Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom

 ,
,  ,
, 





 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
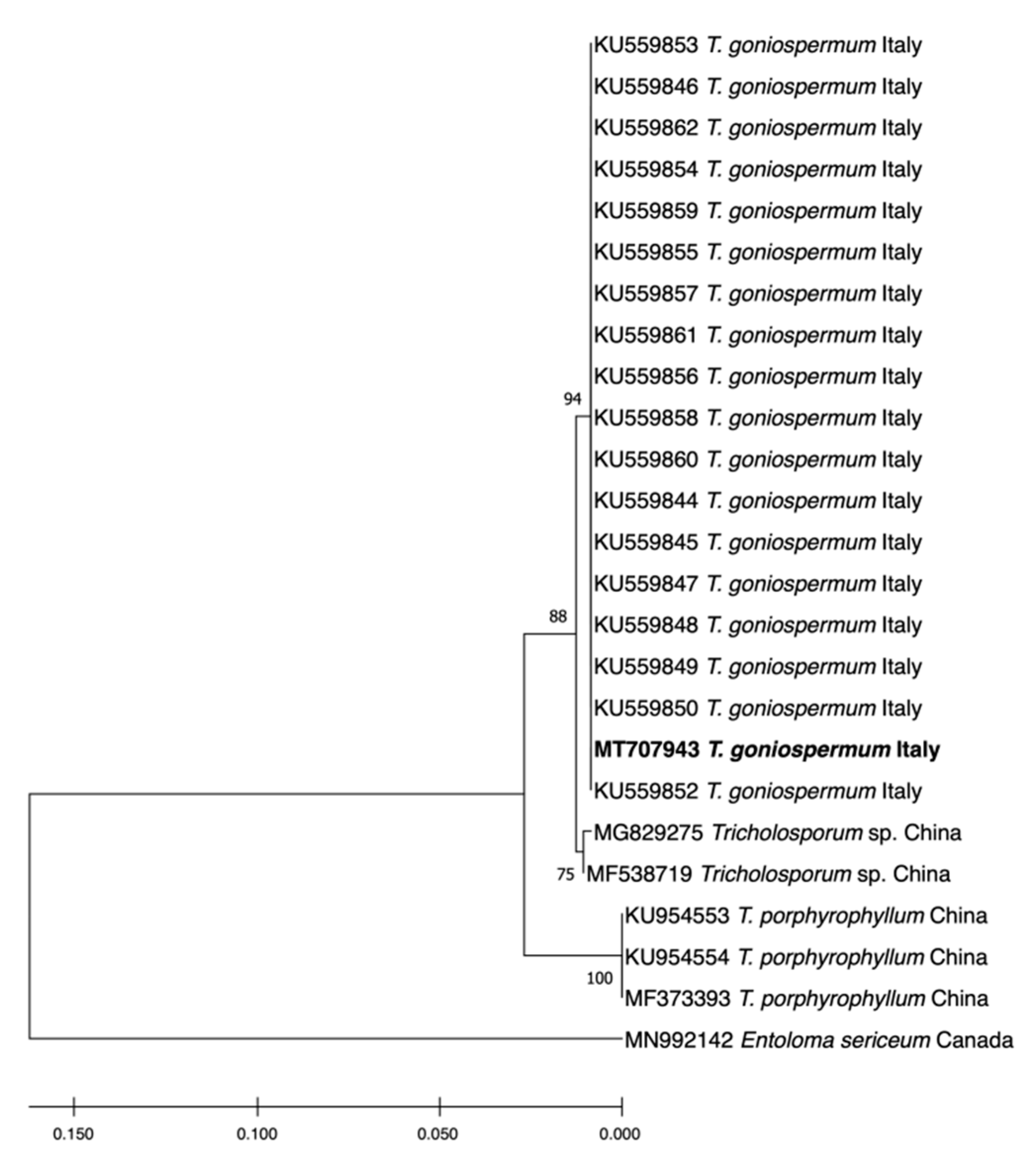
2.1. Molecular Identification of the T. goniospermum Strain
2.2. High Performance Liquid Chromatography (HPLC) Analysis
2.3. Antibacterial and Antifungal Activity
3. Materials and Methods
3.1. Mushroom Material
3.2. Molecular Identification
3.3. Preparation of Mushroom Extracts
3.4. Antioxidant and Enzyme Inhibitory Properties
3.5. HPLC Analysis
3.6. Antimicrobial Susceptibility Testing
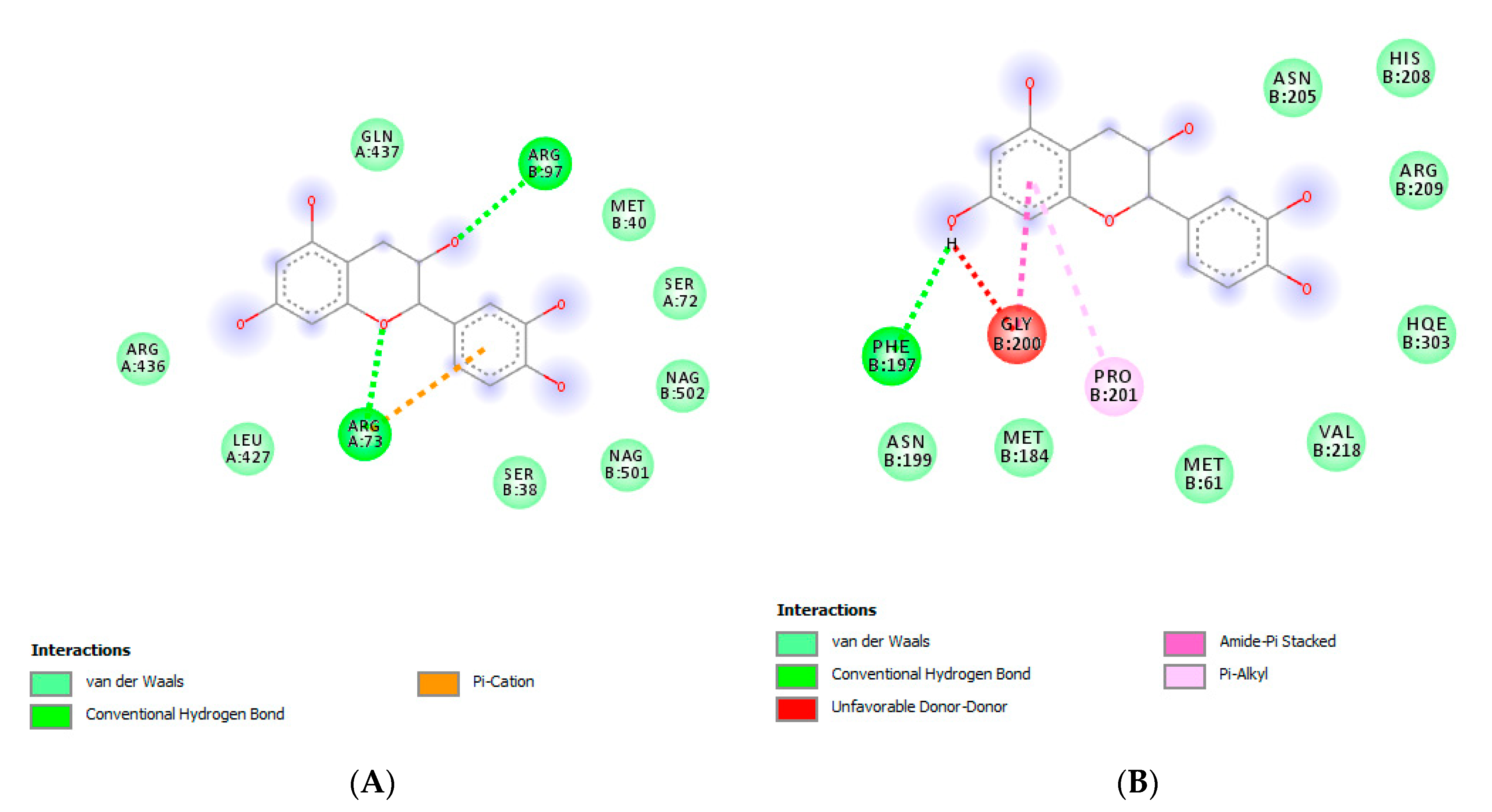
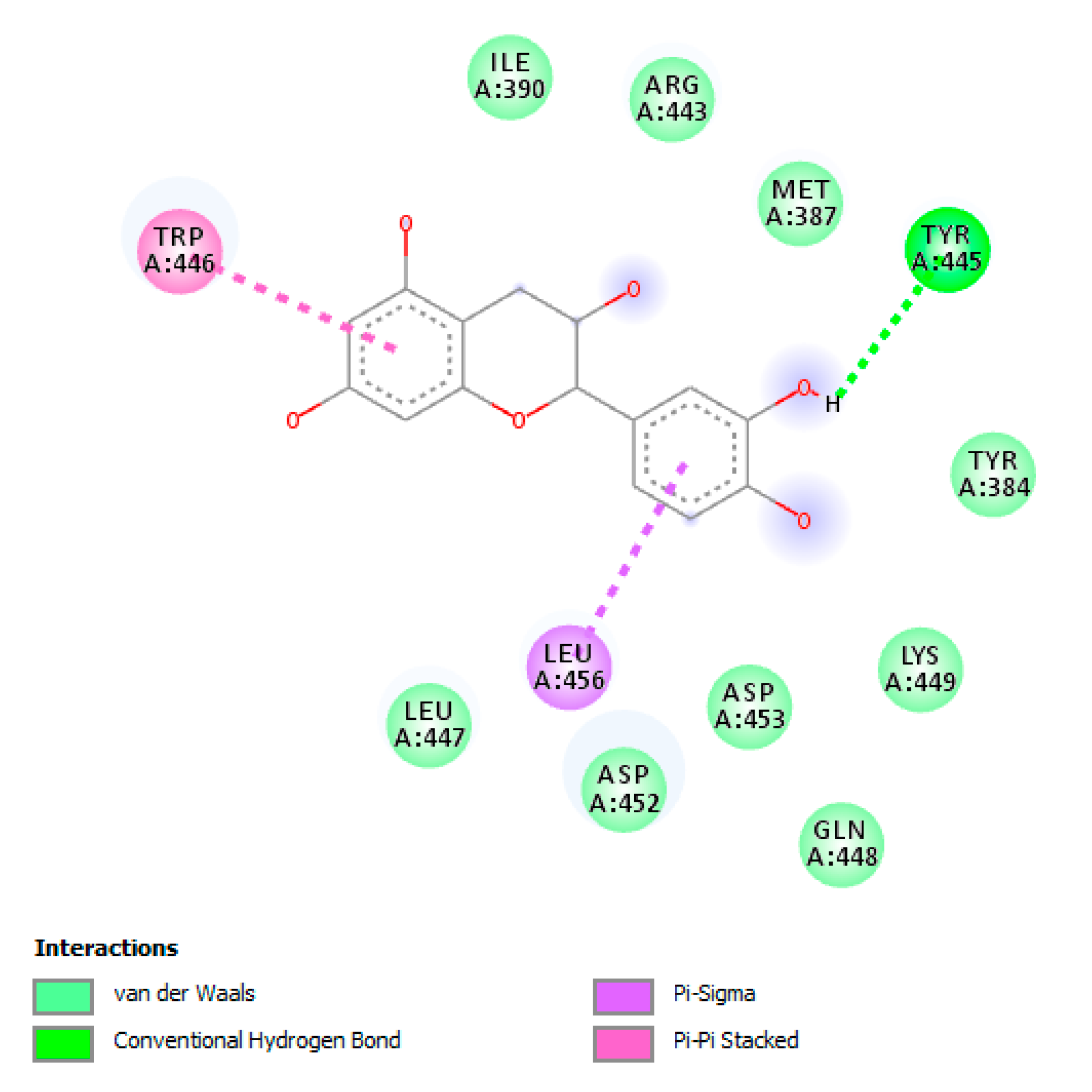
3.7. Bioinformatics Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Angelini, P.; Arcangeli, A.; Bistocchi, G.; Rubini, A.; Venanzoni, R.; Perini, C. Current knowledge of Umbrian macrofungi (central Italy). Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2017, 151, 915–923. [Google Scholar] [CrossRef]
- Angelini, P.; Bistocchi, G.; Arcangeli, A.; Rubini, A.; Venanzoni, R. Inventory, diversity and communities of macrofungi in the Collestrada forest (Umbria, central Italy). Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2016, 150, 1096–1105. [Google Scholar] [CrossRef]
- Angelini, P.; Pagiotti, R.; Granetti, B. Effect of antimicrobial activity of Melaleuca alternifolia essential oil on antagonistic potential of Pleurotus species against Trichoderma harzianum in dual culture. World J. Microbiol. Biotechnol. 2008, 24, 197–202. [Google Scholar] [CrossRef]
- Angelini, P.; Pagiotti, R.; Venanzoni, R.; Granetti, B. Antifungal and allelopathic effects of Asafoetida against Trichoderma harzianum and Pleurotus spp. Allelopath. J. 2009, 23, 357–368. [Google Scholar]
- Pagiotti, R.; Angelini, P.; Rubini, A.; Tirillini, B.; Granetti, B.; Venanzoni, R. Identification and characterisation of human pathogenic filamentous fungi and susceptibility to Thymus schimperi essential oil. Mycoses 2011, 54, e364–e376. [Google Scholar] [CrossRef] [PubMed]
- Perotto, S.; Angelini, P.; Bianciotto, V.; Bonfante, P.; Girlanda, M.; Kull, T.; Mello, A.; Pecoraro, L.; Perini, C.; Persiani, A. Interactions of fungi with other organisms. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2013, 147, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Picco, A.; Angelini, P.; Ciccarone, C.; Franceschini, A.; Ragazzi, A.; Rodolfi, M.; Varese, G.; Zotti, M. Biodiversity of emerging pathogenic and invasive fungi in plants, animals and humans in Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2011, 145, 988–996. [Google Scholar] [CrossRef]
- Feeney, M.J.; Miller, A.M.; Roupas, P. Mushrooms—Biologically distinct and nutritionally unique: Exploring a “third food kingdom”. Nutr. Today 2014, 49, 301. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.; Akavia, E. Regulatory Issues of Mushrooms as Functional Foods and Dietary Supplements: Safety and Efficacy. In Mushrooms as Functional Foods; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 199–226. [Google Scholar]
- Angelini, P.; Tirillini, B.; Bistocchi, G.; Arcangeli, A.; Rubini, A.; Pellegrino, R.M.; Fabiani, R.; Cruciani, G.; Venanzoni, R.; Rosignoli, P. Overview of the Biological Activities of the Methanolic Extract from wild Fomitopsis pinicola Fruiting Body from Central Italy. Int. J. Med. Mushroom 2018, 20, 1047–1063. [Google Scholar]
- Badalyan, S.M.; Borhani, A. Medicinal, Nutritional, and Cosmetic Values of Macrofungi Distributed in Mazandaran Province of Northern Iran. Int. J. Med. Mushrooms 2019, 21, 1099–1106. [Google Scholar] [CrossRef]
- Covino, S.; D’Ellena, E.; Tirillini, B.; Angeles, G.; Arcangeli, A.; Bistocchi, G.; Venanzoni, R.; Angelini, P. Characterization of Biological Activities of Methanol Extract of Fuscoporia torulosa (Basidiomycetes) from Italy. Int. J. Med. Mushrooms 2019, 21, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- CFR Ferreira, I.; A Vaz, J.; Vasconcelos, M.H.; Martins, A. Compounds from wild mushrooms with antitumor potential. Anti-Cancer Agents Med. Chem. 2010, 10, 424–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyigo, V.; Baraza, L.; Nkunya, M.; Mdachi, S.; Joseph, C.; Waziri, A. Chemical constitutents and cytotoxicity of some Tanzanian wild mushrooms. Tanzan. J. Sci. 2005, 31, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Suseem, S.; Saral, A.M. Analysis on essential fatty acid esters of mushroom pleurotus eous and its antibacterial activity. Asian J. Pharm. Clin. Res. 2013, 6, 188–191. [Google Scholar]
- Russo, A.; Piovano, M.; Clericuzio, M.; Lombardo, L.; Tabasso, S.; Chamy, M.; Vidari, G.; Cardile, V.; Vita-Finzi, P.; Garbarino, J. Putrescine-1, 4-dicinnamide from Pholiota spumosa (Basidiomycetes) inhibits cell growth of human prostate cancer cells. Phytomedicine 2007, 14, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef]
- Voss-Rech, D.; Potter, L.; Vaz, C.S.L.; Pereira, D.I.B.; Sangioni, L.A.; Vargas, A.C.; de Avila Botton, S. Antimicrobial resistance in nontyphoidal Salmonella isolated from human and poultry-related samples in Brazil: 20-year meta-analysis. Foodborne Pathog. Dis. 2017, 14, 116–124. [Google Scholar] [CrossRef]
- Rouzaud, C.; Hay, R.; Chosidow, O.; Dupin, N.; Puel, A.; Lortholary, O.; Lanternier, F. Severe dermatophytosis and acquired or innate immunodeficiency: A review. J. Fungi 2016, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Kotra, L.; Mobashery, S. β-Lactam antibiotics, β-lactamases and bacterial resistance. Bull. Institut Pasteur 1998, 96, 139–150. [Google Scholar] [CrossRef]
- Morschhäuser, J.; Köhler, G.; Ziebuhr, W.; Blum-Oehler, G.; Dobrindt, U.; Hacker, J. The Activities Of Bacterial Pathogens In Vivo: Based on Contributions to a Royal Society Discussion Meeting. In Evolution of Microbial Pathogens; World Scientific: Singapore, 2000; pp. 349–373. [Google Scholar]
- Mukherjee, M.; Horwitz, B.A.; Sherkhane, P.D.; Hadar, R.; Mukherjee, P.K. A secondary metabolite biosynthesis cluster in Trichoderma virens: Evidence from analysis of genes underexpressed in a mutant defective in morphogenesis and antibiotic production. Curr. Genet. 2006, 50, 193–202. [Google Scholar] [CrossRef]
- Sandven, P. Epidemiology of candidemia. Rev. Iberoam. Micol. 2000, 17, 73–81. [Google Scholar] [PubMed]
- Thompson, J.D. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüffler, A. Secondary metabolites of basidiomycetes. In Physiology and Genetics; Springer: Berlin/Heidelberg, Germany, 2018; pp. 231–275. [Google Scholar]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef]
- Jose Alves, M.; CFR Ferreira, I.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A Review on Antifungal Activity of Mushroom (Basidiomycetes) Extracts and Isolated Compounds. Curr. Top. Med. Chem. 2013, 13, 2648–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, P.; Arcangeli, A.; Bistocchi, G.; Venanzoni, R.; Rubini, A. Tricholosporum goniospermum, genetic diversity and phylogenetic relationship with the Tricholomatineae [formerly tricholomatoid clade]. Sydowia 2017, 69, 9–18. [Google Scholar]
- Dej-adisai, S.; Meechai, I.; Puripattanavong, J.; Kummee, S. Antityrosinase and antimicrobial activities from Thai medicinal plants. Arch. Pharmacal. Res. 2014, 37, 473–483. [Google Scholar] [CrossRef]
- Hoekstra, E.; Aptroot, A. CBS Course of Mycology; Centrarlbureau Voor Schimmel Cultures: Baarn/Delft, The Netherlands, 1998. [Google Scholar]
- Angelini, P.; Girometta, C.; Tirillini, B.; Moretti, S.; Covino, S.; Cipriani, M.; D’Ellena, E.; Angeles, G.; Federici, E.; Savino, E. A comparative study of the antimicrobial and antioxidant activities of Inonotus hispidus fruit and their mycelia extracts. Int. J. Food Prop. 2019, 22, 768–783. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Sinan, K.I.; Chiavaroli, A.; Orlando, G.; Bene, K.; Zengin, G.; Cziáky, Z.; Jekő, J.; Mahomoodally, M.F.; Picot-Allain, M.C.N.; Menghini, L.; et al. Evaluation of Pharmacological and Phytochemical Profiles Piptadeniastrum africanum (Hook. f.) Brenan Stem Bark Extracts. Biomolecules 2020, 10, 516. [Google Scholar] [CrossRef] [Green Version]
- Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Carradori, S.; Di Simone, S.; Ciferri, M.C.; Zengin, G.; Ak, G.; et al. Water Extract from Inflorescences of Industrial Hemp Futura 75 Variety as a Source of Anti-Inflammatory, Anti-Proliferative and Antimycotic Agents: Results from In Silico, In Vitro and Ex Vivo Studies. Antioxidants 2020, 9, 437. [Google Scholar] [CrossRef]
- Chiavaroli, A.; Sinan, K.I.; Zengin, G.; Mahomoodally, M.F.; Sadeer, N.B.; Etienne, O.K.; Cziáky, Z.; Jekő, J.; Glamocilja, J.; Sokovic, M.; et al. Identification of Chemical Profiles and Biological Properties of Rhizophora racemosa G. Mey. Extracts Obtained by Different Methods and Solvents. Antioxidants 2020, 9, 533. [Google Scholar] [CrossRef]
- Rodrıguez-Delgado, M.; Malovana, S.; Perez, J.; Borges, T.; Montelongo, F.G. Separation of phenolic compounds by high-performance liquid chromatography with absorbance and fluorimetric detection. J. Chromatogr. A 2001, 912, 249–257. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
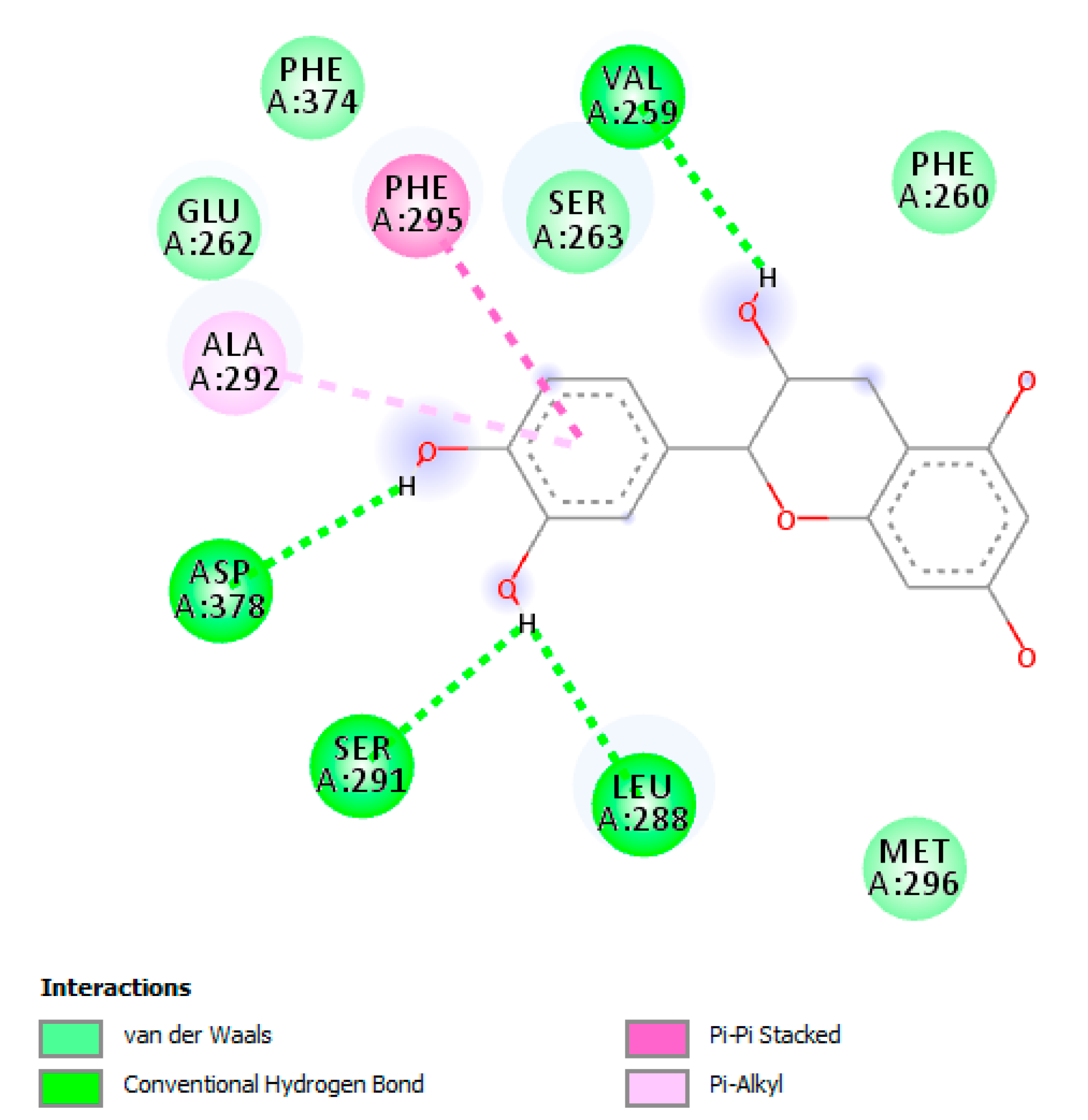{kind=link}
{kind=link}
| T. goniospermum Parts | Solvents | Gallic Acid | Catechin | Epicatechin | Resveratrol |
|---|---|---|---|---|---|
| µg/mL | µg/mL | µg/mL | µg/mL | ||
| n-hex | 0.96 ± 0.5 | 0.00 | 0.00 | 0.00 | |
| Fruiting bodies | EA | 0.09 ± 0.01 | 0.03 ± 0.01 | 0.00 | 0.00 |
| MeOH | 0.65 ± 0.03 | 0.00 | 0.00 | 0.00 | |
| n-hex | 0.95 ± 0.3 | 0.00 | 0.00 | 0.00 | |
| Mycelia | EA | 2.18 ± 0.11 | 0.00 | 0.00 | 0.00 |
| MeOH | 3.95 ± 0.13 | 0.00 | 0.00 | 0.00 |
| T. goniospermum Parts | Solvents | TPC | DPPH | CUPRAC | FRAP |
|---|---|---|---|---|---|
| (mg GAE/g) | (mg TE/g) | (mg TE/g) | (mg TE/g) | ||
| n-hex | 10.91 ± 0.56 | 9.35 ± 0.16 | 53.92 ± 1.31 | 14.86 ± 1.21 | |
| Fruiting bodies | EA | 70.51 ± 0.06 | 88.82 ± 1.47 | 307.71 ± 3.83 | 134.06 ± 1.50 |
| MeOH | 14.87 ± 0.79 | 17.69 ± 0.95 | 131.52 ± 0.67 | 20.54 ± 0.77 | |
| n-hex | 9.63 ± 0.22 | 7.53 ± 0.70 | 54.35 ± 0.92 | 15.15 ± 1.62 | |
| Mycelia | EA | 33.74 ± 0.80 | 29.93 ± 3.54 | 155.31 ± 1.85 | 74.26 ± 1.79 |
| MeOH | 7.39 ± 0.14 | 7.74 ± 0.69 | 129.60 ± 0.56 | 12.94 ± 1.33 |
| T. goniospermum Parts | Solvents | AChE Inhibition | BChE Inhibition | Tyrosinase Inhibition | Amylase Inhibition |
|---|---|---|---|---|---|
| (mg GALAE/g) | (mg GALAE/g) | (mg KAE) | (mmol ACAE/g) | ||
| n-hex | NA | 5.48 ± 0.03 | 83.80 ± 1.45 | 0.55 ± 0.01 | |
| Fruiting bodies | EA | NA | 26.78 ± 0.21 | 554.30 ± 9.41 | 2.97 ± 0.10 |
| MeOH | NA | 5.07 ± 0.02 | 48.48 ± 0.07 | 0.21 ± 0.01 | |
| n-hex | NA | 9.14 ± 0.07 | 127.76 ± 0.73 | 0.73 ± 0.02 | |
| Mycelia | EA | NA | 25.21 ± 0.24 | 412.81 ± 1.39 | 2.25 ± 0.07 |
| MeOH | NA | 2.61 ± 0.01 | 28.17 ± 0.39 | 0.17 ± 0.01 |
| Bacterial Strain (ID) | Extract Typology | Minimum Inhibitory Concentration (MIC) | |||
|---|---|---|---|---|---|
| n-hex (mg mL−1) * | EA (mg mL−1) * | MeOH (mg mL−1) * | Ciprofloxacin (µg mL−1) ** | ||
| Gram− | |||||
| E. coli (ATCC 10536) | mycelia | 0.157 (0.125–0.25) | 0.099 (0.0625–0.125) | 0.099 (0.0625–0.125) | <0.12 |
| fruiting bodies | 0.198 (0.125–0.25) | 0.157 (0.125–0.25) | 0.198 (0.125–0.250) | ||
| E. coli (PeruMycA 2) | mycelia | 0.315 (0.25–0.5) | 0.157 (0.125–025) | 0.198 (0.125–0.25) | 1.23 (1.95–0.98) |
| fruiting bodies | 0.396 (0.25–0.5) | 0.315 (0.25–0.5) | 0.315 (0.25–0.5) | ||
| E. coli (PeruMycA 3) | mycelia | 0.396 (0.25–0.5) | 0.157 (0.125–0.250) | 0.198 (0.125–0.250) | 0.62 (0.98–0.49) |
| fruiting bodies | 0.315 (0.125–0.25) | 0.198 (0.125–0.250) | 0.315 (0.25–0.5) | ||
| P. aeruginosa (PeruMyc 5) | mycelia | 0.63 (0.5–1) | 0.315 (0.25–0.5) | 0.396 (0.25–0.5) | 1.23 (1.95–0.98) |
| fruiting bodies | 0.79 (0.05–1) | 0.396 (0.25–0.5) | 0.63 (0.5–1) | ||
| S. typhy (PeruMyc 7) | mycelia | 0.79 (0.5–1) | 0.315 (0.25–0.5) | 0.62 (0.5–1) | 0.38 (0.49–0.24) |
| fruiting bodies | >1 | 0.62 (0.5–1) | 0.79 (0.5–1) | ||
| Gram+ | |||||
| B. cereus (PeruMycA 4) | mycelia | 0.198 (0.125–0.5) | 0.099 (0.0625–0.125) | 0.157 (0.125–0.250) | <0.12 |
| fruiting bodies | 0.314 (0.25–0.5) | 0.157 (0.125–0.250) | 0.099 (0.0625–0.125) | ||
| B. subtilis (PeruMyc 6) | mycelia | 0.315 (0.125–0.25) | 0.078 (0.062–0.125) | 0.198 (0.125–0.25) | <0.12 |
| fruiting bodies | 0.396 (0.25–0.5) | 0.157 (0.125–0.25) | 0.314 (0.25–0.5) | ||
| S. aureus (ATCC 6538) | mycelia | 0.396 (0.25–0.5) | 0.157 (0.125–0.25) | 0.198 (0.125–0.25) | 0.62 (0.98–0.49) |
| fruiting bodies | 0.63 (0.5–1) | 0.198 (0.125–0.25) | 0.315 (0.25–0.5) | ||
| Yeasts (ID) | Extract Typology | Minimum Inhibitory Concentration (MIC) | |||
|---|---|---|---|---|---|
| n-hex (mg mL−1) * | EA (mg mL−1) * | MeOH (mg mL−1) * | Fluconazole (µg mL−1) ** | ||
| C. albicans (YEPGA 6183) | mycelia | 0.314 (0.25–0.5) | 0.099 (0.0625–0.150) | 0.157 (0.125–0.250) | 2 |
| fruiting bodies | 0.396 (0.25–0.5) | 0.198 (0.125–0.250) | 0.314 (0.25–0.5) | ||
| C. tropicalis (YEPGA 6184) | mycelia | 0.157 (0.125–0.25) | 0.099 (0.0625–0.125) | 0.099 (0.0625–0.125) | 2 |
| fruiting bodies | 0.198 (0.125–0.25) | 0.157 (0.125–0.250) | 0.198 (0.125–0.250) | ||
| C. albicans (YEPGA 6379) | mycelia | 0.099 (0.0625–0.125) | 0.051 (0.031–0.0625) | 0.099 (0.0625–0.125) | 1 |
| fruiting bodies | 0.157 (0.125–0.25) | 0.198 (0.125–0.250) | 0.198 (0.125–0.250) | ||
| C. parapsilopsis (YEPGA 6551) | mycelia | 0.314 (0.25–0.5) | 0.079 (0.125–0.0625) | 0.157 (0.125–0.25) | 4 |
| fruiting bodies | 0.396 (0.25–0.5) | 0.099 (0.0625–0.125) | 0.198 (0.125–0.250) | ||
| Dermatophytes (ID Strain) | Extract Typology | Minimum Inhibitory Concentration (MIC) | |||
|---|---|---|---|---|---|
| n-hex (mg mL−1) * | EA (mg mL−1) * | MeOH (mg mL−1) * | Griseofulvin (µg mL−1) ** | ||
| A. crocatum (CCF 5300) | mycelia | 0.25 (0.125–0.25) | 0.084 (0.031–0.062) | 0.157 (0.125–0.25) | >8 |
| fruiting bodies | 0.314 (0.25–0.5) | 0.46 (0.62–0.125) | 0.198 (0.125–0.25) | ||
| A. curreyi (CCF 5207) | mycelia | 0.794 (0.5–1) | 0.157 (0.125–0.25) | 0.363 (0.62–0.125) | >8 |
| fruiting bodies | >1 | 0.315 (0.25–0.5) | 0.39 (0.031–0.62) | ||
| A. gypseum (CCF 6261) | mycelia | 0.397 (0.25–0.5) | 0.099 (0.62–0.125) | 0.049 (0.031–0.062) | 1.587 (1–2) |
| fruiting bodies | 0.397 (0.25–0.5) | 0.181 (0.031–0.062) | 0.25 (0.125–0.5) | ||
| A. insingulare (CCF 5417) | mycelia | 0.794 (0.5–1) | 0.194 (0.125–0.5) | 0.794 (0.5–1) | >8 |
| fruiting bodies | >1 | 0.623 (0.5–1) | >1 | ||
| A. quadrifidum (CCF 5792) | mycelia | 0.623 (0.5–1) | 0.198 (0.125–0.25) | 0.315 (0.25–0.5) | >8 |
| fruiting bodies | >1 | 0.623 (0.5–1) | 0.794 (0.5–1) | ||
| T. mentagrophytes (CCF 4823) | mycelia | 0.315 (0.25–0.5) | 0.157 (0.125–0.25) | 0.315 (0.25–0.5) | 2.52 (2–4) |
| fruiting bodies | 0.623 (0.5–1) | 0.194 (0.125–0.25) | 0.397 (0.5–1) | ||
| T. mentagrophytes (CCF 5930) | mycelia | 0.794 (0.5–1) | 0.57 (0.125–0.25) | 0.63 (0.5–1) | 3.174 (2–4) |
| fruiting bodies | >1 | 0.198 (0.125–0.25) | >1 | ||
| T. rubrum (CCF 4933) | mycelia | 0.397 (0.25–0.5) | 0.049 (0.031–062) | 0.039 (0.031–0.062) | 1.26 (1–2) |
| fruiting bodies | 0.794 (0.5–1) | 0.198 (0.125–0.25) | 0.315 (0.25–0.5) | ||
| T. rubrum (CCF 4879) | mycelia | 0.397 (0.25–0.5) | 0.049 (0.031–0.062) | 0.315 (0.25–0.5) | 3.175 (2–4) |
| fruiting bodies | 0.794 (0.5–1) | 0.157 (0.125–0.25) | 0.623 (0.5–1) | ||
| T. tonsurans (CCF 4834) | mycelia | 0.397 (0.25–0.5) | 0.039 (0.031–0.062) | 0.099 (0.062–0.125) | 0.198 (0.125–0.25) |
| fruiting bodies | 0.623 (0.5–1) | 0.314 (0.25–0.5) | 0.397 (0.25–0.5) | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Tirillini, B.; Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Di Simone, S.C.; et al. Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom. Antibiotics 2020, 9, 513. https://doi.org/10.3390/antibiotics9080513
Angelini P, Venanzoni R, Angeles Flores G, Tirillini B, Orlando G, Recinella L, Chiavaroli A, Brunetti L, Leone S, Di Simone SC, et al. Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom. Antibiotics. 2020; 9(8):513. https://doi.org/10.3390/antibiotics9080513
Chicago/Turabian StyleAngelini, Paola, Roberto Venanzoni, Giancarlo Angeles Flores, Bruno Tirillini, Giustino Orlando, Lucia Recinella, Annalisa Chiavaroli, Luigi Brunetti, Sheila Leone, Simonetta Cristina Di Simone, and et al. 2020. "Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom" Antibiotics 9, no. 8: 513. https://doi.org/10.3390/antibiotics9080513
APA StyleAngelini, P., Venanzoni, R., Angeles Flores, G., Tirillini, B., Orlando, G., Recinella, L., Chiavaroli, A., Brunetti, L., Leone, S., Di Simone, S. C., Ciferri, M. C., Zengin, G., Ak, G., Menghini, L., & Ferrante, C. (2020). Evaluation of Antioxidant, Antimicrobial and Tyrosinase Inhibitory Activities of Extracts from Tricholosporum goniospermum, an Edible Wild Mushroom. Antibiotics, 9(8), 513. https://doi.org/10.3390/antibiotics9080513