Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators

 ,
,  , , ,
, , ,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field and Experimental Conditions
2.2. Analyses of Soil Microbial Diversity
2.3. Enzymatic Activities and Soil Alteration Index
2.4. Aggregate-Sized C and N Fractions
2.5. Statistical Analyses
3. Results
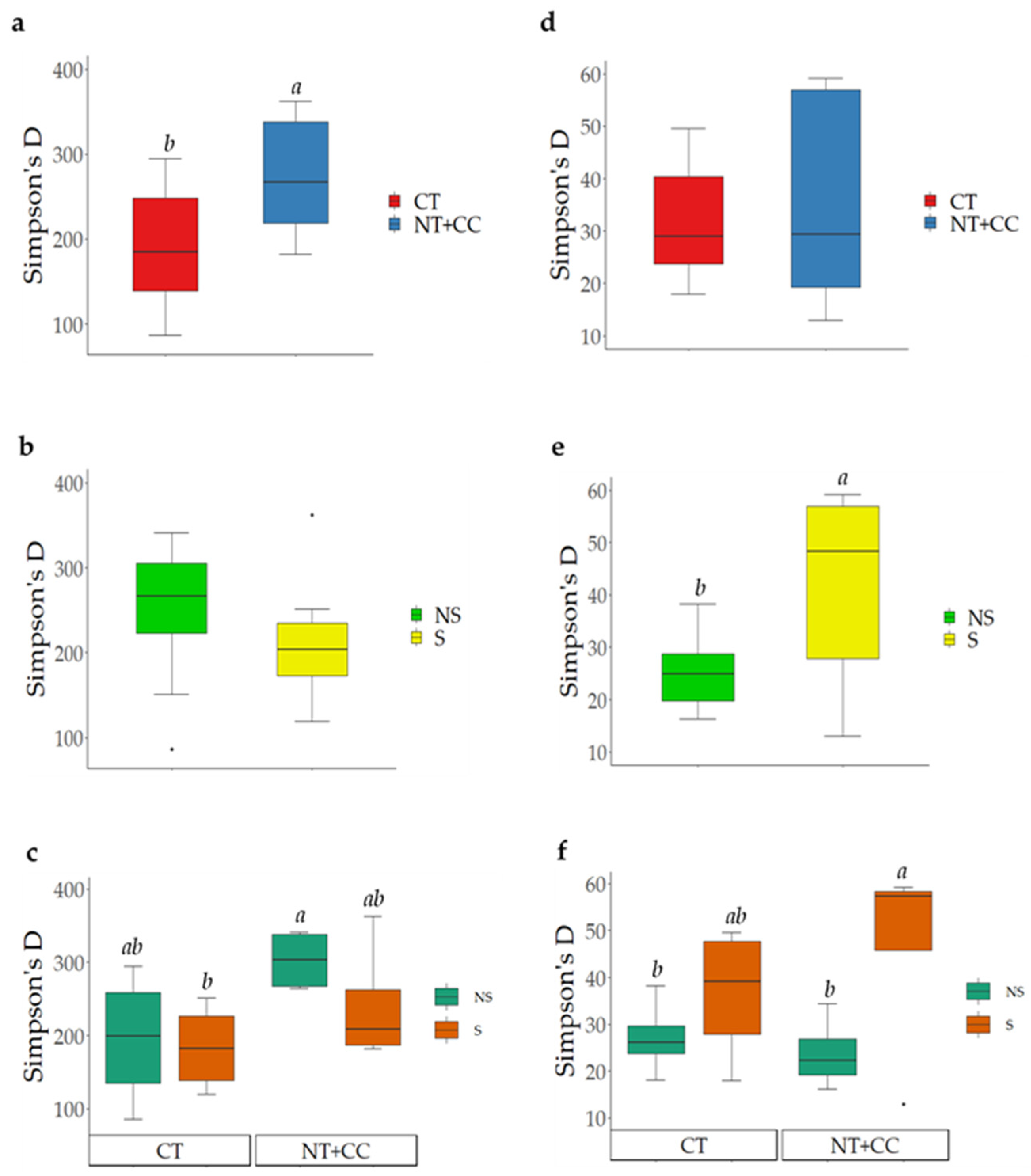
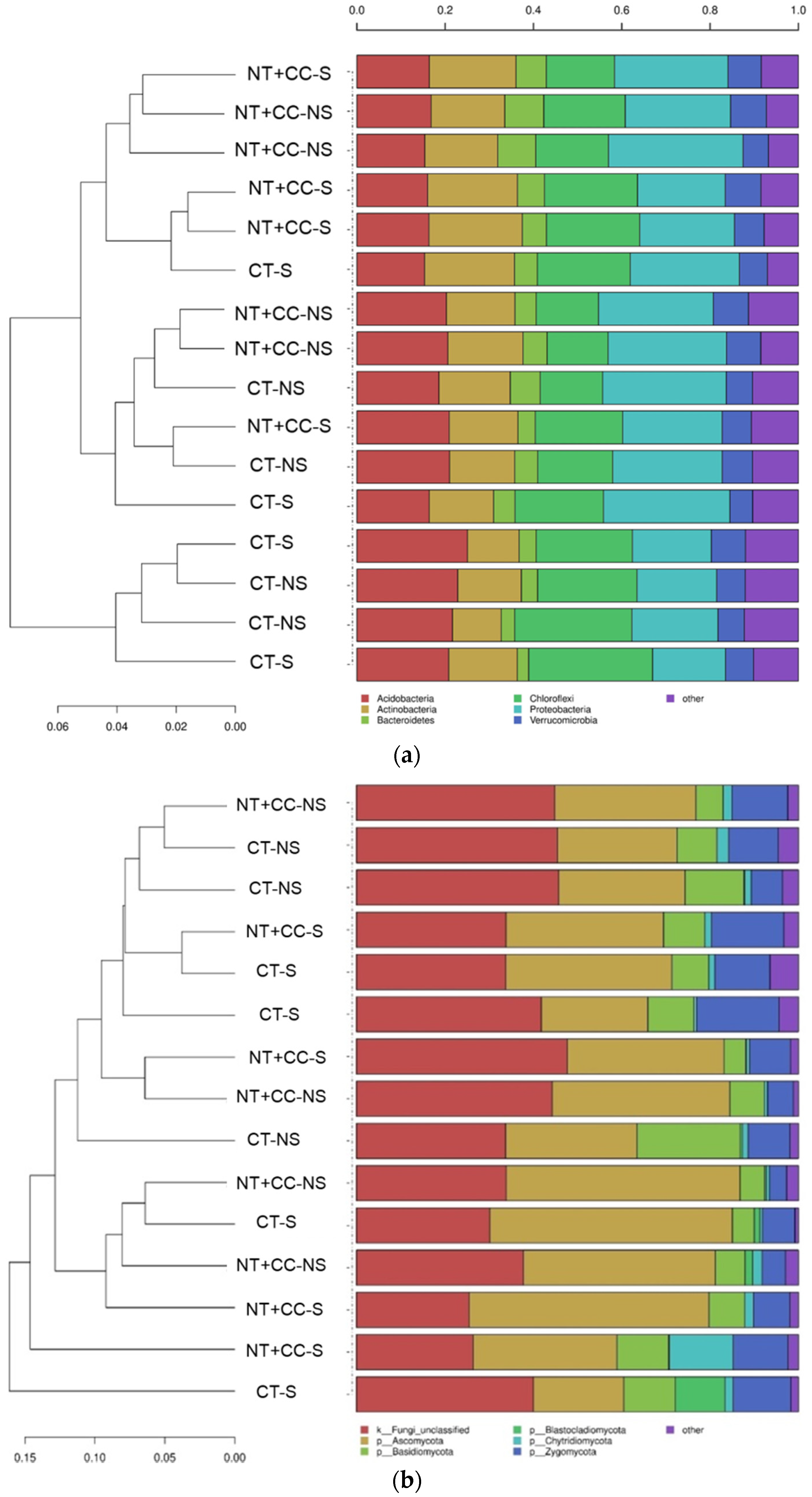
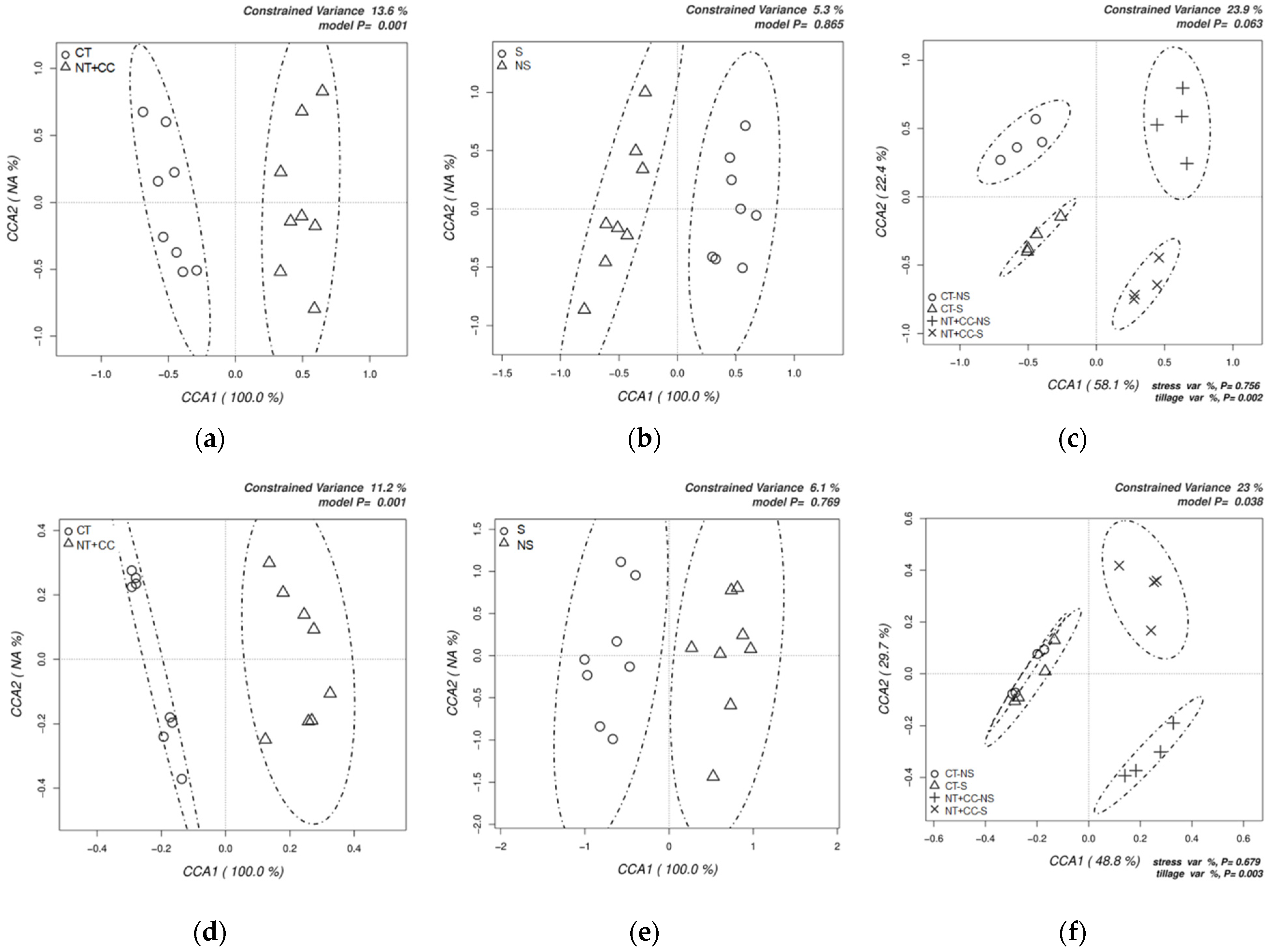
3.1. Analyses of Soil Microbial Diversity
3.2. Enzymatic Activities and Soil Alteration Index
3.3. Aggregate-Associated C and N
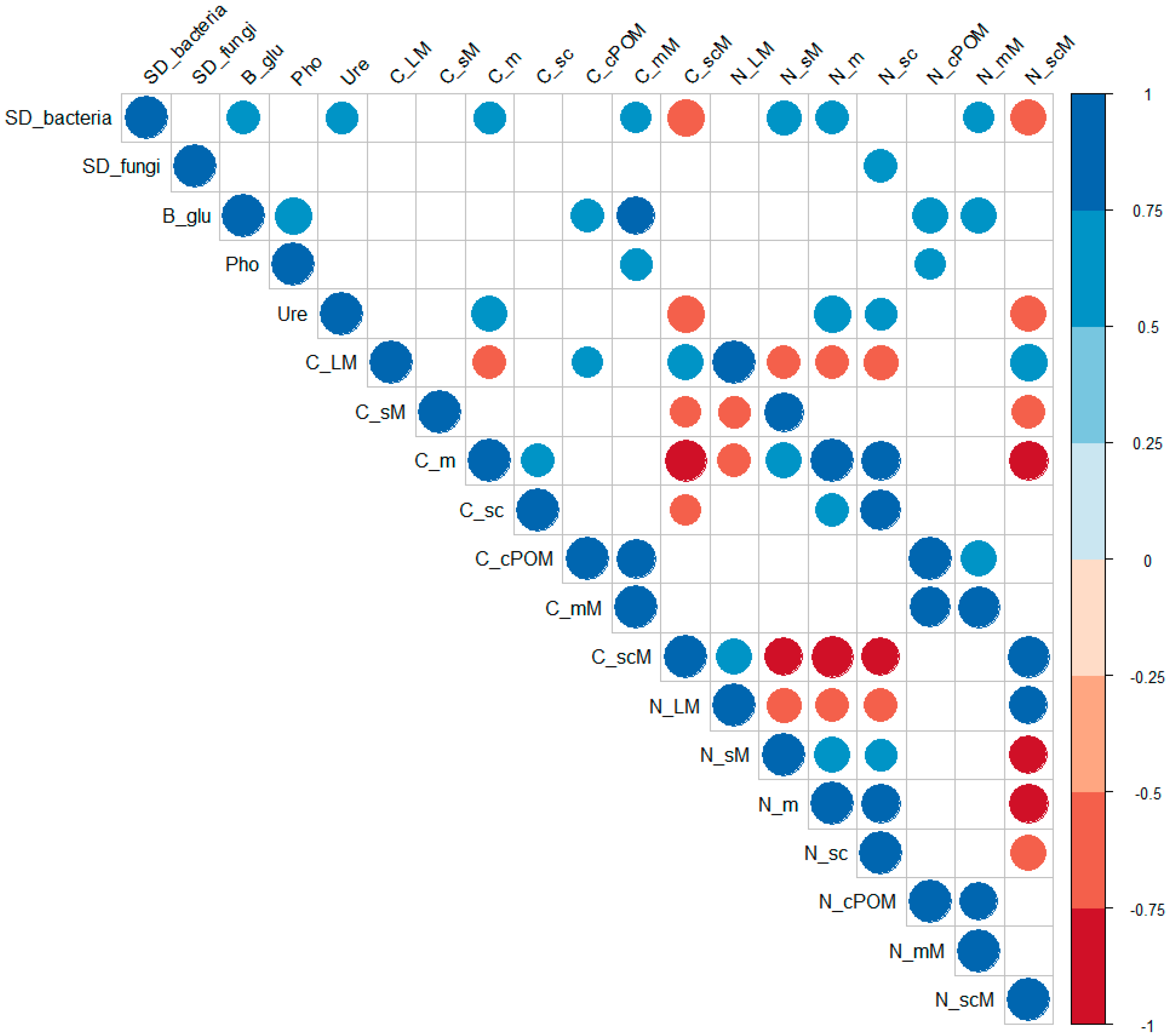
3.4. Correlations between the Microbiological and Physicochemical Properties
4. Discussion
4.1. Analyses of Soil Microbial Diversity
4.2. Enzymatic Activities and Soil Alteration Index
4.3. Aggregate-Associated C and N
4.4. Correlations between the Microbiological and Physicochemical Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, J.; Han, X.Z.; Ji, Z.J.; Li, Y.; Wang, E.T.; Xie, Z.H.; Chen, W.F. Abundance and diversity of soybean-nodulating rhizobia in black soil are impacted by land use and crop management. App. Environ. Microbiol. 2014, 80, 5394–5402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, M.A.; Nicholls, C.I.; Henao, A.; Lana, M.A. Agroecology and the design of climate change-resilient farming systems. Agron. Sustain. Dev. 2015, 35, 869–890. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. A system approach to conservation agriculture. J. Soil Water Conserv. 2015, 70, 82A–88A. [Google Scholar] [CrossRef] [Green Version]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37, 4. [Google Scholar] [CrossRef]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.T.; Mafongoya, P. The potential role of neglected and underutilised crop species as future crops under water scarce conditions in sub-Saharan Africa. Int. J. Environ. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [Green Version]
- Baraniya, D.; Puglisi, E.; Ceccherini, M.T.; Pietramellara, G.; Giagnoni, L.; Arenella, M.; Nannipieri, P.; Renella, G. Protease encoding microbial communities and protease activity of the rhizosphere and bulk soils of two maize lines with different N uptake efficiency. Soil Biol. Biochem. 2016, 96, 176–179. [Google Scholar] [CrossRef]
- Delgado Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome-from metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Cocconcelli, P.S.; Trevisan, M. Soil bacterial diversity screening using single 16S rRNA gene V regions coupled with multi-million read generating sequencing technologies. PLoS ONE 2012, 7, e42671. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 68, 12–26. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Coppolecchia, D.; Puglisi, E.; Balloi, A.; Mapelli, F.; Hamon, R.E.; Daffonchio, D.; Trevisan, M. Response of ammonia oxidizing bacteria and archaea to acute zinc stress and different moisture regimes in soil. Microb. Ecol. 2012, 64, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Li, W.; Dong, W.; Tian, Y.; Hu, C.; Liu, B. Tillage Changes vertical distribution of soil bacterial and fungal communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Kraut Cohen, J.; Zolti, A.; Shaltiel-Harpaz, L.; Argaman, E.; Rabinovich, R.; Green, S.J.; Minz, D. Effects of tillage practices on soil microbiome and agricultural parameters. Sci. Total Environ. 2020, 705, 135791. [Google Scholar] [CrossRef] [PubMed]
- Navarro Noya, Y.E.; Gómez Acata, S.; Montoya Ciriaco, N.; Rojas Valdez, A.; Suárez Arriaga, M.C.; Valenzuela Encinas, C.; Jiménez Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and validation of numerical indexes integrating enzyme activities of soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- Trost, B.; Prochnow, A.; Drastig, K.; Meyer-Aurich, A.; Ellmer, F.; Baumecker, M. Irrigation, soil organic carbon and N2O emissions. A review. Agron. Sustain. Dev. 2013, 33, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Six, J.; Elliott, E.T.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Bio. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Navarrete, A.A.; Venturini, A.M.; Meyer, K.M.; Klein, A.M.; Tiedje, J.M.; Bohannan, B.J.M.; Nüsslein, K.; Tsai, S.M.; Rodrigues, J.L.M. Differential response of acidobacteria subgroups to forest-to-pasture conversion and their biogeographic patterns in the western Brazilian Amazon. Front. Microbiol. 2015, 6, 1443. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, A.; Boselli, R.; Amaducci, S.; Tabaglio, V. Effects of no-till on root architecture and root-soil interactions in a three-year crop rotation. Eur. J. Agron. 2018, 99, 156–166. [Google Scholar] [CrossRef]
- Wang, X.; He, T.; Gen, S.; Zhang, X.-Q.; Wang, X.; Jiang, D.; Li, C.; Li, C.; Wang, J.; Zhang, W.; et al. Soil properties and agricultural practices shape microbial communities in flooded and rainfed croplands. Appl. Soil Ecol. 2020, 147, 103449. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Bertora, C.; Pepe, O.; Giancarlo, M.; Grignani, C.; Fagnano, M. Changes in soil mineral N content and abundances of bacterial communities involved in N reactions under laboratory conditions as predictors of soil N availability to maize under field conditions. Biol. Fertil. Soil 2016, 52, 523–537. [Google Scholar] [CrossRef]
- Fiorini, A.; Boselli, R.; Maris, S.C.; Santelli, S.; Ardenti, F.; Capra, F.; Tabaglio, V. May conservation tillage enhance soil C and N accumulation without decreasing yield in intensive irrigated croplands? Results from an eight-year maize monoculture. Agric. Ecosyst. Environ. 2020, 296, 106926. [Google Scholar] [CrossRef]
- Fiorini, A.; Maris, S.C.; Abalos, D.; Amaducci, S.; Tabaglio, V. Combining no-till with rye (Secale cereale L.) cover crop mitigates nitrous oxide emissions without decreasing yield. Soil Tillage Res. 2020, 196, 104442. [Google Scholar] [CrossRef]
- Kyei Boahen, S.; Savala, C.E.N.; Chikoye, D.; Abaidoo, R. Growth and yield responses of cowpea to inoculation and phosphorus fertilization in different environments. Front. Plant. Sci 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mastro, F.; Traversa, A.; Cocozza, C.; Pallara, M.; Brunetti, G. Soil organic carbon stabilization: Influence of tillage on mineralogical and chemical parameters. Soil Syst. 2020, 4, 58. [Google Scholar] [CrossRef]
- Gattullo, C.E.; Mezzapesa, G.N.; Stellacci, A.M.; Ferrara, G.; Occhiogrosso, G.; Petrelli, G.; Castellini, M.; Spagnuolo, M. Cover crop for a sustainable viticulture: Effects on soil properties and table grape production. Agronomy 2020, 10, 1334. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, E.; Sun, O.J. Can no-tillage stimulate carbon sequestration in agricultural soils? A meta-analysis of paired experiments. Agric. Ecosyst. Environ. 2010, 139, 224–231. [Google Scholar] [CrossRef]
- Powlson, D.S.; Stirling, C.M.; Jat, M.L.; Gerard, B.G.; Palm, C.A.; Sanchez, P.A.; Cassman, K.G. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Chang. 2014, 4, 678–683. [Google Scholar] [CrossRef]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef]
- Boselli, R.; Fiorini, A.; Santelli, S.; Ardenti, F.; Capra, F.; Maris, S.C.; Tabaglio, V. Cover crops during transition to no-till maintain yield and enhance soil fertility in intensive agro-ecosystems. Field Crop. Res. 2020, 255, 107871. [Google Scholar] [CrossRef]
- Guzzetti, L.; Fiorini, A.; Panzeri, D.; Tommasi, N.; Grassi, F.; Taskin, E.; Misci, C.; Puglisi, E.; Tabaglio, V.; Galimberti, A.; et al. Sustainability perspectives of Vigna unguiculata L. Walp. Cultivation under no tillage and water stress conditions. Plants 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.A.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91, fiv114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandini, F.; Misci, C.; Taskin, E.; Cocconcelli, P.S.; Puglisi, E. Biopolymers modulate microbial communities in municipal organic waste digestion. FEMS Microbiol. Ecol. 2020, 96, fiaa183. [Google Scholar] [CrossRef] [PubMed]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Veen, J.A.; Cocconcelli, P.S.; Trevisan, M. Soil microbial diversity patterns of a lowland spring environment. FEMS Microbiol. Ecol. 2013, 86, 172–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. App. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. NAST: A multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D. The effects of alignment quality, distance calculation method, sequence filtering, and region on the analysis of 16S rRNA gene-based studies. PLoS Comput. Biol. 2010, 6, e1000844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2011, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Eivazi, F.; Tabatabai, M.A. Factors affecting glucosidase and galactosidase activities in soils. Soil Biol. Biochem. 1990, 22, 891–897. [Google Scholar] [CrossRef]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soil 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Elliott, E.T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils1. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil structure and organic matter I. Distribution of aggregate-size classes and aggregate-associated carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenhau, R.C. Aggregate Stability and Size Distribution. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Properties, Including Statistics of Measurement and Sampling; Black, C.A., Ed.; ASA SSSA: Madison, WI, USA, 1965; Volume 9, pp. 511–519. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Paulson, J.N.; Pop, M.; Bravo, H.C. Metastats: An improved statistical method for analysis of metagenomic data. Genome Biol. 2011, 12, P17. [Google Scholar] [CrossRef] [Green Version]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Carof, M.; Chen, W.; Le Floch, G. Effect of tillage and static abiotic soil properties on microbial diversity. App. Soil Ecol. 2018, 132, 135–145. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef] [Green Version]
- Babin, D.; Deubel, A.; Jacquiod, S.; Sørensen, S.J.; Geistlinger, J.; Grosch, R.; Smalla, K. Impact of long-term agricultural management practices on soil prokaryotic communities. Soil Biol. Biochem. 2019, 129, 17–28. [Google Scholar] [CrossRef]
- Essel, E.; Xie, J.H.; Deng, C.C.; Peng, Z.K.; Wang, J.B.; Shen, J.C.; Xie, J.H.; Coulter, J.A.; Li, L.L. Bacterial and fungal diversity in rhizosphere and bulk soil under different long-term tillage and cereal/legume rotation. Soil Tillage Res. 2019, 194, 104302. [Google Scholar] [CrossRef]
- Chavez Romero, Y.; Navarro Noya, Y.E.; Reynoso Martinez, S.C.; Sarria Guzman, Y.; Govaerts, B.; Verhulst, N.; Dendooven, L.; Luna Guido, M. 16S metagenomics reveals changes in the soil bacterial community driven by soil organic C, N-fertilizer and tillage-crop residue management. Soil Tillage Res. 2016, 159, 1–8. [Google Scholar] [CrossRef]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 2015, 86, 158–164. [Google Scholar] [CrossRef]
- Sánchez Marañón, M.; Miralles, I.; Aguirre-Garrido, J.F.; Anguita-Maeso, M.; Millán, V.; Ortega, R.; García Salcedo, J.A.; Martínez-Abarca, F.; Soriano, M. Changes in the soil bacterial community along a pedogenic gradient. Sci. Rep. 2017, 7, 14593. [Google Scholar] [CrossRef]
- Mangalassery, S.; Mooney, S.J.; Sparkes, D.L.; Fraser, W.T.; Sjögersten, S. Impacts of zero tillage on soil enzyme activities, microbial characteristics and organic matter functional chemistry in temperate soils. Eur. J. Soil Biol. 2015, 68, 9–17. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Larney, F.J.; Blackshaw, R.E.; Kanashiro, D.A.; Pearson, D.C. Phospholipid fatty acid biomarkers show positive soil microbial community responses to conservation soil management of irrigated crop rotations. Soil Tillage Res. 2017, 168, 1–10. [Google Scholar] [CrossRef]
- Schutter, M.; Sandeno, J.; Dick, R. Seasonal, soil type, and alternative management influences on microbial communities of vegetable cropping systems. Biol. Fertil. Soils 2001, 34, 397–410. [Google Scholar] [CrossRef]
- Panettieri, M.; Knicker, H.; Berns, A.E.; Murillo, J.M.; Madejón, E. Moldboard plowing effects on soil aggregation and soil organic matter quality assessed by 13C CPMAS NMR and biochemical analyses. Agric. Ecosyst. Environ. 2013, 177, 48–57. [Google Scholar] [CrossRef]
- Siebielec, S.; Siebielec, G.; Klimkowicz-Pawlas, A.; Gałazka, A.; Grzadziel, J.; Stuczynski, T. Impact of water stress on microbial community and activity in sandy and loamy soils. Agronomy 2020, 10, 1429. [Google Scholar] [CrossRef]
- Marxsen, J.; Zoppini, A.; Wilczek, S. Microbial communities in streambed sediments recovering from desiccation. FEMS Microbiol. Ecol. 2010, 71, 374–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoppini, A.; Marxsen, J. Importance of extracellular enzymes for biogeochemical processes in temporary river sediments during fluctuating dry-wet conditions. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 103–117. [Google Scholar]
- Jain, N.K.; Jat, R.S.; Meena, H.N.; Chakraborty, K. Productivity, nutrient, and soil enzymes influenced with conservation agriculture practices in peanut. Agron. J. 2018, 110, 1165–1172. [Google Scholar] [CrossRef]
- Zibilske, L.M.; Bradford, J.M. Soil aggregation, aggregate carbon and nitrogen, and moisture retention induced by conservation tillage. Soil Sci. Soc. Am. J. 2007, 71, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Lichter, K.; Govaerts, B.; Six, J.; Sayre, K.D.; Deckers, J.; Dendooven, L. Aggregation and C and N contents of soil organic matter fractions in a permanent raised-bed planting system in the Highlands of Central Mexico. Plant Soil 2008, 305, 237–252. [Google Scholar] [CrossRef]
- Huang, S.; Sun, Y.-N.; Rui, W.-Y.; Liu, W.-R.; Zhang, W.-J. Long-term effect of no-tillage on soil organic carbon fractions in a continuous maize cropping system of northeast China. Pedosphere 2010, 20, 285–292. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Bergmann, J.; Verbruggen, E.; Veresoglou, S.D.; Lehmann, A. Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phytol. 2015, 205, 1385–1388. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K.; Doran, J.W. Aggregation and soil organic matter accumulation in cultivated and native grassland soils. Soil Sci. Soc. Am. J. 1998, 62, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Denef, K.; Six, J.; Merckx, R.; Paustian, K. Carbon sequestration in microaggregates of no-tillage soils with different clay mineralogy. Soil Sci. Soc. Am. J. 2004, 68, 1935–1944. [Google Scholar] [CrossRef]
- Balesdent, J.; Chenu, C.; Balabane, M. Relationship of soil organic matter dynamics to physical protection and tillage. Soil Tillage Res. 2000, 53, 215–230. [Google Scholar] [CrossRef]
- Smith, S.F.; Brye, K.R.; Gbur, E.E.; Chen, P.Y.; Korth, K. Residue and water management effects on aggregate stability and aggregate-associated carbon and nitrogen in a wheat-soybean, double-crop system. Soil Sci. Soc. Am. J. 2014, 78, 1378–1391. [Google Scholar] [CrossRef]
- Sainju, U.M.; Caesar-TonThat, T.; Jabro, J.D. Carbon and nitrogen fractions in dryland soil aggregates affected by long-term tillage and cropping sequence. Soil Sci. Soc. Am. J. 2009, 73, 1488–1495. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.J.; Gregorich, E.G.; McLaughlin, N.B.; Zhang, X.P.; Guo, Y.F.; Liang, A.Z.; Fan, R.Q.; Sun, B.J. No-tillage with continuous maize cropping enhances soil aggregation and organic carbon storage in Northeast China. Geoderma 2018, 330, 204–211. [Google Scholar] [CrossRef]
- Wang, X.; Qi, J.Y.; Zhang, X.Z.; Li, S.S.; Virk, A.L.; Zhao, X.; Xiao, X.P.; Zhang, H.L. Effects of tillage and residue management on soil aggregates and associated carbon storage in a double paddy cropping system. Soil Tillage Res. 2019, 194, 104339. [Google Scholar] [CrossRef]
- Lopez Bellido, R.J.; Munoz Romero, V.; Fuentes Guerra, R.; Fernandez Garcia, P.; Lopez Bellido, L. No-till: A key tool for sequestering C and N in microaggregates on a Mediterranean Vertisol. Soil Tillage Res. 2017, 166, 131–137. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Oades, J.M. The role of biology in the formation, stabilization and degradation of soil structure. In Soil Structure/Soil Biota Interrelationships; Brussaard, L., Kooistra, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 377–400. [Google Scholar] [CrossRef]
- Sekaran, U.; Sagar, K.L.; Denardin, L.G.D.O.; Singh, J.; Singh, N.; Abagandura, G.O.; Kumar, S.; Farmaha, B.S.; Bly, A.; Martins, A.P. Responses of soil biochemical properties and microbial community structure to short and long-term no-till systems. Eur. J. Soil Sci. 2019, 71, 1018–1033. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measured Soil Enzymes | |||||
|---|---|---|---|---|---|
| Source of Variation | AI3 5 | β GLU (μmol PNG g−1 h−1) | PHO (μmol PNP g−1 h−1) | URE (μg urea g−1 h−1) | |
| Tillage (T) | CT 1 | −85.35 (±5.05) b | 8.86 (±0.8) b | 18.51 (±0.48) b | 5.85 (±1.07) a |
| NT + CC 2 | −38.43 (±5.86) a | 20.85 (±2) a | 24.21 (±1.6) a | 7.05 (±1.1) a | |
| Water (W) | NS 3 | −60.72 (±4.47) a | 16.84 (±4.17) a | 23.13 (±1.85) a | 6.47 (±1.63) a |
| S 4 | −63.05 (±5.83) a | 12.86 (±1.41) a | 19.6 (±0.55) a | 6.42 (±1.12) a | |
| T × W | CT-NS | −96.14 (±3.63) b | 8.06 (±0.81) b | 19.04 (±0.53) ab | 6.18 (±1.13) a |
| CT-S | −74.56 (±4.65) ab | 9.65 (±0.85) b | 17.98 (±0.39) b | 5.51 (±1.1) a | |
| NT + CC-NS | −25.3 (±3.33) a | 25.63 (±1.51) a | 27.21 (±1.4) a | 6.77 (±2.52) a | |
| NT + CC-S | −72.09 (±5.38) ab | 16.07 (±1.14) b | 23.72 (±1.08) ab | 7.33 (±1.99) a | |
| Depth | Source of Variation | Code | Whole Soil—C Amount (g C kg−1 Soil) | Macroaggregates—C Amount (g C kg−1 Soil) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LM 1 | sM 2 | m 3 | s + c 4 | cPOM 5 | mM 6 | s + cM 7 | |||||||
| 0–5 cm | Tillage (T) | CT | 2.71 | b | 6.49 | 1.73 | 1.43 | 0.66 | b | 6.34 | b | 2.75 | |
| NT + CC | 7.37 | a | 5.67 | 1.85 | 1.66 | 2.20 | a | 8.29 | a | 3.65 | |||
| p value | 0.0093 | 0.4249 | 0.5076 | 0.2816 | 0.0039 | 0.0433 | 0.0688 | ||||||
| Water (W) | NS | 6.20 | 6.59 | 1.77 | 1.63 | 1.66 | a | 7.59 | a | 3.55 | |||
| S | 3.88 | 5.57 | 1.81 | 1.45 | 1.19 | b | 7.03 | b | 2.85 | ||||
| p value | 0.0779 | 0.2344 | 0.7912 | 0.3374 | 0.0401 | 0.0258 | 0.0676 | ||||||
| T × W | CT-NS | 3.74 | 5.52 | 1.63 | 1.57 | 0.43 | c | 6.27 | b | 2.93 | |||
| CT-S | 1.68 | 7.47 | 1.83 | 1.28 | 0.89 | bc | 6.40 | ab | 2.57 | ||||
| NT + CC-NS | 8.68 | 5.62 | 1.91 | 1.69 | 2.90 | a | 8.92 | a | 4.18 | ||||
| NT + CC-S | 6.07 | 5.72 | 1.79 | 1.62 | 1.50 | b | 7.65 | ab | 3.13 | ||||
| p value | 0.8142 | 0.2800 | 0.3219 | 0.5429 | 0.0028 | 0.0109 | 0.3187 | ||||||
| 5–20 cm | Tillage (T) | CT | 4.59 | 5.24 | 1.42 | 0.89 | 0.57 | 5.97 | 3.84 | a | |||
| NT + CC | 4.17 | 5.64 | 1.62 | 0.94 | 0.56 | 6.61 | 2.70 | b | |||||
| p value | 0.6124 | 0.4097 | 0.0636 | 0.5325 | 0.7415 | 0.0720 | 0.0396 | ||||||
| Water (W) | NS | 4.89 | 5.53 | 1.31 | 0.81 | 0.60 | 6.31 | 3.53 | |||||
| S | 3.87 | 5.35 | 1.73 | 1.01 | 0.53 | 6.27 | 3.01 | ||||||
| p value | 0.1196 | 0.5502 | 0.0698 | 0.1453 | 0.5609 | 0.8356 | 0.0688 | ||||||
| T × W | CT-NS | 4.94 | 4.92 | 1.24 | 0.83 | 0.55 | 5.91 | 4.15 | |||||
| CT-S | 4.24 | 5.57 | 1.59 | 0.94 | 0.60 | 6.02 | 3.52 | ||||||
| NT + CC-NS | 4.84 | 5.50 | 1.37 | 0.79 | 0.65 | 6.71 | 2.92 | ||||||
| NT + CC-S | 3.50 | 4.92 | 1.86 | 1.08 | 0.46 | 6.51 | 2.49 | ||||||
| p value | 0.5888 | 0.1531 | 0.6745 | 0.4593 | 0.3593 | 0.4858 | 0.6980 | ||||||
| Depth | Source of Variation | Code | Whole Soil—N Amount (g N kg−1 Soil) | Macroaggregates—N Amount (g N kg−1 Soil) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LM 1 | sM 2 | m 3 | s + c 4 | cPOM 5 | mM 6 | s + cM 7 | ||||||||
| 0–5 cm | Tillage (T) | CT | 0.38 | b | 0.76 | 0.22 | 0.14 | 0.05 | b | 0.95 | 0.34 | b | ||
| NT + CC | 0.87 | a | 0.65 | 0.21 | 0.16 | 0.17 | a | 0.79 | 0.51 | a | ||||
| p value | 0.0317 | 0.2986 | 0.9460 | 0.1566 | 0.0040 | 0.1375 | 0.0356 | |||||||
| Water (W) | NS | 0.73 | 0.78 | 0.20 | 0.16 | 0.12 | a | 0.88 | 0.45 | |||||
| S | 0.51 | 0.63 | 0.23 | 0.15 | 0.09 | b | 0.87 | 0.40 | ||||||
| p value | 0.1188 | 0.1685 | 0.3011 | 0.5377 | 0.0426 | 0.6622 | 0.3658 | |||||||
| T × W | CT-NS | 0.45 | 0.62 | 0.19 | 0.14 | 0.03 | c | 0.78 | 0.36 | |||||
| CT-S | 0.30 | 0.90 | 0.24 | 0.13 | 0.07 | bc | 0.80 | 0.32 | ||||||
| NT + CC-NS | 1.02 | 0.63 | 0.21 | 0.17 | 0.21 | a | 0.98 | 0.54 | ||||||
| NT + CC-S | 0.72 | 0.67 | 0.21 | 0.16 | 0.12 | b | 0.93 | 0.47 | ||||||
| p value | 0.5748 | 0.2753 | 0.3251 | 0.8816 | 0.0020 | 0.2588 | 0.7685 | |||||||
| 5–20 cm | Tillage (T) | CT | 0.58 | 0.62 | 0.17 | b | 0.12 | 0.04 | 0.75 | b | 0.49 | |||
| NT + CC | 0.53 | 0.71 | 0.21 | a | 0.13 | 0.05 | 0.88 | a | 0.38 | |||||
| p value | 0.6322 | 0.2829 | 0.0479 | 0.5391 | 0.1422 | 0.0213 | 0.0547 | |||||||
| Water (W) | NS | 0.62 | 0.66 | 0.22 | a | 0.11 | 0.05 | 0.81 | 0.46 | |||||
| S | 0.49 | 0.69 | 0.16 | b | 0.14 | 0.04 | 0.82 | 0.41 | ||||||
| p value | 0.1118 | 0.8928 | 0.0351 | 0.0998 | 0.4358 | 0.7583 | 0.2126 | |||||||
| T × W | CT-NS | 0.64 | 0.58 | 0.15 | 0.11 | 0.04 | 0.75 | 0.52 | ||||||
| CT-S | 0.52 | 0.68 | 0.19 | 0.12 | 0.04 | 0.75 | 0.45 | |||||||
| NT + CC-NS | 0.61 | 0.74 | 0.17 | 0.10 | 0.06 | 0.88 | 0.39 | |||||||
| NT + CC-S | 0.46 | 0.67 | 0.24 | 0.16 | 0.05 | 0.89 | 0.36 | |||||||
| p value | 0.8395 | 0.1653 | 0.5989 | 0.1403 | 0.3549 | 0.7353 | 0.6329 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taskin, E.; Boselli, R.; Fiorini, A.; Misci, C.; Ardenti, F.; Bandini, F.; Guzzetti, L.; Panzeri, D.; Tommasi, N.; Galimberti, A.; et al. Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators. Biology 2021, 10, 23. https://doi.org/10.3390/biology10010023
Taskin E, Boselli R, Fiorini A, Misci C, Ardenti F, Bandini F, Guzzetti L, Panzeri D, Tommasi N, Galimberti A, et al. Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators. Biology. 2021; 10(1):23. https://doi.org/10.3390/biology10010023
Chicago/Turabian StyleTaskin, Eren, Roberta Boselli, Andrea Fiorini, Chiara Misci, Federico Ardenti, Francesca Bandini, Lorenzo Guzzetti, Davide Panzeri, Nicola Tommasi, Andrea Galimberti, and et al. 2021. "Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators" Biology 10, no. 1: 23. https://doi.org/10.3390/biology10010023
APA StyleTaskin, E., Boselli, R., Fiorini, A., Misci, C., Ardenti, F., Bandini, F., Guzzetti, L., Panzeri, D., Tommasi, N., Galimberti, A., Labra, M., Tabaglio, V., & Puglisi, E. (2021). Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators. Biology, 10(1), 23. https://doi.org/10.3390/biology10010023