 overexpressing the AcrAB pump, Porin + | [29] |
| AG1004 plasmid AG100A FloR | wid plasmid without floR Expressing floR | [30] [30] |
| K. aerogenes | ||
| Ea CM64 | CHL variant obtained from ATCC 13048 overexpressing the AcrAB pump; Porin+ | [31] |
| Ea 289 | KAN-sensitive derivative of EA27, Porin- | [32] |
| Ea 298 | EA289 tolC::Kanr; Porin- | [32] |
| EaATCC 15038 | Porin +; Normal efflux | [33] |
| K. pneumoniae | ||
| ATCC 12296 | Reference strain; Porin +; Normal efflux | [34] |
| KP 45 | Porin +; Normal efflux | [35] |
| KP55 | Clinical MDR isolate; Tetr Ampr Atmr Cefr; Porin-; Normal efflux | [35] |
| Antibiotics | E. coli | K. aerogenes | K. pneumoniae | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AG100 | AG100A | AG102 | Ea 289 | Ea298 | Ea ATCC 15038 | CM64 | KP45 | KP55 | KPATCC 11296 | |
| Chloramphenicol | 8 | 1 | 64 | 1024 | 32 | 4 | 512 | 8 | 16 | 8 |
| Florfenicol | 16 | 1 | 128 | 256 | 4 | 8 | 512 | 8 | 8 | 16 |
| Thiamphenicol | 256 | 16 | 1024 | >1024 | >1024 | 64 | >1024 | 128 | 256 | 256 |
| Ciprofloxacin | 0.03 | 0.008 | 0.25 | 32 | 4 | <0.125 | 0.5 | 0.06 | 4 | 0.06 |
| Cefepime | <0.125 | <0.125 | 0.25 | 2 | 8 | <0.125 | 0.25 | <0.125 | 16 | <0.125 |
| Ceftazidime | 0.5 | 0.25 | 2 | >64 | >64 | 0.03 | 2 | 0.25 | >64 | 0.5 |
| Plant Extract | E. coli | K. aerogenes | K. Pneumoniae | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AG100 | AG100A | AG102 | Ea 289 | Ea298 | Ea ATCC 15038 | CM64 | KP45 | KP55 | KP ATCC 11296 | |
| Soxhlet extraction Acacia senegal(leaves) | ||||||||||
| Hexane extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Dichloromethane extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Ethyl acetate extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Methanolic extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Acacia seyal(leaves) | ||||||||||
| Hexane extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Dichloromethane extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Ethyl acetate extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Methanolic extract | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Cold-Maceration | ||||||||||
| Acacia senegal | ||||||||||
| Hydroethanolic extract (leaves) | 256 | 128 | >512 | >512 | 128 | 256 | >512 | >512 | 512 | >512 |
| Hydroethanolic (Root Bark) | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Acacia seyal | ||||||||||
| hydroethanolic (Leaves) | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Hydroethanolic (Root Bark) | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Aqueous | ||||||||||
| Acacia senegal | ||||||||||
| Root bark | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Acacia seyal | ||||||||||
| Root bark | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| E. coli | K. pneumoniae | K. aerogenes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plants | Extracts | ATBs | Extracts (mg/L) | AG100 | AG100A | AG102 | KP45 | KP55 | KP ATCC 11296 | Ea289 | Ea298 | Ea ATCC 15038 | Ea CM64 |
| A. senegal | Decoction root bark | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 |
| 512 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 64 | 4 | 512 | |||
| 128 | 16 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| 16 | 16 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| CIP | 0 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 512 | 0.03 | 0.008 | 0.25 | 0.03 | 4 | 0.06 | 64 | 8 | <0.125 | 1 | |||
| 128 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 1 | |||
| 16 | 0.03 | 0.008 | nd | nd | 4 | Nd | 64 | 4 | <0.125 | 0.5 | |||
| CAZ | 0 | 0.5 | 0.25 | 2 | 0.25 | >64 | 0.25 | >64 | >64 | 0.03 | 2 | ||
| 512 | 1 | 0.5 | 2 | 0.5 | >64 | 0.25 | >64 | >64 | 0.0.3 | 2 | |||
| 128 | 0.5 | 0.25 | 2 | 0.5 | >64 | 0.25 | >64 | >64 | 0.03 | 2 | |||
| 16 | 1 | <0.125 | nd | nd | >64 | nd | >64 | >64 | >2 | 2 | |||
| Hydroethanolic root bark | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |
| 512 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| 128 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| 16 | 8 | 1 | 64 | 8 | 16 | 16 | 1024 | 32 | 4 | 512 | |||
| CIP | 0 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 512 | 0.03 | 0.008 | 0.25 | 0.125 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | |||
| 128 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.125 | 64 | 4 | <0.125 | 0.5 | |||
| 16 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | |||
| CEF | 0 | 0.06 | <0.03 | 0.5 | <0.125 | 16 | <0.125 | 2 | 8 | <0.125 | 0.25 | ||
| 512 | 0.06 | <0.03 | 0.5 | 0.5 | 8 | <0.125 | 4 | 8 | <0.125 | 0.25 | |||
| 128 | 0.06 | <0.03 | 0.5 | 0.06 | 8 | <0.125 | 2 | 16 | <0.125 | 0.25 | |||
| 16 | 0.06 | <0.03 | 0.5 | 0.06 | 8 | <0.125 | 2 | 16 | <0.125 | 0.25 | |||
| hydroethanolic leaves extract | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |
| 512 | 1 | <0.125 | 2 | 2 | 1 | 2 | 64 | 4 | <2 | 16 | |||
| 128 | 2 | 1 | 4 | 2 | 4 | 2 | 128 | 16 | <2 | 32 | |||
| 16 | 8 | 2 | 64 | 8 | 16 | 4 | 1024 | 32 | 4 | 512 | |||
| CIP | 0 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 512 | 0.03 | <0.008 | 0.125 | 0.06 | 1 | 0.06 | 64 | 1 | <0.125 | <0.125 | |||
| 128 | 0.03 | <0.008 | 0.25 | 0.06 | 2 | 0.06 | 64 | 2 | <0.125 | <0.125 | |||
| 16 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.03 | 64 | 2 | <0.125 | 0.5 | |||
| CAZ | 0 | 0.5 | 0.25 | 2 | 0.25 | >64 | 0.25 | >64 | >64 | 0.03 | 2 | ||
| 512 | 0.25 | <0.125 | 0.5 | 0.5 | >64 | 2 | >64 | >64 | 0.03 | 1 | |||
| 128 | 0.5 | 0.25 | 1 | 0.5 | >64 | 1 | >64 | >64 | 0.03 | 2 | |||
| 16 | 0.25 | 0.25 | 1 | 0.25 | >64 | 0.5 | >64 | >64 | 0.03 | 2 | |||
| FLOR | 0 | 16 | 1 | 128 | 8 | 8 | 16 | 256 | 4 | 8 | 512 | ||
| 512 | 0.5 | <0.125 | 4 | 2 | 2 | 2 | 8 | 0.25 | 2 | 32 | |||
| 128 | 2 | 0.5 | 8 | 4 | 2 | 2 | 16 | 1 | 2 | 32 | |||
| 16 | 16 | 1 | 128 | 8 | 8 | 8 | 256 | 2 | 8 | 256 | |||
| THIAM | 0 | 256 | 16 | 1024 | 128 | 256 | 256 | >1024 | >1024 | 64 | >1024 | ||
| 512 | 8 | <0.125 | 128 | 128 | 64 | 64 | >1024 | 4 | 64 | 512 | |||
| 128 | 128 | 4 | 256 | 128 | 64 | 128 | >1024 | 1024 | 128 | 1024 | |||
| 16 | 128 | 16 | 1024 | 128 | 128 | 256 | >1024 | >1024 | >1024 | >1024 | |||
| A. seyal | Decoction root bark | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 |
| 512 | 8 | 1 | 32 | 8 | 16 | 2 | 1024 | 32 | 4 | 256 | |||
| 128 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| 16 | 8 | 1 | 64 | 16 | >64 | 16 | 1024 | 32 | 4 | 1024 | |||
| CIP | 0 | 0.06 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 512 | 0.06 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 8 | <0.125 | 0.5 | |||
| 128 | 0.06 | 0.008 | 0.5 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | |||
| 16 | 0.06 | 0.008 | 0.25 | 0.5 | 8 | 0.06 | 64 | 4 | <0.125 | 0.5 | |||
| CAZ | 0 | 0.5 | 0.25 | 2 | 0.25 | >64 | 0.25 | >64 | >64 | 0.03 | 2 | ||
| 512 | 0.5 | 0.5 | 2 | 2 | >64 | 0.125 | >64 | >64 | 0.03 | 2 | |||
| 128 | 0.5 | 0.5 | 2 | 1 | >64 | 0.5 | >64 | >64 | 0.03 | 2 | |||
| 16 | 1 | 0.25 | 2 | 1 | >64 | 0.5 | >64 | >64 | 0.06 | 4 | |||
| Hydroethanolic root bark | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |
| 128 | 8 | <0.125 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |||
| 16 | 16 | <0.125 | 64 | 8 | 16 | 8 | 1024 | 64 | 8 | 512 | |||
| 1 | 8 | 0.25 | 64 | 8 | 16 | 8 | 1024 | 32 | 512 | 512 | |||
| CIP | 0 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 128 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | |||
| 16 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 2 | <0.125 | 0.5 | |||
| 1 | 0.03 | 0.008 | 0.5 | 0.06 | 4 | 0.06 | 64 | 2 | <0.125 | 0.5 | |||
| CEF | 0 | <0.125 | <0.125 | 0.25 | 2 | 8 | 0.125 | 2 | 16 | <0.125 | 0.25 | ||
| 128 | <0.125 | <0.125 | 0.5 | 0.125 | 8 | 0.125 | 4 | 8 | <0.125 | 0.25 | |||
| 16 | <0.125 | <0.125 | 0.25 | 0.125 | 8 | 0.125 | 4 | 8 | <0.125 | 0.25 | |||
| 1 | 0.06 | <0.125 | 0.5 | 0.25 | 16 | 0.125 | 4 | 16 | <0.125 | 0.25 | |||
| Hydroethanolic leaves extract | CHL | 0 | 8 | 1 | 64 | 8 | 16 | 8 | 1024 | 32 | 4 | 512 | |
| 512 | 2 | 1 | 16 | 4 | 4 | 4 | 256 | 64 | 2 | 128 | |||
| 128 | 4 | 1 | 32 | 4 | 16 | 8 | 1024 | 128 | 4 | 512 | |||
| 16 | 8 | 1 | 64 | 4 | 16 | 8 | 1024 | 128 | 4 | 512 | |||
| CIP | 0 | 0.03 | 0.008 | 0.25 | 0.06 | 4 | 0.06 | 64 | 4 | <0.125 | 0.5 | ||
| 512 | 0.03 | 0.016 | 0.25 | 0.06 | 8 | 0.06 | >64 | 4 | <0.125 | 0.5 | |||
| 128 | 0.03 | 0.008 | 0.125 | 0.06 | 4 | 0.125 | 64 | 8 | <0.125 | 0.5 | |||
| 16 | 0.03 | 0.008 | 0.125 | 0.06 | 4 | 0.125 | 64 | 4 | <0.125 | 0.5 | |||
| CAZ | 0 | 0.5 | 0.25 | 2 | 0.25 | >64 | 0.25 | >64 | >64 | 0.03 | 2 | ||
| 512 | 0.5 | 0.25 | 0.25 | 0.25 | >64 | 0.5 | >64 | >64 | 0.03 | 1 | |||
| 128 | 0.5 | 0.25 | 1 | 0.25 | >64 | 0.5 | >64 | >64 | 0.03 | 2 | |||
| 16 | 0.5 | 0.25 | 0.5 | 0.25 | >64 | 0.25 | >64 | >64 | 0.06 | 1 | |||
| Permeating Effect | ||||||
|---|---|---|---|---|---|---|
| Concentration extract (mg/L) | 256 | 128 | 64 | 32 | 16 | 8 |
| % of polymyxin B | 50 (±18 *) | 31 (±13 *) | 12 (±4 *) | 7.3 (±2 *) | 2 (±1 *) | 2 (±1 *) |
| Hydroethanolic Extract of Acacia Senegal (mg/L) | Methanolic Extract Acacia Senegal (mg/L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ATB | Strain | MIC ATB | MIC ASG | 8 | 16 | 32 | 64 | MIC ASG | 8 | 16 | 32 | 64 |
| CHL | E. coli | |||||||||||
| AG100 | 8 | 256 | 8 (-) | 4 (2) | 2 (4) | 1 (8) | >256 | 16 (-) | 8 (-) | 4 (2) | 2 (4) | |
| AG100A | 1 | 128 | 1 (-) | 2 (-) | 1 (-) | 0.5(2) | >256 | 1 (-) | 1 (-) | 1 (-) | 1 (-) | |
| AG102 | 64 | >256 | 64 (-) | 16 (4) | 4 (16) | 4 (16) | >256 | 64 (-) | 64 (-) | 32 (2) | 8 (8) | |
| K. aerogenes | ||||||||||||
| Ea289 | 1024 | >256 | 1024 (-) | 512 (2) | 64 (16) | 64 (16) | >256 | 1024 (-) | 1024 (-) | 512 (2) | 128 (8) | |
| Ea298 | 32 | 128 | 32 (-) | 32 (-) | 16 (2) | 4 (8) | >256 | 32 (-) | 32 (-) | 32 (-) | 16 (2) | |
| Ea ATCC 15038 | 4 | 256 | 4 (-) | 2 (2) | 1 (4) | 1 (4) | >256 | 4 (-) | 4 (-) | 2 (2) | 2 (2) | |
| Ea CM64 | 512 | >256 | 512 (-) | 128 (4) | 16 (32) | 16 (32) | >256 | 512 (-) | 256 (2) | 256 (2) | 32 (16) | |
| FLOR | E. coli | |||||||||||
| AG100 | 16 | 256 | 8 (2) | 4 (4) | 2 (8) | 2 (8) | >256 | 32 (-) | 16 (-) | 8 (2) | 4 (4) | |
| AG100A | 1 | 128 | 1 (-) | 1 (-) | 1 (-) | 0.25 (4) | >256 | 1 (-) | 1 (-) | 1 (-) | 1 (-) | |
| AG102 | 64 | >256 | 128 (-) | 32 (2) | 8 (8) | 4 (16) | >256 | 64 (-) | 64 (-) | 32 (2) | 16 (4) | |
| K. aerogenes | ||||||||||||
| Ea289 | 128 | >256 | 32 (4) | 64 (2) | 16 (8) | 8 (16) | >256 | 128 (-) | 64 (2) | 64 (2) | 32 (4) | |
| Ea298 | 4 | 128 | 4 (-) | 2 (2) | 2 (2) | 1 (4) | >256 | 4 (-) | 4 (-) | 4 (-) | 2 (2) | |
| Ea ATCC 15038 | 4 | 256 | 4 (-) | 2 (2) | 2 (2) | 2 (2) | >256 | 4(-) | 4 (-) | 4 (-) | 2 (2) | |
| Ea CM64 | 256 | >256 | 256 (-) | 128 (2) | 16 (16) | 16 (16) | >256 | 256 (-) | 256 (-) | 64 (4) | 32 (8) | |
| ATBs | Strain | MIC (mg/L) | Conc. HE Acacia senegal (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| E. coli | ATBs | ASG | 10 | 20 | 30 | 40 | 50 | 64 | |
| CHL | AG100 A | 2 | 128 | 2 | 2 | 2 | 2 | 2 | 2 |
| Plasmid | 1 | 128 | 1 | 1 | 1 | 1 | 1 | 1 | |
| FloR | 8 | 128 | 8 | 4 | 4 | 2 | 2 | 1 | |
| FLOR | AG100 A | 1 | 128 | 1 | 1 | 1 | 1 | 1 | 1 |
| Plasmid | 1 | 128 | 1 | 1 | 1 | 1 | 1 | 1 | |
| FloR | 4 - 2 | 128 | 4 | 4 | 1 | 1 | 1 | 0.5 | |
| THIAM | AG100 A | 16 | 128 | 16 | 8 | 8 | 8 | 4 | 4 |
| Plasmid | 16 | 128 | 16 | 8 | 8 | 8 | 4 | 4 | |
| FloR | 512 | 128 | 256 | 256 | 128 | 64 | 64 | 32 | |
| CAZ | AG100 A | 0.25 | 128 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| Plasmid | 0.25 | 128 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | |
| FloR | 0.25 | 128 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.125 | |
| NOR | AG100 A | 0.03 | 128 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Plasmid | 0.016 | 128 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | |
| FloR | 0.016 | 128 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | 0.016 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magnini, R.D.; Hilou, A.; Millogo-Koné, H.; Pagès, J.-M.; Davin-Regli, A. Acacia senegal Extract Rejuvenates the Activity of Phenicols on Selected Enterobacteriaceae Multi Drug Resistant Strains. Antibiotics 2020, 9, 323. https://doi.org/10.3390/antibiotics9060323
Magnini RD, Hilou A, Millogo-Koné H, Pagès J-M, Davin-Regli A. Acacia senegal Extract Rejuvenates the Activity of Phenicols on Selected Enterobacteriaceae Multi Drug Resistant Strains. Antibiotics. 2020; 9(6):323. https://doi.org/10.3390/antibiotics9060323
Chicago/Turabian StyleMagnini, René Dofini, Adama Hilou, Helana Millogo-Koné, Jean-Marie Pagès, and Anne Davin-Regli. 2020. "Acacia senegal Extract Rejuvenates the Activity of Phenicols on Selected Enterobacteriaceae Multi Drug Resistant Strains" Antibiotics 9, no. 6: 323. https://doi.org/10.3390/antibiotics9060323
APA StyleMagnini, R. D., Hilou, A., Millogo-Koné, H., Pagès, J. -M., & Davin-Regli, A. (2020). Acacia senegal Extract Rejuvenates the Activity of Phenicols on Selected Enterobacteriaceae Multi Drug Resistant Strains. Antibiotics, 9(6), 323. https://doi.org/10.3390/antibiotics9060323