1. Introduction
Tissue abnormalities and changes in regional loading or maladaptation of myocardial tissue can contribute to the growth and remodeling of the heart. Long-term effects such as this can lead to apoptosis and diseases such as cardiac hypertrophy. Concomitantly, soft tissues, including cardiac tissue, are heterogeneous, nearly incompressible, and anisotropic, where the anisotropy arises from the underlying muscle fibers. Such tissues have complex geometries, and often are subject to large deformations [
1,
2]. Therefore, analyzing mechanical behavior and deformations of anisotropic tissue is vital and challenging due to its complex nature.
Indentation is a widely used technique in biomechanics to analyze the mechanical behavior and properties of soft tissue [
3,
4]. Due to the complex nature of the mechanical problems arising in traditional indentation tests on soft tissues, numerical techniques can provide an alternative way of analyzing the mechanical behavior of soft tissues on both cellular and regional scales [
5]. They also can predict what mechanotransduction occurs during indentation: how the indenter pressing on the tissue causes it to grow and remodel.
This paper presents a finite difference analysis of indentation of anisotropic tissue using the mechanical bidomain model, a macroscopic mathematical model based on the microscopic properties of the tissue. As an example, we take the tissue to be cardiac muscle, but we do not stress specific parameter values because the model should apply qualitatively to a variety of soft tissues (e.g., cartilage).
The mechanical bidomain model accounts for the fiber-based anisotropy in both intra- and extracellular spaces and assumes that each can have different degrees of anisotropy (unequal anisotropy ratios). In 2019, a 2D mechanical bidomain model was implemented that included these properties of anisotropy [
6]. In our derivation of the 3D mechanical bidomain model, we extended the equations of the 2D model with anisotropic properties.
Another reason for choosing this study of tissue indentation is that it is analogous to unipolar electrical stimulation of cardiac tissue, where current is injected through a small electrode. In 1989, Sepulveda et al. used the electrical bidomain model to show that unequal anisotropy ratios are critical to understanding unipolar stimulation, and the insights gained from that study carried over to analyzing pacing and defibrillation of the heart [
7].
Over the past decade, the mechanical bidomain model was developed as a two-dimensional (2D) model and it was able to successfully address unresolved questions in the growth and remodeling of soft tissue [
8,
9]. The most significant difference between the mechanical bidomain model and other monodomain models that are currently available, is that the mechanical bidomain model can distinguish the mechanical properties in the intra- and extracellular spaces of cardiac tissue. Our hypothesis is that differences in the intra- and extracellular displacements activate integrins and thereby drive mechanotransduction. In this paper we developed a three-dimensional numerical implementation of the mechanical bidomain model to simulate tissue indentation.
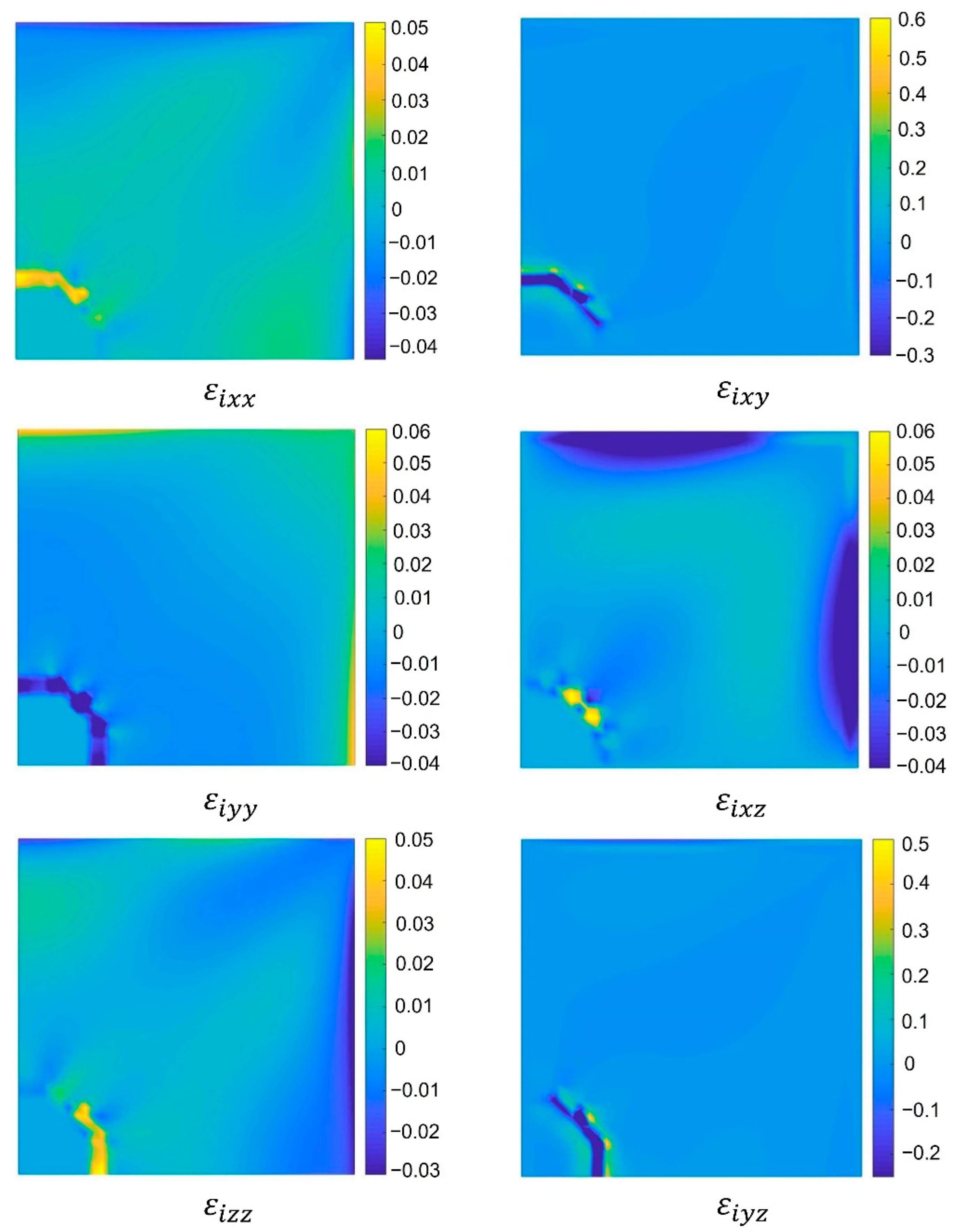
4. Discussion
A 3D version of the mechanical bidomain model was derived using vector potentials rather than the stream functions that are used for the 2D mechanical bidomain model [
6,
10,
11,
16]. Additionally, we defined the terms
and
which helped to distinguish the monodomain and bidomain behaviors. The magnitude of bidomain displacement,
, was important, since it represents the bidomain mechanotransduction effects in the mechanical bidomain model. In the mechanical bidomain model, effects of the direction of
on mechanotransduction is an open question. This study quantitatively analyzed the magnitude of the bidomain displacement,
.
We used the finite difference method for the numerical analysis of the tissue indentation using the 3D mechanical bidomain model with anisotropy. Using a finite difference method for this analysis had both advantages and disadvantages. The finite difference method resulted in less complicated computational code with shorter execution time compared with using the finite element method. One major disadvantage of using the finite difference method was that we could not easily implement a variable grid spacing. A finite element approach would easily allow a smaller grid spacing near the indenter. It would also allow us to analyze irregularly shaped tissues, rather than the tissue cube we studied. Additionally, by using symmetry conditions we were able to analyze one octant of the cube, which made calculations computationally less demanding but limited the tissue geometry that we could study.
We tested the tissue by applying different boundary conditions. When free boundary conditions were applied to the outer edges of X and Z directions, the calculation became unstable and the overrelaxation routine would give large oscillations rather than converging to a solution. At large distances, the displacements of intra- and extracellular spaces and should go to zero regardless of outer boundary conditions. Therefore, we believed that zero displacement boundary conditions were reasonable for a large tissue. However, we had limitations for the size of the cube due to computational constraints.
The mechanical bidomain model contained pressure terms in the intracellular and extracellular spaces,
and
, which arose because the tissue was incompressible. We did not calculate
and
in our calculations, because we were focused on mechanotransduction driven by
. We could easily have removed the pressure terms from the basic equations governing equilibrium in the tissue, Equations (7) and (8), but had a more difficult time removing the pressure from a free boundary condition (zero stress). In that case these pressure terms were removed by taking derivatives of the stress in the directions perpendicular to the surface normal, a technique explained by William H. Mitchell [
17]. It would be interesting to extend our calculations to compute the pressures during tissue indentation.
We assumed linear and small strains and incompressible spaces in cardiac tissue with straight fibers. The mechanical bidomain model can also be derived for compressible spaces [
18] and tissue with curving fibers [
6,
11].
When both intra- and extracellular spaces pressed downward, the bidomain displacement was zero in isotropic tissue and tissue with equal anisotropy ratios. However, for the tissue with unequal anisotropy ratios, the magnitude of the bidomain displacement was nonzero (
Figure 7). This is because Equations (20) and (21) uncouple for isotropic and equal anisotropic tissues, but not for tissue with unequal anisotropy ratios. In 2019, Wijesinghe and Roth observed the effects of anisotropy ratios in a 2D tissue with curving fibers. They observed nonzero bidomain displacement only in tissue with unequal anisotropy ratios [
6].
The issue of the appropriate boundary condition is subtle. In a study of shearing a slab of tissue [
19], it was assumed that the extracellular space was being pulled, not the intracellular space. The intracellular space would also shear because it was pulled by the extracellular space, but this pulling was through the integrins coupling the two spaces, so it caused mechanotransduction at the surface. Pulling only on the extracellular space is easy to imagine (picture the tissue pulled by tiny ropes attached to the extracellular matrix). However, when pushing on tissue, it is more difficult to picture how the intracellular space could be free. Ultimately, the correct boundary condition would need to be determined by either experiment or by a microscopic mechanical model.
We predict tissue growth as an effect of a localized force using an in vitro tissue culture experiment. This prediction needs to be tested. By growing a 3D tissue cube, pressing it down and observing over a long time, we would be able to observe how tissue grows or remodels around the area affected by the force.
We assumed that the mechanical bidomain model has unequal anisotropy ratios. Analogously, the electrical bidomain is known to have unequal anisotropy ratios. Our calculations indicate that the condition of unequal anisotropy ratios can have a dramatic effect on the distribution of mechanotransduction, but this effect is sensitive to the exact boundary condition used at the surface. This result highlights the importance of the fibrous structure of the tissue.
In comparison with the conclusions of the study conducted by Sepulveda et al., on unipolar electrical stimulation of cardiac tissue [
7], our results from this study indicate that unequal anisotropy ratios can likewise affect a localized mechanical force, as in indentation. This calculation using the mechanical bidomain model is, therefore, the mechanical analogue of the Sepulveda et al.’s calculation using the electrical bidomain model. The implications of our calculation may extend beyond indentation experiments and impact any situation in which there is a localized force acting on the tissue.
In 2018, analytical expressions were derived predicting how the mechanical bidomain model responds to a localized force caused by a point source in the extracellular space [
20]. This calculation, however, was for isotropic tissue. We can develop an experiment to test this phenomenon by taking a magnetic-particle based technique which is being used to activate mechanosensitive ion channels, as an example. This method allows the direct application of a highly localized mechanical force to a selected region of a mechanosensitive ion channel [
21]. By inserting a superparamagnetic nanoparticle into the extracellular space, a localized force can be applied on the nanoparticle, using magnetic tweezers. This can make the nanoparticle pull the extracellular matrix, causing the integrins to stretch. The stretching of integrins in the membrane can trigger mechanotransduction [
20]. The advantage of our calculation is that we could examine the influence of unequal anisotropy ratios on such an experiment.
The mechanical bidomain model was derived based on the hypothesis that the difference of the displacement of the intracellular and extracellular spaces drives the mechanotransduction. Most of the other monodomain models assume that stresses or strains cause the mechanotransduction. Macro and micro tissue indentation is a widely used technique in analyzing mechanical properties using monodomain models. From our analysis, we were able to introduce a different perspective: that tissue indentation can also affect bidomain properties. The results of this study show that the distribution of monodomain strain and distribution of bidomain displacement are qualitatively different. Therefore, to analyze the growth and remodeling of cardiac tissue, it is important to consider the forces acting on membrane integrins. The clinical significance of our results are still being determined, but our results suggest that any localized force applied to the surface of a tissue will induce growth and remodeling in a characteristic pattern around the location of the applied force.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







